
Abstract
Hormone-sensitive lipase (HSL) is a key regulator of cholesterol esters metabolism. The aim of this study was to determine HSL localization in rat female reproductive organs during the ovarian cycle by IHC methods. HSL was located in the ovarian epithelium. The granulosa cells and oocytes of primordial follicles were immunonegative. In mature follicles, HSL was found in oocytes and theca and granulosa cells. However, HSL expression in theca cells and oocytes decreased during follicular atresia. Luteal cells showed HSL staining in cytoplasm during proestrus and estrus, in the nucleus during metestrus, and in cytoplasm and the nucleus during diestrus. In the tubaric ampulla, HSL was located in the epithelial cells nuclei and in the cilia during proestrus and estrus but mainly in the nucleus during metestrus and diestrus. In the isthmus, cells showed HSL immunolabeling in the nucleus and cilia during proestrus, but only in the cilia during estrus, metestrus, and diestrus. In the uterus, HSL was found in the epithelial cells nuclei. HSL-immunoreactive bands at 84, 67, 54, and 43 kDa were found in rat female reproductive organs. HSL labeling in the nucleus of epithelial and germ cells suggests an as yet unknown function for this protein, probably related to oogenesis and cell proliferation.
H
This multifunctional enzyme has merged as a key regulator of lipid metabolism in multiple tissues. HSL activity or HSL mRNA has been found in white and brown adipose tissue, adrenal glands, ovaries, corpora lutea, testis, placenta, macrophages, heart, skeletal and smooth muscle, and mammary glands (Cook et al. 1983; Holm et al. 1987; Small et al. 1991; Fredrikson et al. 1992; Kraemer et al. 1993; Holst et al. 1994; Martín-Hidalgo et al. 2005).
The HSL gene is located on chromosome 19q13.3 (Holm et al. 1988) and was initially reported to contain nine exons that encode the adipocyte protein, spanning 11 and 10 kb in humans (Langin et al. 1993) and mice (Li et al. 1994), respectively. Interestingly, the size of HSL mRNAs is variable. In rat heart, skeletal muscle, placenta, and ovaries, the expressed HSL mRNAs are slightly larger (3.5 kb) than in adipose tissue (3.3 kb) (Holm et al. 1988). The mRNA species expressed in the testis are even larger at 3.9 kb (Holm et al. 1988; Holst et al. 1994; Mairal et al. 2002).
The purified adipose tissue enzyme has a molecular mass of 84 and 88 kDa on SDS-PAGE, in rats (Holm et al. 1988) and humans (Langin et al. 1993), respectively. Recently, a novel 89-kDa HSL isoform has been described that is expressed in rat pancreatic β cells, adipocytes, adrenal glands, and ovaries (Lindvall et al. 2004). The testis expresses several protein species with apparent molecular masses ranging from 26 to 130 kDa, encoded by a single 3.9-kb HSL mRNA in rat testis (Kraemer et al. 1993) and both 3.3- and 3.9-kb HSL mRNAs in human testis (Holst et al. 1994; Mairal et al. 2002; Lindvall et al. 2004). Moreover, in the rodent testis, HSL is located only in the seminiferous tubules (Sertoli and spermatogenic cells), whereas in the normal human testis, HSL is located in both seminiferous tubules and Leydig cells (Arenas et al. 2004).
Although it is clear that HSL acts as a triacylglycerol lipase in adipose tissue and, perhaps, in muscles that have accumulated triglycerides, it is the ability of HSL to hydrolyze stored cholesterol esters to free cholesterol for use in steroid hormone production that predominates in adrenal, ovary, and testis tissues (Trzeciak and Boyd 1973; Beins et al. 1982; Cook et al. 1982).
The cholesterol used for steroidogenesis is derived from a range of sources (Gwynne and Strauss 1982): (a) the mobilization of stored cholesterol esters through the action of neutral cholesteryl ester hydrolase activity (CEH), (b) lipoprotein-derived cholesteryl esters obtained by cellular uptake, and (c) de novo cholesterol synthesis. Although the exact functional role of HSL in tissues other than adipose and steroid-producing tissues is not completely understood, it seems that, in these tissues, HSL participates in intracellular cholesterol homeostasis by mobilizing stored cholesterol for synthesis of steroid hormones or for excretion. The most direct evidence comes from HSL knockout mice, where no detectable HSL and no neutral cholesteryl ester hydrolase activity are observed in the testis (Osuga et al. 2000) or in the adrenal glands (Kraemer et al. 2002). This being the case, a direct interaction of HSL with steroidogenic acute regulatory protein (StAR) has been proposed (Shen et al. 2003). The interaction of HSL with StAR in cytosol increases the hydrolytic activity of HSL and facilitates cholesterol movement from stored lipid droplets to mitochondria for steroidogenesis. Recently, it has also been shown that HSL is required to hydrolyze the cholesterol esters from high-density lipoproteins and provide free cholesterol to support adrenal steroidogenesis (Kraemer et al. 2004).
