
Abstract
It has been shown that adult pancreatic ductal cells can dedifferentiate and act as pancreatic progenitors. Dedifferentiation of epithelial cells is often associated with the epithelial-mesenchymal transition (EMT). In this study, we investigated the occurrence of EMT in adult human exocrine pancreatic cells both in vitro and in vivo. Cells of exocrine fraction isolated from the pancreas of brain-dead donors were first cultured in suspension for eight days. This led to the formation of spheroids, composed of a principal population of cells with duct-like phenotype. When cultivated in tissue culture-treated flasks, spheroid cells exhibited a proliferative capacity and coexpressed epithelial (cytokeratin7 and cytokeratin19) and mesenchymal (vimentin and α-smooth muscle actin) markers as well as marker of progenitor pancreatic cells (pancreatic duodenal homeobox factor-1) and surface markers of mesenchymal stem cells. The switch from E-cadherin to N-cadherin associated with Snail1 expression suggested that these cells underwent EMT. In addition, we showed co-expression of epithelial and mesenchymal markers in ductal cells of one normal adult pancreas and three type 2 diabetic pancreases. Some of the vimentin-positive cells were found to coexpress glucagon or amylase. These results point to the occurrence of EMT, which may take place on dedifferentiation of ductal cells during the regeneration or renewal of human pancreatic tissues.
Keywords
T
Some authors claim that ductal cells can also transdifferentiate, in particular, into insulin-secreting cells (Bouwens 1998; Bonner-Weir et al. 2008; Inada et al. 2008). This has been demonstrated in models of duct ligation (Wang et al. 1995), streptozotocin-induced type 1 diabetes (Dutrillaux et al. 1982; Nagasao et al. 2003; Park et al. 2007) or sub-total pancreatectomy (Sharma et al. 1999), and in pathological conditions such as nesidioblastosis (Hollande et al. 1976). A similar mechanism has been observed in vitro for rodent (Katdare et al. 2004; Leng and Lu 2005; Kim et al. 2006) or human ductal cells (Bonner-Weir et al. 2000; Yatoh et al. 2007).
The switch in pancreatic cell phenotype may require several steps including dedifferentiation, proliferation, and redifferentiation (Bonner-Weir et al. 2008; Inada et al. 2008), but direct transdifferentiation with mixed cells has also been described (Bertelli and Bendayan 1997). Dedifferentiation of epithelial cells has often been associated with the epithelial-mesenchymal transition (EMT) (Saika et al. 2004; Ulianich et al. 2008; Godoy et al. 2009). EMT plays a key role in embryogenesis [reviewed in Moustakas and Heldin (2007)], but is also involved in some diseases such as renal (Okada et al. 2000) or pulmonary fibrosis (Willis et al. 2005) and in some invasive cancers [reviewed in Thiery (2002); Moustakas and Heldin (2007)]. A recent study showed that induction of EMT in immortalized human mammary epithelial cells generates stem cells (Mani et al. 2008). EMT may also be involved in tissue regeneration, particularly in the liver (Sicklick et al. 2006). In the pancreas, in vitro adult human β-cells have been shown to undergo EMT before redifferentiating into insulin-producing cells (Gershengorn et al. 2004; Ouziel-Yahalom et al. 2006). Although a matter of subsequent debate (Chase et al. 2007; Davani et al. 2007), a recent study using a genetic lineage-tracing method has confirmed the occurrence of EMT in cultured human β-cells (Russ et al. 2009).
In this study, we investigated the occurrence of EMT during transdifferentiation/dedifferentiation of adult human exocrine pancreatic cells both in vitro and in vivo. We showed that, when maintained successively under different culture conditions, these cells still expressed epithelial markers and started to express mesenchymal markers, providing evidence of EMT. In support of these in vitro findings, we showed that, in normal and type 2 diabetic adult human pancreases, some ductal and centroacinar cells coexpressed epithelial and mesenchymal markers. Moreover, some of these mixed epithelial-mesenchymal cells were found to express glucagon or amylase, suggesting the involvement of partial EMT in the renewal of both endocrine and exocrine cells in adult human pancreas.
Materials and Methods
Isolation of Human Exocrine Pancreatic Cells
Primary cell cultures were isolated from exocrine-enriched preparations following islet purification. Human pancreases (
Culture of Human Exocrine Pancreatic Cells
Cells were successively maintained under three culture conditions. First, on day 2 post-isolation (day 0 of culture), small clusters of exocrine cells were cultured in RPMI 1640 (Gibco) supplemented with 20% FCS in non-tissue culture-treated dishes at 37C in 5% CO2-95% air. The medium was renewed every 2 days. Second, tissue cultures were made by seeding 8-day-old spheroids formed in suspension cultures into tissue culture-treated flasks (Nunc; Roskilde, Denmark). They were maintained in RPMI 1640 supplemented with 10% FCS at 37°C in 5% CO2-95% air. Finally, subcultures were obtained after dissociating subconfluent epithelial cell sheets (6- to 8-day-old) with a solution of trypsin (0.05%)-EDTA (0.02%; Gibco). Cells were seeded at a concentration of 7.5 × 10 4 cells/ml in a mixture of DMEM (50%)-Ham F12 (50%) supplemented with 10% FCS, 1% penicillin-streptomycin-fungizone, 1 mM nicotinamide, and 1% insulin-transferrin-selenium (Gibco). Cells were maintained during 5 passages. Cells cultured without antibiotics were checked routinely for any possible contamination with mycoplasma by PCR or by seeding the cells on specific medium.
Growth Assays
For the growth assays, cells were seeded in Petri dishes at 7.5 × 10
4
cells/ml, and the medium was changed daily. Cell proliferation was measured every day (days 1–10) by counting cells in a Malassez hemocytometer. To check the non-tumorigenicity of pancreatic cells maintained in cell cultures, heterotransplantations into nude mice were carried out. Human pancreatic cells (4 × 10
6
, 6-day-old, passage 4) were injected subcutaneously into the dorsal region of 6-week-old female Swiss nu-nu mice (
Transmission Electron Microscopy
To determine the ultrastructure of pancreatic cells, 8-day-old spheroids were fixed with glutaraldehyde (2.5%)-paraformaldehyde (PFA; 2%) in cacodylate buffer (0.1 M; pH 7.2) for 1 hr at 4C, post-fixed with 1% osmium tetroxide (45 min; 4C), dehydrated and embedded in epon-araldite resin. Ultra-thin sections were stained with uranyl acetate and lead citrate and then examined in a Hitachi H600 electron microscope (Tokyo, Japan).