The roles of HSL in the male reproductive system are multiple and complex. The presence of HSL in human Leydig cells suggests a role for this protein in steroidogenesis and testicular cholesterol metabolism. Furthermore, this protein seems to be involved in spermiogenesis, as suggested by its presence in spermatids (Kraemer et al. 1997; Osuga et al. 2000; Arenas et al. 2004) and because HSL knockout mice are infertile (Osuga et al. 2000).
The presence of HSL in ovaries has been previously reported (Cook et al. 1983; Holm et al. 1987; Kraemer et al. 1993; Lindvall et al. 2004), and it has also been shown that overexpression of HSL in Chinese hamster ovary cells leads to abnormalities in cholesterol homeostasis (Kraemer et al. 1997). However, there are no previous studies of its cellular localization and how this might change during the estrus cycle of the ovary. To gain a better understanding of the potential function of HSL in the female reproductive system, this study was carried out to determine the cell types that might express this protein in the rat ovary, oviduct, and uterus and to assess the changes that take place in its cellular localization and expression during the different phases of the estrus cycle.
Materials and Methods
Animals and Tissue Collection
Prepuberal and adult female Wistar rats from our colony, weighing 200-225 g, were used for this study. Rats were maintained at 22-24C under standard conditions of illumination (from 8:00 AM to 8:00 PM) and feeding (Purina Chow diet; Panlab, Barcelona, Spain). The animals were fed ad libitum and had constant access to tap water. The experimental protocol was approved by the Animal Research Committee of the Hospital Ramón y Cajal, Madrid, Spain.
Vaginal smears were taken to determine the different phases of the estrus cycle: proestrus, estrus, metestrus, and diestrus. At least six rats at each phase of the estrus cycle were analyzed. Animals in all phases of the estrus cycle were killed between 10:00 and 11:00 AM by decapitation after normal overnight access to food. Tissue samples were immediately removed from the rats, frozen in liquid nitrogen, and stored at −80C until processing for protein extraction (see Western Blotting) or fixed in 4% paraformaldehyde in PBS for 6-24 hr at 4C. The fixed tissues were embedded in paraffin following conventional methods and sectioned at 5 μm, either to be stained with hematoxylin and eosin or to be used for IHC.
Western Blotting
Tissues were homogenized in a buffer containing 0.25 M sucrose, 1 mM EDTA, 1 mM dithiothreitol, 20 μg/ml antipain, and 20 μg/ml leupeptin at pH 7.4. Protein determination was performed by the Bradford dye method (Bradford 1976), using Bio-Rad reagent and using BSA as the standard. Cellular proteins (20 μg) were dissolved in SDS-PAGE sample solution [60 mM Tris, pH 6.5, 10% (w/v) glycerol, 5% (w/v) β-mercaptoethanol, 20% (w/v) SDS, and 0.025% (w/v) bromophenol blue] by boiling (2 × 5 min), and vortex mixing (2×30 sec). After fractionation by SDS-PAGE in slab gels (7 × 14 cm), proteins were electroblotted onto polyvinylidene difluoride (PVDF) membranes (Millipore; Bedford, MA) using a blotting apparatus (BioRad Laboratories; Hercules, CA). Reference proteins were myosin (Mr 209,000), β-galactosidase (Mr 137,000), BSA (Mr 84,000), carbonic anhydrase (Mr 44,000), soybean trypsin inhibitor (Mr 32,800), lysozyme (Mr 18,700), and aprotinin (Mr 7200) (all from BioRad). Blots were blocked for 2 hr in 10% defatted dried milk in Tris-buffered saline (TBS)-0.1% Tween and incubated for 2 hr at room temperature with the primary antibody [1:20,000; chicken anti-HSL (LIPE), polyclonal antibody correspond to synthetic peptides direct against HSL from white rat adipose tissue (Affinity Bioreagents)] in 5% defatted dried milk-TBS-0.1% Tween. This was followed by 1-hr incubation with the secondary peroxidase-conjugated antibody (1:20,000; donkey anti-chicken IgY, RDI Research Diagnostics, Flanders, NJ) in 5% defatted dried milk-TBS-0.1% Tween. All steps were performed at room temperature, and blots were washed between incubation steps with TBS-0.1% Tween. The immunoreactive bands were visualized using the enhanced chemiluminescence detection method according to the manufacturer's instructions (ECL system; Amersham, Dübendorf, Switzerland) and subsequent exposure of the membrane to X-ray film.
The specificity of the procedure was assessed by means of negative controls that were performed omitting the primary antibody and incubating only with the secondary antibody at optimal titters.
IHC
Streptavidin-peroxidase immunostaining was performed as previously described (Arenas et al. 2004; Martín-Hidalgo et al. 2005). Briefly, serial sections were mounted on silanized slides and allowed to dry overnight before IHC staining. Paraffin was removed with xylene. The sections were hydrated with H2O in a graded ethanol series and double-distilled water and placed in a glass jar containing 10 mM sodium citrate buffer, pH 6.0, and heated in a microwave oven for 4 × 2 min cycles at medium power. The sections were allowed to cool inside the microwave oven for 20 min. After rinsing with double-distilled water, the sections were washed twice in TBS buffer, pH 7.6, for 5 min. The endogenous peroxidase activity was inhibited by incubation with 2% H2O2 for 10 min. Sections were washed with H2O and TBS and incubated with 3% normal goat serum, 0.01% Triton X-100, and 0.1% glycine in TBS, pH 7.6, at room temperature for 30 min. to prevent nonspecific binding of the first antibody. The sections were incubated for 12 hr at 4C with the primary antibody, anti-HSL (1:800; chicken polyclonal antiserum directed against HSL from white rat adipose tissue) diluted in 0.3% normal goat serum, 0.001% Triton X-100, and 0.01% glycine in TBS at pH 7.6. The sections were washed twice in TBS to remove unbound primary antibody and incubated with the secondary antibody for 1 hr at room temperature. The biotinylated second antibody used was goat anti-chicken IgY (1:200) for HSL (Vector Labs; Burlingame, CA), diluted in 0.3% normal goat serum, 0.001% Triton X-100, and 0.01% glycine in TBS, pH 7.6. Sections were rinsed in TBS and incubated with the streptavidin-peroxidase complex (Zymed Labs; San Francisco, CA) for 30 min and washed in TBS followed by Tris-HCl buffer, pH 7.6. Peroxidase activity was developed using DAB as the chromogen (Sigma; St. Louis, MO). The sections were counterstained with Carazzi's hematoxylin. Thereafter, the sections were dehydrated in ethanol, cleared with xylene, mounted in DePeX, and observed under a light microscope.
The specificity of the IHC procedures was assessed by means of negative controls that were performed as follows: (a) omitting the primary antibodies, (b) using non-immune serum instead of the primary antibodies, and (c) incubating with an inappropriate secondary antibody after the incubation with the primary antibodies at optimal titers.
The staining intensity was qualitatively evaluated and classified as absent (-), weak (+), moderate (++), or intense (+++). The assessment of the grade of staining was performed blinded, always by two independent investigators, in high-power fields (X400) using standard light microscopy. When there was a disagreement between observers on scores, the observers were blinded to their last score, and consensus judgment was reached through discussion.