Immunocytochemistry
Immunocytochemical reactions were carried out on sections of spheroids or scraped cell sheets fixed in Duboscq-Brasil solution or PFA (3%) and on cells in tissue cultures or subcultures fixed in situ with PFA (3%). Some reactions were carried out on sections of pancreases from normal (
For whole preparations, after blocking nonspecific antibody binding sites with 1% BSA (Sigma; St Louis, MO) in PBS, preparations were incubated overnight with primary antibodies at the indicated dilutions (Table 1). After washing in PBS, cells were incubated with either goat anti-mouse or anti-rabbit IgG immune serum coupled to a dextran polymer carrying peroxidase (EnVision System; DakoCytomation, Glostrup, Denmark) or with goat anti-mouse (1:100 dilution), anti-rabbit (1:300 dilution), or anti-guinea pig IgG (1:100 dilution) coupled to FITC or tetramethylrhodamine-isothiocyanate (TRITC) (Chemicon; Hampshire, UK). The peroxidase activity was revealed using 3-amino-9-ethylcarbazole or DAB as substrate in the presence of H2O2. Some immunoperoxidase preparations were counterstained with Harris hematoxylin (Sigma). Double labeling reactions [amylase/cytokeratin 7 (CK7), amylase/CK19, amylase/pancreatic duodenal homeobox factor-1 (PDX1), CK7/PDX1, CK7/vimentin, CK19/vimentin, CK7/Snail1, vimentin/amylase, vimentin/glucagon] were performed on sections and on cells in tissue cultures and subcultures. After blocking nonspecific antibody binding sites with 1% BSA, preparations were first incubated with a mix of the primary antibodies. The antigen-antibody complexes were revealed after incubation with a mix of fluorochrome-conjugated goat anti-rabbit and goat anti-mouse IgG. The immunofluorescence reactions were observed in a laser scanning confocal microscope (LSM 410; Carl Zeiss, Oberkochen, Germany) equipped with an argon laser (488 nm) for FITC excitation and a helium laser (543 nm) for TRITC excitation. Cell counts were performed on a minimum of 1500 cells per sample, and percentages were calculated as number of positive cells per 1500. The following controls were used: (a) reactions on sections of human pancreas as positive controls and (b) reactions with only the secondary antibody coupled to peroxidase, FITC, or TRITC.
Primary antibodies
Immunophenotyping by Flow Cytometry
Cell surface antigens were analyzed by flow cytometry to detail the phenotype of human pancreatic cells 24 hr post-isolation, after 5 days in suspension culture, and in cell cultures (passage 5). Briefly, cells from exocrine clusters, spheroids, or cell cultures were dissociated using trypsin-EDTA, washed, and kept for 1 hr at 37C in medium culture without FCS before incubating with antibodies. For each antigen, 2 × 10 5 cells were incubated (30 min, 4C) in 100 μl PBS-0.5% BSA-2 mM EDTA containing the following fluorochrome-conjugated anti-human antibodies (1:10 dilution): CD13-allophycocyanin (APC), CD16-FITC, CD29-phycoerythrin (PE), CD34-peridin chlorophyll protein (PerCP), CD44-PE, CD45-PerCP, CD49b-FITC, CD90-FITC, CD166-PE (all from BD Biosciences; Heidelberg, Germany), CD105-APC (CALTAG Laboratories; Burligame, CA), or the respective isotype controls (1:20 dilution; BD Biosciences). The labeled cells were analyzed by multiparameter flow cytometry using a FACSCalibur flow cytometer and the CellQuest Pro software (BD Bioscience).
Results
Characterization of Human Pancreatic Cells Maintained in Suspension
The small fragments of pancreatic exocrine tissue obtained after digestion and separation of islets were maintained initially in suspension in bacteriological culture dishes. They rapidly aggregated to form spheroids with smooth outlines measuring 100–380 μm in diameter. The immunocytochemical reactions performed on sections of 8-day-old spheroids showed that they were mostly formed of three cell populations:
A principal population representing 75–82% of the cells and displaying CK19 (Figure 1A), CK7 (Figure 1B), and carbonic anhydrase II (CA II) (Figure 1C) immunoreactivity. Most of these cells have shown a nuclear PDX1 immunoreactivity (Figure 1D, arrows). The presence of CK7+/PDX1- cells (Figure 1D, small arrows) and CK7-/PDX1- cells at the periphery of spheroids (Figure 1D, arrowheads) is worth noting. None of the spheroid cells were found to coexpress CK19 and vimentin (Figure 1E). Electron microscopic examination showed an organization of duct-like structures in which the cells were associated by junctional complexes (Figure 1F, arrows) and desmosomes (Figure 1F, arrowheads). They presented digitations along their lateral membranes and some microvilli at their apical poles, and they were generally polygonal in shape. Their cytoplasm contained numerous free polysomes and poorly developed rough endoplasmic reticulum (RER), and their nuclei displayed small amounts of heterochromatin.
A population of amylase-positive cells representing 12–19% of the spheroid cells. Double labeling reactions showed that a part of these cells coexpressed CK19 (Figures 2A–2C, arrows) or CK7. These cells were mainly located in the border of spheroids. These mixed cells suggest the occurrence of acinoductal transdifferentiation, also supported by the presence of cells displaying some ultrastructural characteristics of acinar cells (Figure 2D). The latter presented an abundant RER and Golgi apparatus with dilated cisternae, lysosomes, but no secretory granules. They presented apical microvilli and were joined by terminal bars (Figure 2D, arrow) and desmosomes (Figure 2D, arrowheads). These cells were assumed to be acinar cells undergoing transdifferentiation. It is likely that they were not detected by ICC using the anti-amylase antibody and were, therefore, not accounted for in the estimation of acinar cells. The presence of amylase+/CK19- cells was observed (Figures 2A–2C, arrowhead). Some amylase-immunoreactive cells displayed nuclear PDX1 labeling (Figure 2E, arrows), whereas others were devoid of PDX1 (Figure 2E, arrowheads).
A minority of vimentin-positive cells (3.5%). These cells were grouped in small clumps within the spheroids and did not display CK19 (Figure 1E) or CK7 labeling. Finally, it should be noted that no CA IV-immunoreactive cell was observed in 8-day-old spheroids, and only a small number of endocrine cells (3%) were detected.