Results
HSL Location and Protein Expression in the Rat Ovary: Follicular Maturation
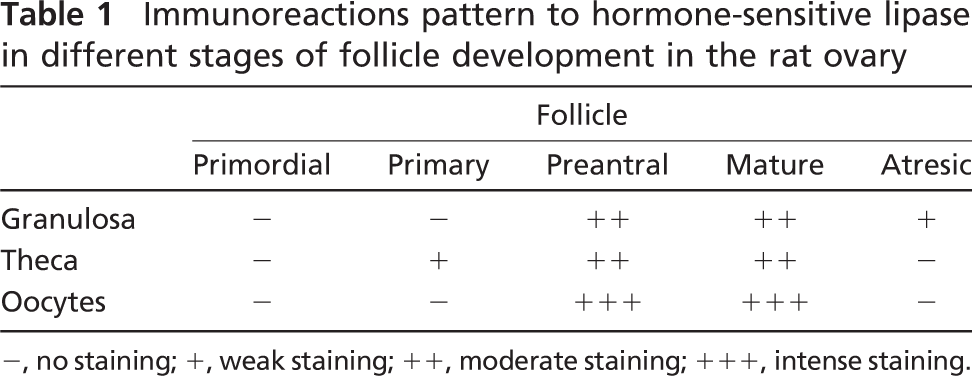
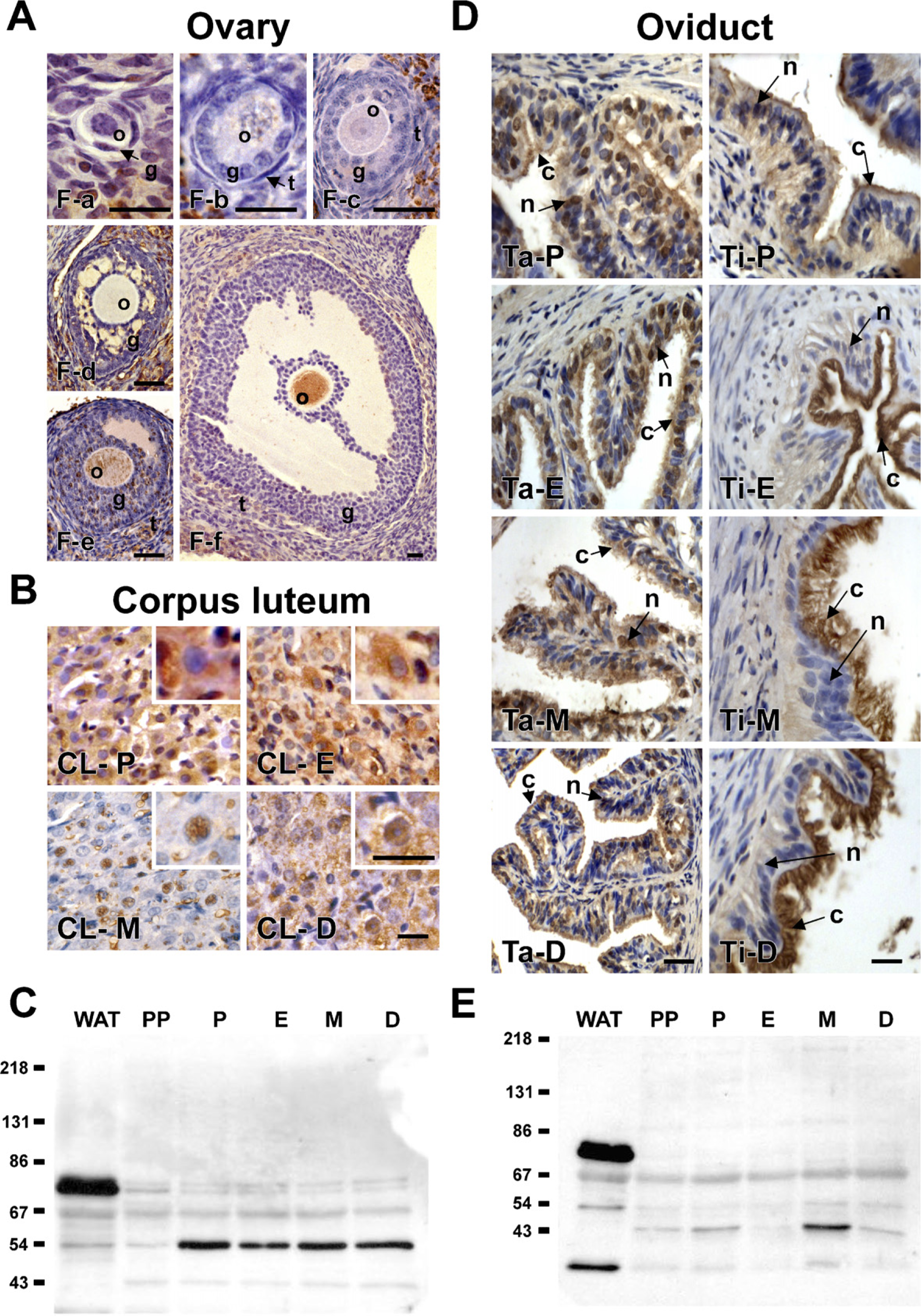
The patterns of HSL immunostaining in the different stages of follicle development in the rat ovary are summarized in Table 1. HSL immunostaining varied with the different stages of follicle (F) development (Figure 1A). In primordial follicles, the single layer of granulosa (g) cells and the immature oocytes (o) were immunonegative (Figure 1A, F-a and F-b). When follicle development began and primary follicles contained two or three layers of granulosa cells, the oocytes and granulosa cells remained unstained, and the initial theca (t) cells became immunostained (Figure 1A, F-c). In preantral follicles showing a developed theca bilayer, HSL was found in the internal and the external theca cells, granulosa cells, and oocytes (Figure 1A, F-e). When the oocytes were positively stained, HSL appeared to be distributed in both the cytoplasm and nucleus (Figure 1A, F-e). However, HSL was located in the nucleus of the granulosa cells and in the nucleus and cytoplasm of the theca cells in mature follicles (Figure 1A, F-e and F-f). In contrast to mature follicles (Figure 1A, F-e and F-f), HSL-positive immunoreactivity progressively disappeared from the oocyte and theca cells during follicular atresia (Figure 1A, F-d). Atresic follicles were identified by the presence of pyknotic cells in the granulosa layer, cell debris in the follicular antrum, a shrinking oocyte, accumulation of lipid droplets in theca and granulosa cells, and the final appearance of the collapsed follicle with a hypertrophied theca layer (Figures 1A, F-d).
Immunoreactions pattern to hormone-sensitive lipase in different stages of follicle development in the rat ovary
−, no staining; +, weak staining; ++, moderate staining; +++, intense staining.
No immunostaining was observed in the negative controls (data not shown).
HSL immunoreactivity changes were observed in the corpus luteum (CL) in the different phases of the ovarian cycle (Figure 1B, CL). Positive HSL staining was observed in the cytoplasm of the luteal cells during the proestrus and estrus phases of the ovarian cycle (Figure 1B, CL-P and CL-E). However, metestrus corpora lutea showed positive HSL staining only in the nucleus of luteal cells (Figure 1B, CL-M); during diestrus, positive labeling for HSL in the corpora lutea was found in the cytoplasm and nucleus of the luteal cells (Figure 1B, CL-D).