Characterization of Human Pancreatic Cells Maintained in Tissue Culture
The majority of spheroids placed in tissue culture-treated flasks adhered to the substrate during the first 24 hr after explantation, rapidly resulting in growth areas following cell migration (Figure 3A). In 2- to 11-day-old cultures, the growth areas were mostly formed of epithelial cells. Some cells, often isolated, had a fibroblast-like appearance (Figure 3A, arrows). Immunocytochemical reactions showed that, in 3- to 9-day-old growth areas, most cells (average 97%) expressed CK7 (Figures 3B and 3C) and CK19 (Figure 3D). In the polygonal-shaped cells, CK7-immunoreactive filaments were seen as a dense network located in the perinuclear cytoplasms, irradiating to the cell periphery (Figure 3C, arrowheads). In the fibroblast-like cells, the CK7- immunoreactive filament bundles connected the two poles of the cell (Figure 3C, arrow). Most of the cells in the growth areas, whether of epithelial or fibroblast appearances, displayed nuclear PDX1 immunoreactivity (Figures 3E and 3F). Cytoplasmic PDX1 labeling was observed in a few cells (Figures 3E, arrows, 3G and 3H, arrows). Double labeling reactions showed vimentin and CK7 coexpression in 46–85% of cells, depending on the growth areas (Figures 4A–4F). In young growth areas, foci of cells coexpressing CK7/vimentin were located either at the periphery (Figure 4A, arrowhead) or in central regions (Figures 4A, arrows, and 4B) of growth areas. In older growth areas, when cells were well spread out, most of them coexpressed CK7 and vimentin. The vimentin filaments were located at the periphery of cells, whereas CK7 filaments were dispersed throughout the cytoplasm (Figure 4C). Various stages of vimentin appearance in CK7-immunoreactive cells were observed (Figures 4D-4F). First, cells displayed one (Figure 4D, arrow) or several (Figure 4E, arrows) foci of vimentin-positive filaments. At later stages, vimentin and CK7 filaments were dispersed throughout the cytoplasm and were often colocalized (Figure 4F). A very small proportion of cells (average 2%) expressed only vimentin (not shown). The acquisition of vimentin expression in CK7-positive cells suggests that these cells are capable of EMT. In this respect, we investigated the expression of Snail1, one of the transcription factors that regulate EMT. Double labeling reactions showed that, in young tissue cultures, when cells emerge from the explant, Snail1 was not expressed in CK7-positive cells (Figure 4G). In older cultures, when cells were spread out in growth areas, nuclear Snail1 immunoreactivity was observed in the majority of CK7-positive cells (Figure 4H). It should be noted that some CK7-immunoreactive cells were devoid of Snail1 (Figure 4I, arrow).
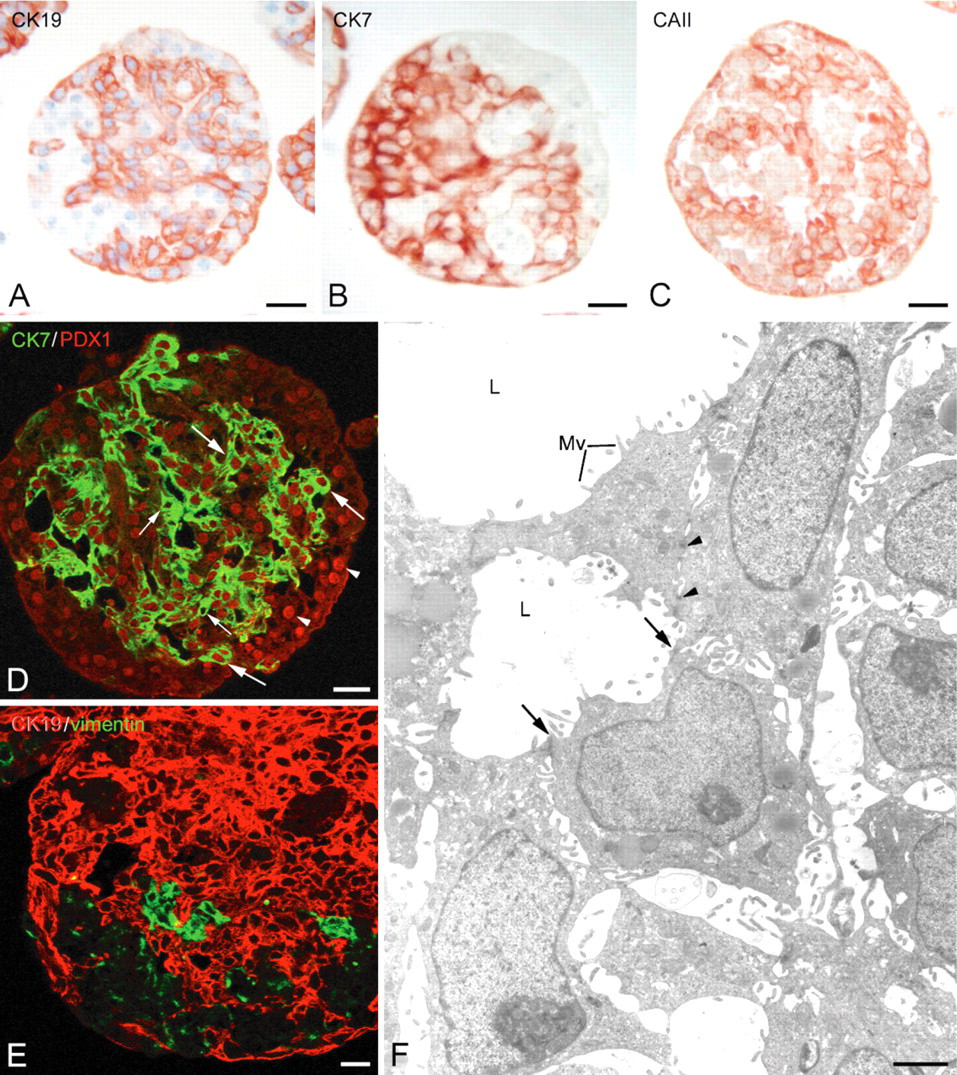
Immunocytochemical characterization of cells with duct-like phenotype in 8-day-old spheroids. (A-C) Immunoperoxidase reactions showing the distribution of cytokeratin 19+ (CK191+
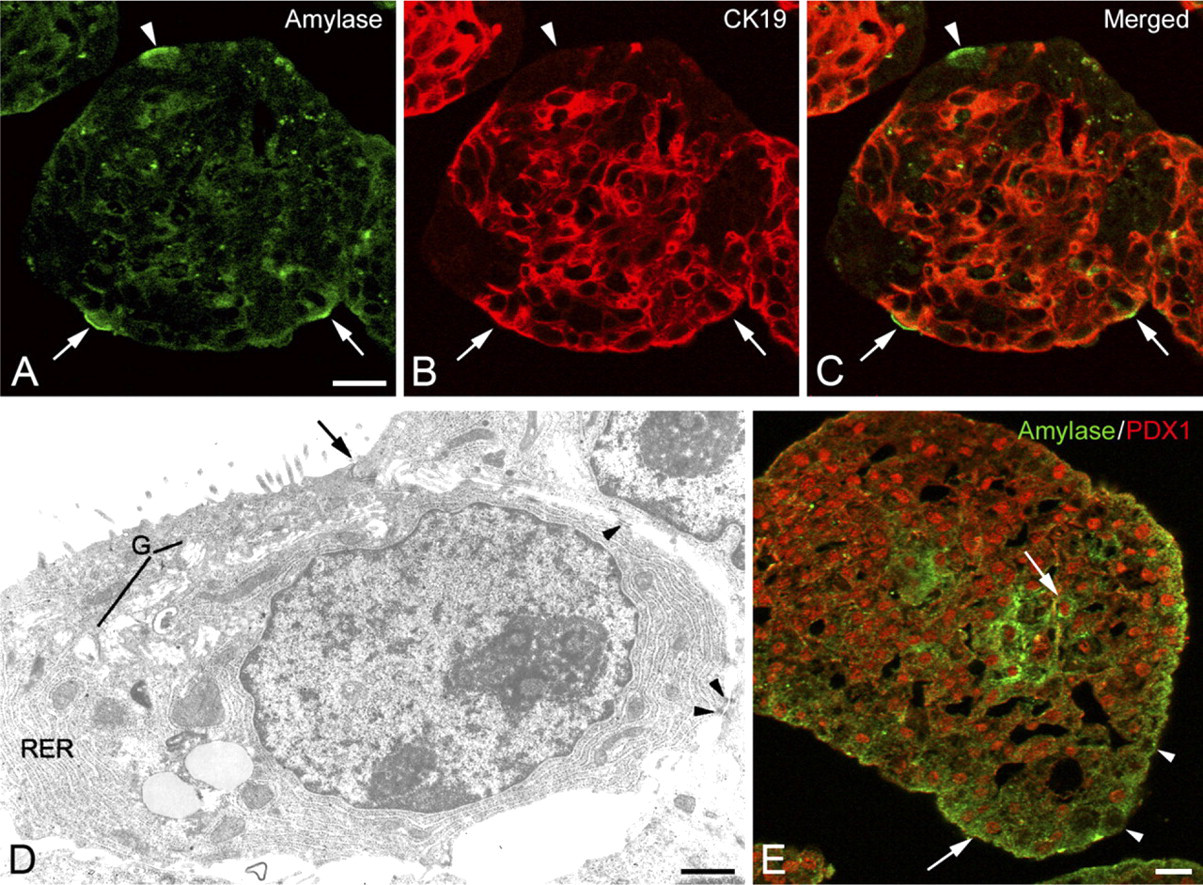
Immunocytochemical characterization of cells with acinar-like phenotype in 8-day-old spheroids. (
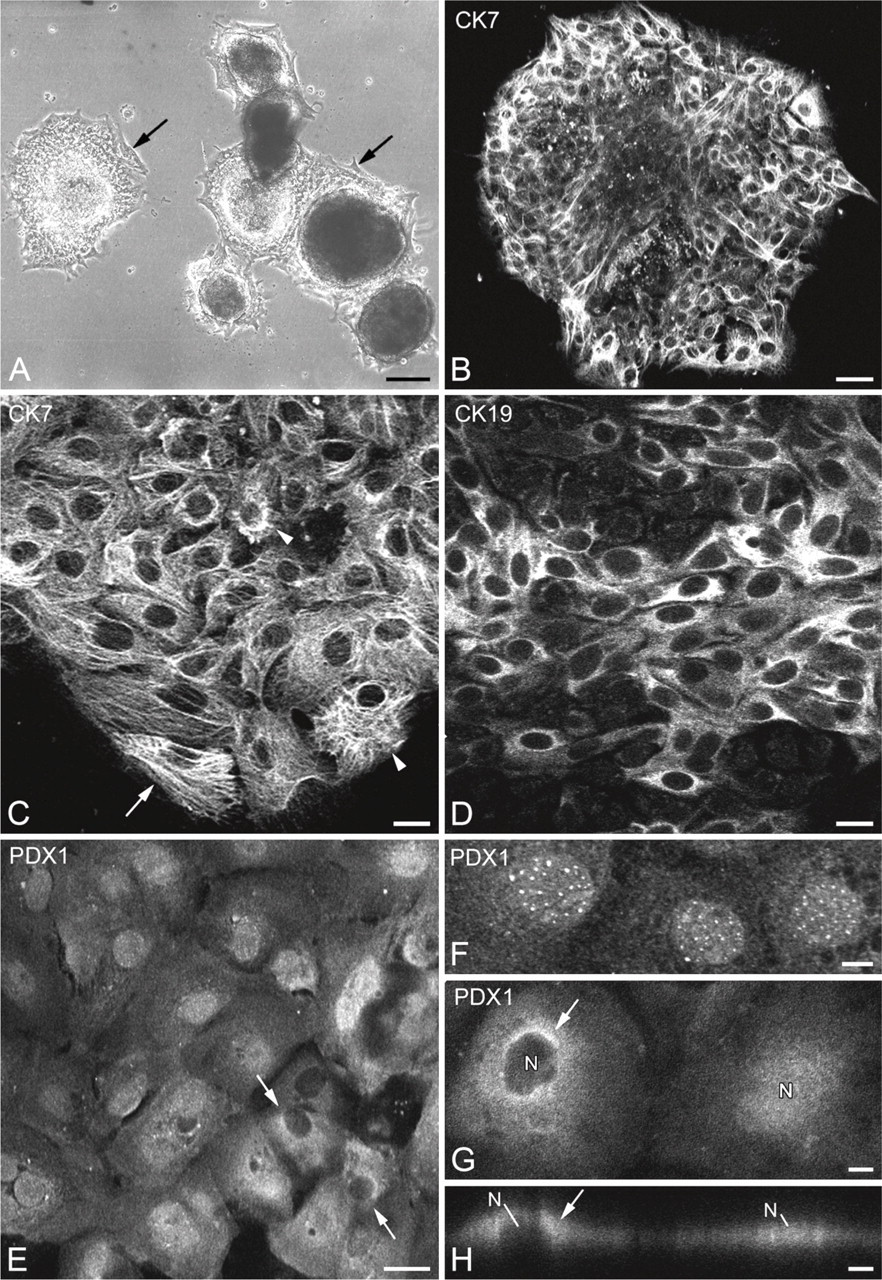
Immunocytochemical characterization of human exocrine pancreatic cells in tissue cultures. (
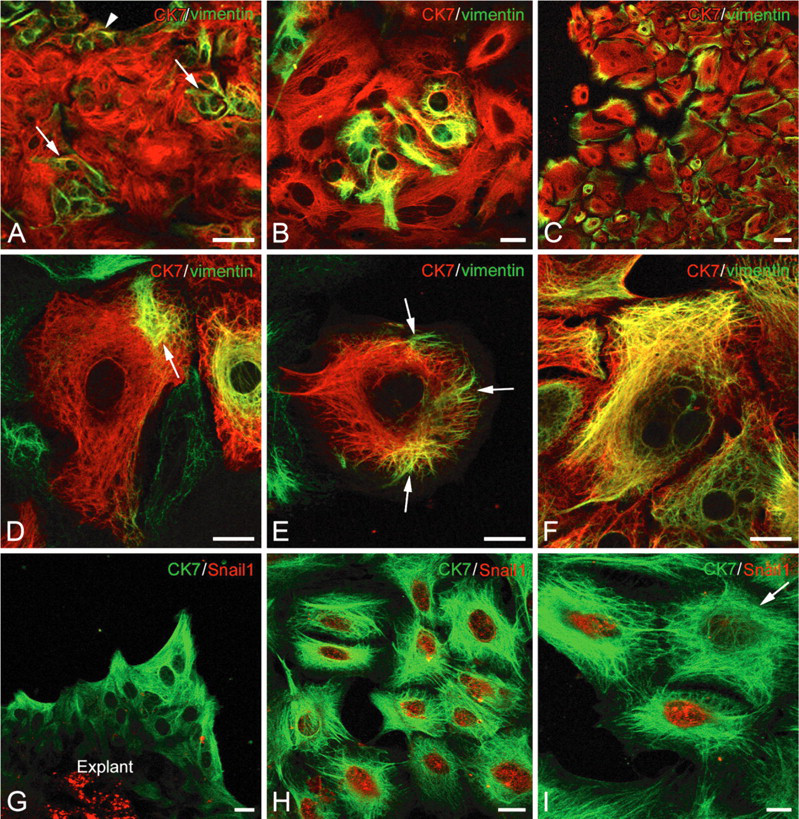
Evidence of epithelial-mesenchymal transition markers in pancreatic cells in tissue cultures. (
Characterization of Human Pancreatic Cells Maintained in Cell Culture
In tissue culture, subconfluence was reached between 8 and 16 days of culture. At this stage, cells were trypsinized and seeded at a concentration of 7.5 × 10 4 cells/ml. From this first passage, cells displayed a fibroblast-like appearance with a narrow cell body and long cytoplasmic prolongations (Figure 5A). Numerous mitoses were observed from day 3 after trypsinization (Figure 5A, arrows). The cells showed proliferation abilities: one passage being performed every 7 days. The doubling time was approximately 36 hr at passage 3. Dorsal subcutaneous heterotransplantation (4 × 10 6 cells/ml) in the nude mouse did not result in transplants, demonstrating that these human pancreatic cells were not tumoral. Immuno-cytochemical reactions carried out between passage 2 and 5 revealed nuclear PDX1 expression in the great majority of cells (Figure 5B). CK7 expression was detected in most cells (average 92%) (Figure 5C). Vimentin (Figure 5D) and α-smooth muscle actin (α-SMA) (Figure 5E) expression was found in all the cells, regardless of passage. All CK7 positive cells coexpressed vimentin (not shown). No labeling was found for desmin, GFAP, and pancreatic hormones (insulin, glucagon, somatostatin, pancreatic polypeptide, or C-peptide).
To confirm that exocrine pancreatic cells underwent EMT in culture, expression of N-cadherin and E-cadherin was examined by immunoperoxidase reaction. N-cadherin immunoreactivity was observed in cells maintained in cell culture (passage 5) (Figure 6A), whereas no E-cadherin labeling was found (Figure 6B). Conversely, E-cadherin expression was detected in spheroid cells (Figure 6C), with no N-cadherin immunoreactivity found (Figure 6D). All the control reactions indicated the specificity of the immunocytochemical results.