Western blot analyses were used to explore changes in HSL protein expression in the rat ovary during the different phases of the estrus cycle, and some representative autoradiograms of the bands are shown in Figure 1C. In all samples analyzed, HSL antibody stained multiple protein bands at different molecular mass. An extract of protein from rat white adipose tissue (A) was used as an internal standard for HSL expression.
In normal rat ovary (Figure 1C), HSL-immunoreactive proteins were observed at ∼84, 67, 54, and 43 kDa. The intensity of the 84- and 67-kDa bands did not change in the ovaries during the different phases of the estrus cycle. However, the 54-kDa band was more intensely stained in rat ovary at proestrus (P), estrus (E), metestrus (M), and diestrus (D) compared with the prepuberal (PP) rat ovary.
HSL Location and Protein Expression in the Rat Oviduct
We studied the distribution of HSL in the oviduct of non-pregnant rats in the different phases of the estrus cycle. The patterns of HSL immunostaining in the tubaric ampulla and tubaric isthmus of the oviduct during the different phases of the ovary cycle are summarized in Table 2. In all oviducts examined, HSL was mainly found in the luminal epithelium, which contained two main cells types: ciliated and non-ciliated cells. The ciliated cells were identified by the presence of kinocilia on the apical cell surface. The apical surface of non-ciliated cells was either at the same level as the ciliated surface or protruded into the oviduct lumen (Figure 1D). In adult rats, HSL immunoreactivity was found in the utero-tubal junction, preampulla or fimbriae, itshmus, and ampulla. In the tubaric ampulla (Ta; Figure 1D, left), HSL immunoreactivity was found in the nucleus (n) of the epithelial cells and the cilia (c) of the ciliated cells during the different phases of the estrus cycle. In the tubaric ampulla, HSL staining was observed in the nucleus and the cilia during proestrus and estrus (Figure 1D, Ta-P and Ta-E). However, the ampulla cilia were less stained for HSL during metestrus and diestrus (Figure 1D, Ta-M and Ta-D), the nucleus remaining intensely stained for HSL during these phases of the ovarian cycle. In the tubaric isthmus (Ti; Figure 1D, right), HSL immunolabeling can be seen in the cytoplasm of epithelial cells and in the nucleus (n) and the cilia (c) of the ciliated cells during proestrus (Figure 1D, Ti-P). HSL staining disappeared from the nucleus and was maintained in the cilia during estrus, metestrus, and diestrus (Figure 1D, Ti-E, Ti-M, and Ti-D).
Hormone-sensitive lipase (HSL) localization and protein expression in the ovary, corpus luteum, and oviduct during the different phases of the ovarian cycle. F, ovary follicles.
Immunoreaction pattern to hormone-sensitive lipase in the oviduct, uterus, and vagina in different phases of the rat estrus cycle

−, no staining; +, weak staining; ++, moderate staining; +++, intense staining.
In summary, in the tubaric ampulla, immunostaining for HSL was mainly found in the nucleus of the epithelial cells during the different phases of the ovarian cycle. However, in the tubaric isthmus, HSL staining occurred mostly in the cilia.
HSL-immunoreactive proteins in the rat oviduct were observed at ∼67, 54, 43, and 33 kDa (Figure 1E). The 84-kDa band was absent in the rat oviduct. The 67-kDa band was expressed in the rat oviduct, and its intensity did not show any changes at proestrus (P), estrus (E), metestrus (M), and diestrus (D) (Figure 1E). Cyclic variations were observed in the 43-kDa band. In the estrus phase, the oviduct clearly showed less accumulation of the 43-kDa HSL protein than in the proestrus and prepuberal (PP) stages, whereas there was an increase in the intensity of this band at metestrus compared with the proestrus and prepuberal stages. A similar pattern was observed for the 33-kDa band.