Surface marker flow cytometry studies were performed on human pancreatic cells 24 hr after their isolation, cells from 5-day-old spheroids, and cells from adherent cultures (8-day-old, passage 5) using antibodies against mesenchymal stem cell (MSC) markers from bone marrow, adipose tissue, or umbilical cord and hematopoietic cell markers. As shown in Figure 7, cells analyzed 24 hr after their isolation expressed surface antigens CD13 (96.3%), CD29 (99.2%), CD44 (37.9%), CD49b (9.07%), CD90 (4.6%), CD105 (5.7%), and CD166 (84.9%). Expression profile in cells from 5-day-old spheroids showed a slight decrease for CD13 (59.5%) and CD29 (75.9%), similar levels for CD90 (1.7%) and CD105 (5.8%), and an increase for CD44 (94.4%) and CD49b (73.8%). When compared with cells at passage 5, no significant difference was observed for CD13 (99.3%) or CD29 (90.1%), and an increase was observed in CD44 (98.5%), CD49b (55.8%), CD90 (40.6%), and CD105 (70.3%). It is worth noting that CD90 and CD105, the most characteristic markers of MSC, were coexpressed on average in 40% of cells at passage 5 vs only 3% 24 hr post-isolation. Hematopoietic cell surface antigens CD16, CD45, and CD34 were absent. Only CD34 was detected in 3.7% of cells analyzed 24 hr post-isolation. These CD34+ cells, which coexpressed CD31 (not shown), were assumed to be endothelial cells.
Expression of Mesenchymal Markers in Ductal Cells of Adult Human Pancreases
To determine whether exocrine epithelial pancreatic cells also express mesenchymal markers in vivo, immunocytochemical studies were carried out on sections of pancreases from normal and type 2 diabetic brain-dead donors. Immunoperoxidase reactions showed the presence of vimentin-immunoreactive cells in some intralobular ducts of one normal pancreas (Figure 8A, small arrow). These cells were located in the ductal epithelium and were polarized (Figure 8B, arrows); they often contained a large nucleus. Double labeling reactions revealing vimentin (Figures 8C and 8F) and CK19 (Figures 8D and 8G) showed the presence of cells coexpressing vimentin/CK19 in the epithelium of intralobular ducts (Figures 8E and 8H, arrows), providing evidence for the epithelial nature of vimentin-positive cells. Reactions carried out on sections of pancreases from type 2 diabetic donors revealed the presence of vimentin-immunoreactive cells within the exocrine parenchyma of the three cases examined (Figure 9). Double labeling reactions evidenced the presence of vimentin/CK19-immunoreactive cells in intercalated and intralobular ducts (Figures 9A-9C, arrows). Some centroacinar cells were also stained for both vimentin and CK19 (Figures 9D-9F, arrow). To find out whether these epithelial-mesenchymal cells in exocrine parenchyma might be involved in the regeneration of endocrine and/or exocrine tissues, we conducted a series of double labeling reactions. Some vimentin-positive cells located in ductal epithelium exhibited glucagon immunoreactivity (Figures 9G-9I, arrows). In acinar structures, vimentin-positive cells were detected (Figures 9J-9L, small arrow and arrow). Some of them were also labeled with amylase (Figures 9J-9L, arrow).