HSL Location and Protein Expression in the Rat Uterus and Vagina
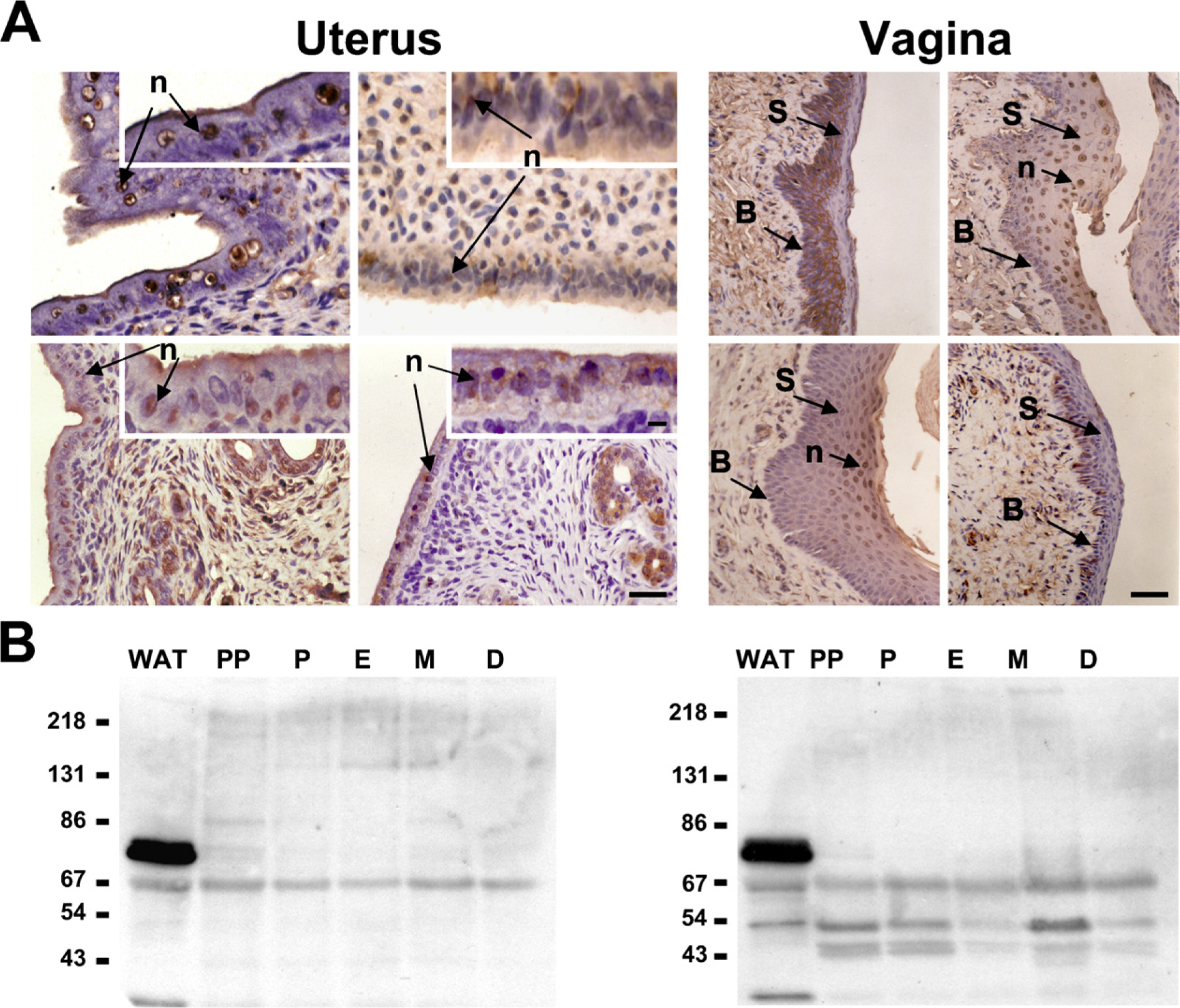
The patterns of HSL immunostaining in the rat uterus and vagina during the different phases of the ovary cycle are summarized in Table 2. Cyclic variations were observed in the epithelial cells of the uterus (U; Figure 2A, left). The epithelial cells of the uterus showed HSL immunostaining mainly restricted to the nucleus (n). Most of the cells of the luminal and glandular epithelia were immunopositive for HSL during diestrus and metestrus (Figure 2A, U-D and U-M), whereas less epithelial cells were immunopositive for HSL during proestrus and estrus (Figure 2A, U-P and U-E). Moreover, many stromal cells (fibroblasts, vascular endothelial cells, lymphocytes, and macrophages) were stained for HSL, and in U-D, the immunoreaction in the stroma is lower compared with the other phases of the estrus cycle (Figure 2A, U-D). The smooth muscle cells of the uterus were stained for HSL (Figure 2A, U-M and U-D).
Cyclic variations were observed in the cells of the vagina (V; Figure 2A, right). The cells of the basal (B) strata and first layer of the spinous (S) strata in the vagina showed HSL immunostaining, mainly restricted to the cytoplasm during proestrus and diestrus (Figure 2A, V-P and V-D). Immunostaining for HSL disappeared, however, from cells of the basal strata, and staining for HSL appeared in the nucleus (n) of the cells of the spinous strata during estrus and metestrus (Figure 2A, V-E and V-M).
In the rat uterus, which we have described previously, the epithelial cells showed HSL immunostaining mainly restricted to the nucleus (n); one major immunoreactive protein, 67 kDa in size, was identified by the anti-HSL chicken polyclonal antibody (Figure 2B, left). No cyclic changes of HSL expression were found in the uterus.
HSL-immunoreactive proteins in the rat vagina were observed at ∼67, 54, and 43 kDa (Figure 2B, right). The intensity of the 67-kDa band did not change in the vagina during the different phases of the ovarian cycle. However, the intensity of the 54- and 43-kDa bands decreased during estrus (E) and diestrus (D) compared with proestrus (P) and metestrus (M).
In summary, these results indicate that HSL is located in the nucleus of the epithelial cells in different segments of the rat oviduct, uterus, and vagina.