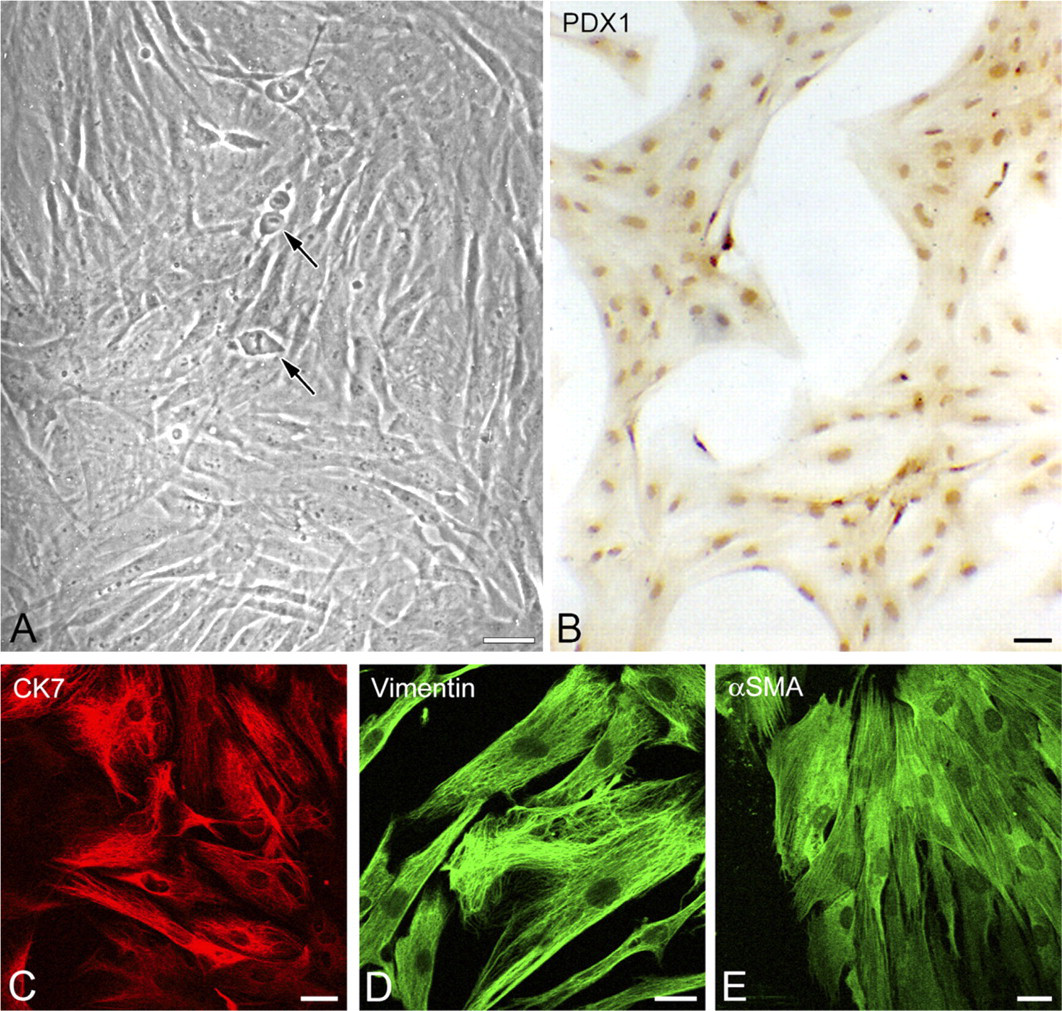
Characterization of human pancreatic cells maintained in cell culture. (
Discussion
We report here that adult human pancreatic exocrine cells maintained under different culture conditions initially dedifferentiate/transdifferentiate, giving rise to duct-like cells. In a second step, they undergo EMT, resulting in a cell population with proliferative capacity and coexpressing epithelial markers, mesenchymal markers, and progenitor cell markers. Moreover, the expression of vimentin along with glucagon or amylase in exocrine epithelial cells in human pancreases suggests the possible involvement of EMT in the tissue regeneration and/or the renewal of pancreatic cells.
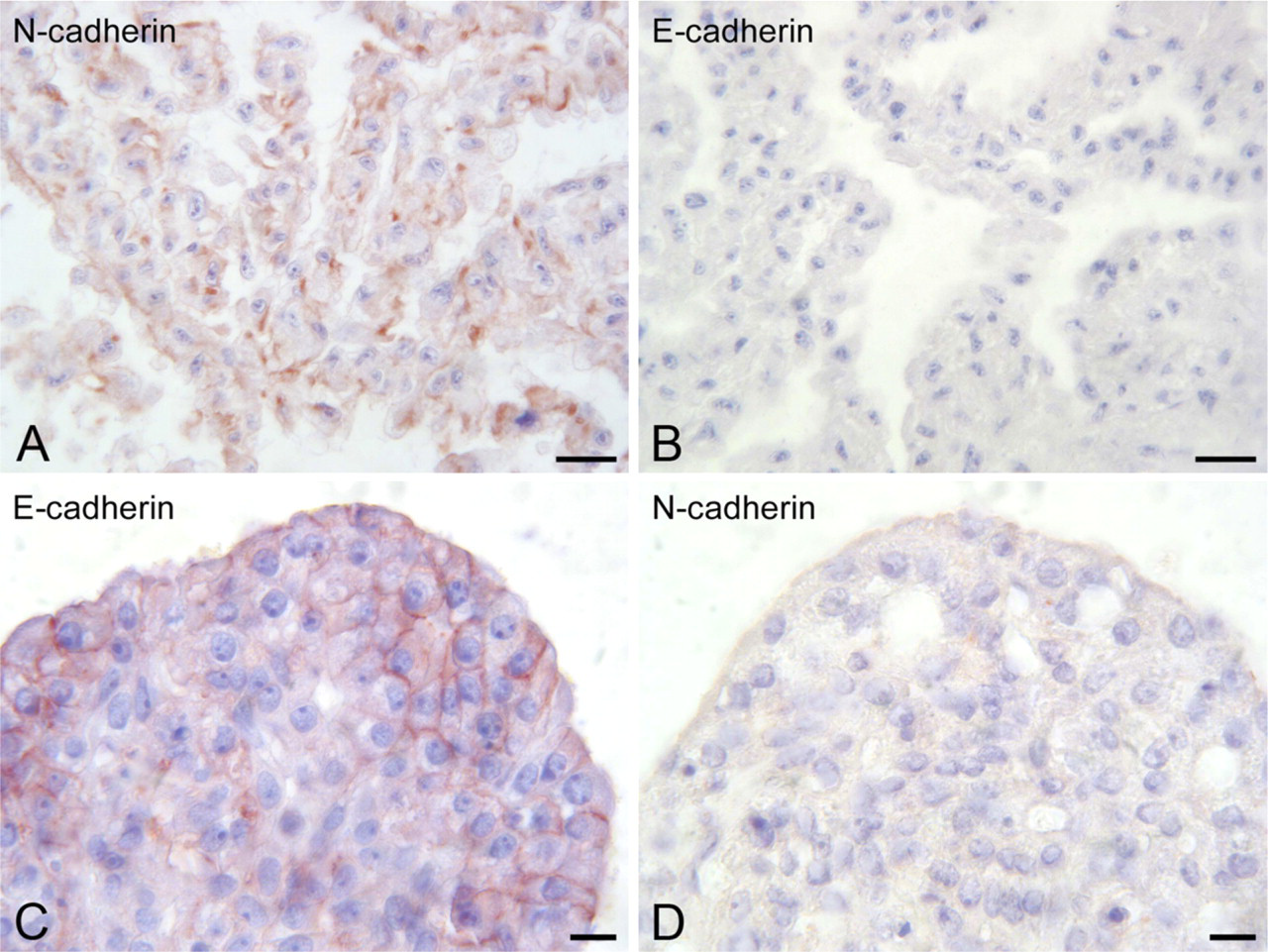
Differential expression of N-cadherin and E-cadherin in exocrine pancreatic cells after immunoperoxidase reactions. (
Culture in suspension of small fragments from adult human pancreatic exocrine tissue consisting of acinar cells, ductal cells (centroacinar and intercalated cells), and a few endocrine cells resulted in spheroid formation from the second day of culture. After 8 days, the principal cell population of spheroids displayed some of the features of adult pancreatic ductal cells, such as CK7, CK19, and CA II expression together with the ability to form duct-like structures by maintaining intercellular junctions. However, these cells no longer express CA IV, enzyme present in adult differentiated ductal cells (Fanjul et al. 2004,2007). They also express proteins, which are absent from mature ductal cells such as PDX1. In view of these features, the cells forming the main population of the spheroids were considered to be duct-like cells. These cells probably had two origins. Some derived from dedifferentiation of ductal cells. Ductal cells, which are mainly intercalated and centroacinar cells, suffered little from apoptosis (data not shown). They evolved during suspension culture into duct-like cells. Moreover, the characteristic ultrastructural appearance of undifferentiated cells and nuclear expression of transcription factor PDX1 are additional arguments supporting a dedifferentiation process. PDX1 is expressed transiently during embryonic development, first in pancreatic progenitor cells located in ducts, which give rise to the different pancreatic cell lineages, and second, during terminal differentiation of β- and δ-cell precursors (Habener et al. 2005). PDX1 is also expressed during pancreatic regeneration (Sharma et al. 1999; Song et al. 1999) and in transdifferentiation of pancreatic (Baeyens et al. 2005; Lipsett et al. 2007) and extra-pancreatic (Yang et al. 2002; Timper et al. 2006; Gao et al. 2008) cells into insulin-producing cells. Other cells in the spheroids were derived from transdifferentiation of acinar cells, although a proportion of the acinar cells underwent cell death by apoptosis and necrosis in the first days after isolation (data not shown). Demonstration by double labeling of mixed cells coexpressing amylase and CK19, making up 7–12% of the cells after 8 days in culture, indicated that some of the acinar cells underwent an acinoductal transdifferentiation. This proportion may be underestimated, because the acinar-like cells devoid of zymogen granule observed on electron microscopy and which are most likely in the process of transdifferentiation were not accounted for, as they were no longer amylase-immunoreactive. These results differ from those of Street et al. (2004), who, using similar culture conditions, did not find mixed cells coexpressing ductal and acinar markers. They concluded that nearly all acinar cells die, thereby selecting duct-like cells. An acinoductal transdifferentiation giving rise to cells with similar expression patterns to those observed in the present study has been observed in vitro for human (Gmyr et al. 2000) and rodent acinar cells (Arias and Bendayan 1993; Rooman et al. 2000; Lipsett et al. 2007) and in animal models of pancreatic regeneration (Wang et al. 1995; Sharma et al. 1999). The expression of PDX1 in amylase-immunoreactive cells is a further argument for acinoductal transdifferentiation. Indeed, PDX1 has been shown to be induced in duct-like cells arising from acinar cells (Rooman et al. 2000). Moreover, it has been shown that persistent PDX1 expression induces acino-to-ductal metaplasia (Miyatsuka et al. 2006).
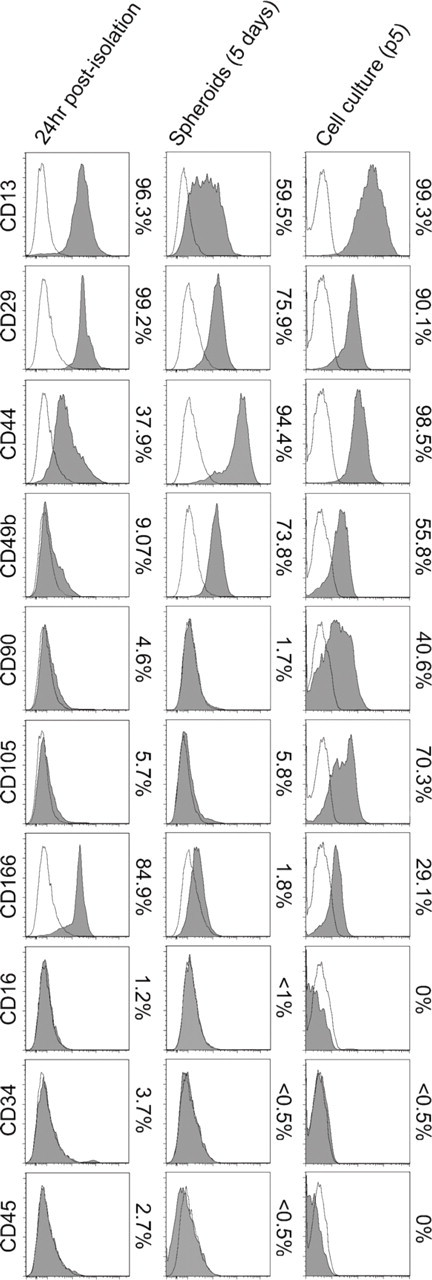
Cell surface antigen-expression profile of human exocrine pancreatic cells analyzed by flow cytometry. Analyses were performed on cells 24 hr after their isolation, after 5 days in suspension culture, and in cell cultures (8 days, passage 5). Plots show specific antibody staining profile (solid) vs isotype control staining profile (line). Percentages represent the ratio of positive cells.
Changing from 3D culture to 2D culture resulted in adhesion of spheroids and migration of the cells. The great majority of these cells were of epithelial type, although some resembled fibroblasts. The expression of CK7/CK19 was preserved, irrespective of morphology. The progressive appearance of vimentin, a protein of the intermediate filaments of cells of mesenchymal origin, was observed during tissue culture. After the first trypsinization, all the cells were fibroblast-like, coexpressing vimentin and CK7. The presence of α-SMA, another cytoskeletal protein of cells of mesenchymal origin, was also found in these cells. These alterations in cell architecture and cytoskeleton are a feature of EMT. It has been shown that vimentin and α-SMA can be synthesized by epithelial cells during EMT, particularly during formation of metastases (Thiery 2002) or diseases such as proliferative vitreous retinopathy (Casaroli-Marano et al. 1999), renal fibrosis (Okada et al. 2000), and idiopathic pulmonary fibrosis (Willis et al. 2005). EMT also leads to disssociation of cells, with alterations in the expression of adhesion molecules. The switch in expression from E-cadherin to N-cadherin that we observed in pancreatic cells maintained in adherent culture is a further indicator of the occurrence of EMT. The fall in E-cadherin is known to be regulated by various transcription factors, including Snail1 (Cano et al. 2000). The demonstration of Snail1 in cells migrating in growth areas is another argument in favor of the occurrence of EMT in human pancreatic exocrine cells in vitro. These findings are consistent with those of Gershengorn et al. (2004) and Russ et al. (2009), who demonstrated presence of EMT in human β cells in vitro. Moreover, flow cytometry studies showed that pancreatic cells in culture express surface markers of MSC from bone marrow, umbilical cord blood, or adipose tissue (Wagner et al. 2005) and do not express surface markers for cells of hematopoietic origin. Although some of these markers were also detected in exocrine pancreatic cells 24 hr after isolation, we noted on passage 5 a marked increase in the proportion of cells expressing CD90 and CD105, both regarded as the most characteristic surface markers of MSC. Nevertheless, the demonstration of multipotency is required to claim that these cells are true MSC. The expansion of pluripotent MSC from cultures of human exocrine pancreas has been described by Seeberger et al. (2006) and Baertschiger et al. (2008). They have considered these cells to stem from proliferation of preexisting MSC in the adult exocrine pancreas. Other authors have considered these cells as stem-like cells and have concluded that they arise from dedifferentiation of mature exocrine cells (Rapoport et al. 2009). This latter interpretation is more consistent with our results, which led us to conclude that the dedifferentiation of adult human exocrine pancreatic cells in vitro occurred via EMT.