Discussion
It has previously been shown that the rat ovary contains both HSL protein and mRNA (Holm et al. 1987; Kraemer et al. 1993; Lindvall et al. 2004). However, the distribution and expression of HSL in the female reproductive organs have not been previously analyzed in detail.
In this study, we showed that HSL is not evenly distributed in the female reproductive organs but is located in various specific cell types and depends on estrus cycle phases. Thus, HSL immunoreactivity seems to vary in the oocyte and theca cells during follicular development and atresia, in luteinic cells during corpus luteum formation and regression, and in the oviduct and the epithelium of the uterus and vagina during the estrus cycle.
HSL localization and protein expression in the rat uterus and vagina during the different phases of the ovarian cycle. U, uterus.
It is interesting to note that the rat immature oocytes of primordial follicles were unstained, but when the oocytes initiated the maturation process, they became immunoreactive for HSL, and the staining for HSL was distributed in both cytoplasm and nucleus. The presence of HSL in the rat oocytes, once the follicle enters the growing pool, supports a role of HSL in gamete maturation. Osuga et al. (2000) showed that HSL is essential for spermatogenesis because the absence of HSL in the testis of HSL-null mice resulted in a 2-fold increase in testicular cholesterol ester content, as well as severe oligospermia and infertility (Osuga et al. 2000). More recently it has been shown that the generation of transgenic mice expressing the testicular form of hormone-sensitive lipase (HSLtes) in postmeiotic germ cells rescues the male infertility observed in HSL-deficient mice (Vallet-Erdtmann et al. 2004; Wang et al. 2004). It seems, therefore, that HSL is involved in spermiogenesis in the testis, and this study suggests that HSL is also involved in gamete maturation and oogenesis in the ovary.
We found that external theca cells were intensely stained for HSL when follicle development had begun, whereas granulosa cells were unstained. In preantral follicles showing a morphologically distinct theca layer, HSL was found in internal and external theca cells and in granulosa cells. The primary function of ovary theca cells is to synthesize and provide an aromatizable androgen substrate (i.e., testosterone, androstenedione) for granulosa cell estrogen biosynthesis (Bjersing 1967; Erickson et al. 1985). In this regard, theca interstitial cells have been shown to express SR-BI, which allows the supply of lipoprotein-derived cholesterol ester required for androgen production (Wu et al. 2003). In the corpus luteum, which also expresses HSL, steroidogenesis depends on exogenous cholesterol, supplied by lipoprotein and mobilized from lipid droplets by HSL or neutral cholesteryl ester hydrolase (Strauss et al. 1981; Gwynne and Strauss 1982). In the testis, HSL is located in Leydig cells, which are androgen-producing cells (Gwynne and Strauss 1982; Rao et al. 2003). Therefore, in both the testis and the ovary, the cells that use the steroidogenic pathway for the conversion of cholesterol to androgens and estrogens contain HSL, which suggests that HSL is involved in cholesterol ester hydrolysis to provide free cholesterol for the synthesis of steroid hormones in these steroidogenic tissues.
There seems to exist multiple HSL isoforms depending on the tissue. By Western blot, we detected four immunoreactive bands (at ∼84, 67, 54, and 43 kDa) in ovaries from Wistar rats. In ovaries from Sprague-Dawley rats, Kraemer et al. (1993) observed HSL-immunoreactive proteins of 84, 89, 67, and 43 kDa. The mRNA and protein species expressed in the testis are larger at 3.9 kb and 120 kDa, respectively (Holm et al. 1988; Holst et al. 1996; Mairal et al. 2002; Arenas et al. 2004). In the heart, skeletal muscle, placenta, and ovaries of rats, the expressed HSL mRNAs are slightly larger (3.5 kb) than in rat adipose tissue (3.3 kb) (Holm et al. 1988). The purified adipose tissue enzyme has a molecular mass of 84 and 88 kDa on SDS-PAGE in rats (Holm et al. 1988) and humans (Langin et al. 1993), respectively. The 89-kDa HSL isoform has been described in pancreatic β cells, adipocytes, adrenal glands, and ovaries (Lindvall et al. 2004). Little is known about the factors regulating HSL gene expression. However, this pattern of expression suggests that several tissue-specific promoters control the different forms of HSL, because when a gene is expressed in somatic tissues and in germ cells, tissue-specific expression often results from alternate promoter use (Sun et al. 1995). In the mouse, an HSL testis-specific promoter has been identified that contains four regions binding testicular nuclear proteins (Blaise et al. 1999).
The oviduct epithelium consists of two kinds of cells, namely the ciliated and non-ciliated (secretory) cells. Ciliated cells play important roles in the transport of gametes, whereas non-ciliated cells synthesize and release specific products into the oviduct fluid (Abe 1996). These results show HSL expression in both cell types. In non-ciliated cells, HSL was mainly found in the nucleus, whereas in ciliated cells, it was found in the nucleus and the cilia. The presence of HSL in the oviduct could be related to the secretion of this enzyme into the lumen. The secretion of HSL has been previously described by us in lactating rat milk (Martín-Hidalgo et al. 2005). The secretion of others lipases has been reported by Paliwal and De (2007), who described that acid-lipase-like protein expression is markedly induced in lacrimal glands and secreted in tears of hamsters during lactation, and this secretion is repressed by androgens and estrogens. In addition, it has been reported that bile salt-dependent lipase is secreted as a component of the pancreatic juice (Nganga et al. 2000). The secretory products released by the epithelium of the oviduct have special relevance in relation to the physiological events, such as gamete transport, capacitation, fertilization, and early embryo development, which occur in this region (Abe 1996). These results also showed HSL immunostaining in the cilia from the tubaric isthmus. During ciliogenesis, cholesterol is required for plasma membrane formation (Chailley and Boisvieux-Ulrich 1985). Thus, it is suggested that HSL, which exhibits cholesteryl ester hydrolase activity, could provide free cholesterol to the plasma membrane during cilia growth.
An interesting, and not previously reported, finding is the positive reaction to HSL observed in the rat uterine epithelium. We found low positive HSL immunoreactivity in the uterine epithelium in the proestrus and estrus phases, which correspond with high estrogen levels in the plasma, whereas the highest levels of HSL were observed in the metestrus and diestrus phases, which correspond with low estrogen levels in the plasma. Moreover, studies that have investigated the dynamics of the two types of estrogen receptors (α and β) in the female reproductive tract have shown that estrogen receptor-α is the predominant subtype in the uterus and that its expression changes during the estrus cycle (Wang et al. 1999, 2000). Cyclic variations and changes in the cellular localization of HSL were observed in the cells of the vaginal epithelium. During proestrus and diestrus, the cells of the basal strata and first layer of the spinosum strata showed HSL immunostaining mainly restricted to the cytoplasm, whereas during estrus and metestrus, the immunostaining for HSL disappeared from the basal strata cells while being observed in the nucleus of the spinosum strata cells. Western blot analysis of HSL-immunoreactive proteins in rat vagina showed bands at 67, 54, and 43 kDa, whereas the 84-kDa band was absent in the vaginal segment. The intensity of the 67-kDa band did not change in the vagina during the different phases of the estrus cycle. However, the intensity of the 54- and 43-kDa bands decreased during estrus and diestrus compared with proestrus and metestrus. The physiological significance of these results remains to be determined.
Although HSL is thought to be a cytosolic enzyme in different tissues, our findings clearly indicate that there is substantial enrichment of HSL in the nucleus of the epithelial cells in different segments of the rat oviduct, uterus, and oocytes. HSL has been also reported to be present in the nucleus of enlarged spermatids (Osuga et al. 2000; Vallet-Erdtmann et al. 2004) and in neoplastic cells (Arenas et al. 2004). The significance of the nuclear immunostaining observed by light microscopy is unknown. Active cholesteryl esters metabolism has been detected in the nucleus of the hepatocytes (Albi and Magni 2002). Thus, the possibility exists that HSL is involved in the hydrolysis of cholesteryl esters present in the nucleus. Additionally, because HSL has also been shown to hydrolyze retinol esters (Wei et al. 1997), this enzyme could participate in the metabolism of retinoids in the nucleus. These possibilities should be directly evaluated in further biochemical studies.
These results showed that epithelial cells of the uterus, where HSL was mainly restricted to the nucleus, expressed one major 67-kDa protein. This isoform is also expressed in the oviduct and vagina, where we also observed HSL localization in the nucleus of the cells. It may be, thus, speculated that the 67-kDa isoform may correspond to the nuclear HSL form, which is highly expressed in the epithelial cells in different segments of the rat oviduct, uterus, and vagina, cells that are actively proliferating and being renewed during the estrus cycle.
This study describes, for the first time, the distribution of HSL in the rat female reproductive organs. Although the physiological significance of these results remains to be determined, the expression and localization of HSL in the rat female reproductive organs change during the ovarian cycle in a stage-specific and cell-dependent manner, suggesting that this protein is under specific regulation. The presence of HSL in theca and granulosa cells of normal rat ovaries suggests that cholesteryl ester activity of this protein may play a key role in steroidogenesis through the supply of free cholesterol for steroid hormone synthesis. This protein is also involved in oogenesis, as its presence in oocytes suggests, supporting the growing body of evidence that HSL may have important roles in the reproductive system and in gamete maturation in female rats. Finally, HSL labeling in the nucleus of epithelial and germ cells suggest an as yet unknown function for this enzyme, probably related to cell proliferation.
Footnotes
Acknowledgements
This study was supported by a grant from the Instituto de Salud Carlos III (FIS PI050403) and Centro de Investigacion Biomédica en Red (CIBER) de Fisiopatologia de la Obesidad y Nutricion (CB06/03), an initiative of ISCIII, Spain.
The authors thank Carlos Correa for help in examining the animals and vaginal smears.