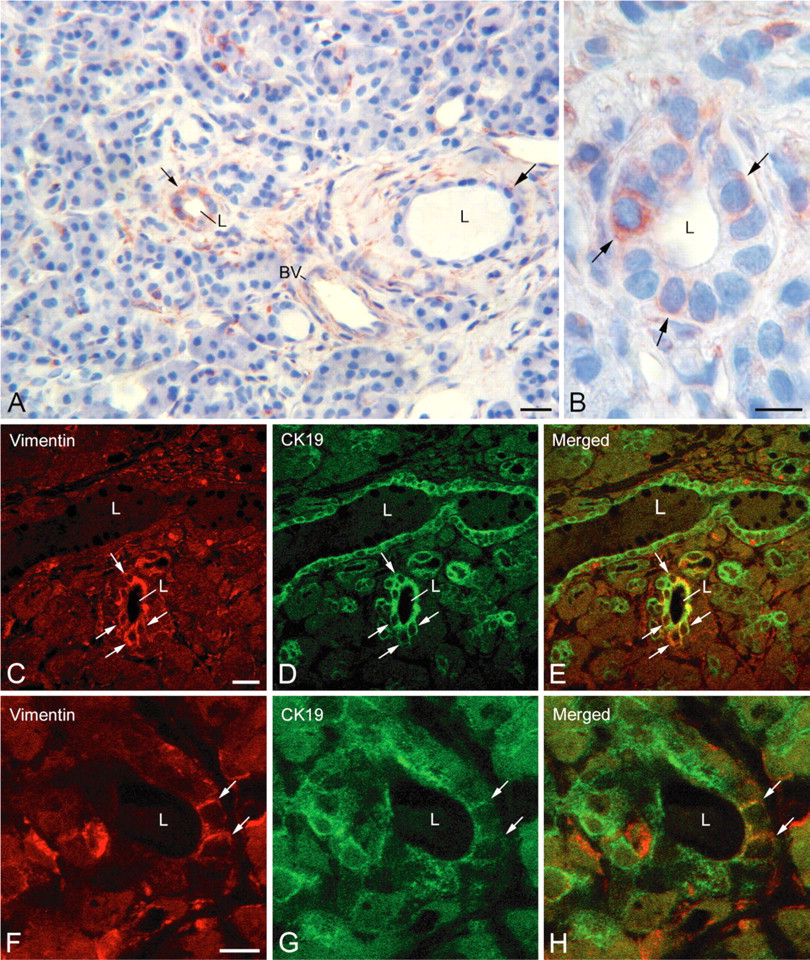
Expression of vimentin in ductal cells of normal human pancreas. (
The experiments we conducted on healthy or diabetic human pancreases showed the presence of mixed epithelial-mesenchymal cells, indicative of a certain extent of EMT in vivo. This conflicts with the study of Seeberger et al. (2009), who failed to find mesenchymal markers in epithelial cells of adult human pancreas and who assumed that EMT was a culture artifact. The presence of mixed epithelial-mesenchymal cells was more common in type 2 diabetic pancreases, as we detected them in all three cases vs one in the normal pancreases. Our results are partly in agreement with those of Ko et al. (2004), who observed cells coexpressing CK19 and vimentin in ductal epithelium in regenerating foci of a pancreas removed from a type 2 diabetic patient with pancreatic adenocarcinoma. Along with regeneration areas, we also observed CK19/vimentin coexpression in intercalated ductal cells, interlobular ductal cells, and in centroacinar cells. These cells may reflect a stage of dedifferentiation of ductal cells toward a progenitor cell, enabling regeneration of exocrine and endocrine pancreatic tissues. In the model of streptozotocin-induced diabetic rat, it has been suggested that the centroacinar cells and cells of intercalated ducts are precursors of β-cells (Nagasao et al. 2003). In human type 2 diabetes, it has been shown that neogenesis of α- and β-cells can occur during regeneration, probably from cells already present in the ductal epithelium (Jones and Clark 2001, Yoon et al. 2003). Our results showing the presence of vimentin-positive cells coexpressing either glucagon or amylase in the exocrine parenchyma of diabetic pancreas argue in favor of the involvement of epithelial cells expressing vimentin in tissue regeneration and/or cell renewal. It is still not clear whether these vimentin-expressing cells correspond to activated progenitor cells undergoing differentiation toward endocrine cells as described in the developing pancreas of rat and mouse embryos (Bouwens and De Blay 1996; Di Bella et al. 2009) or whether they are ductal cells that dedifferentiate to afford progenitor cells that can redifferentiate to an endocrine or exocrine pathway. This latter possibility is argued by our in vitro results showing that human exocrine cells dedifferentiate undergoing EMT (expression of vimentin, α-SMA, PDX1, Snail1, and cell surface antigens of MSC). It is also supported by the observations of Bonner-Weir et al. (2008) and Inada et al. (2008), who have shown after lineage tracing in newborn mouse pancreas or after duct ligature in adult mouse pancreas that it is ductal cells which dedifferentiate to give rise to endocrine and exocrine cells. Taken together these findings argue in favor of dedifferentiation/redifferentiation of epithelial exocrine cells in adult human pancreas. We suggest that, in the adult human pancreas, the vimentin-positive cells in the ductal epithelium correspond to epithelial cells dedifferentiating via EMT for tissue regeneration. Involvement of EMT during regeneration has been described in both normal and diseased liver (Sicklick et al. 2006). A recent study showed the involvement of EMT in the formation of islets of Langerhans from ductal epithelium during embryonic development of sheep and human pancreas (Cole et al. 2009). Nonetheless, these findings do not exclude the possibility of cellular renewal from mesenchymal stem cells of the stroma as suggested by several authors (Seeberger et al. 2006; Davani et al. 2007).
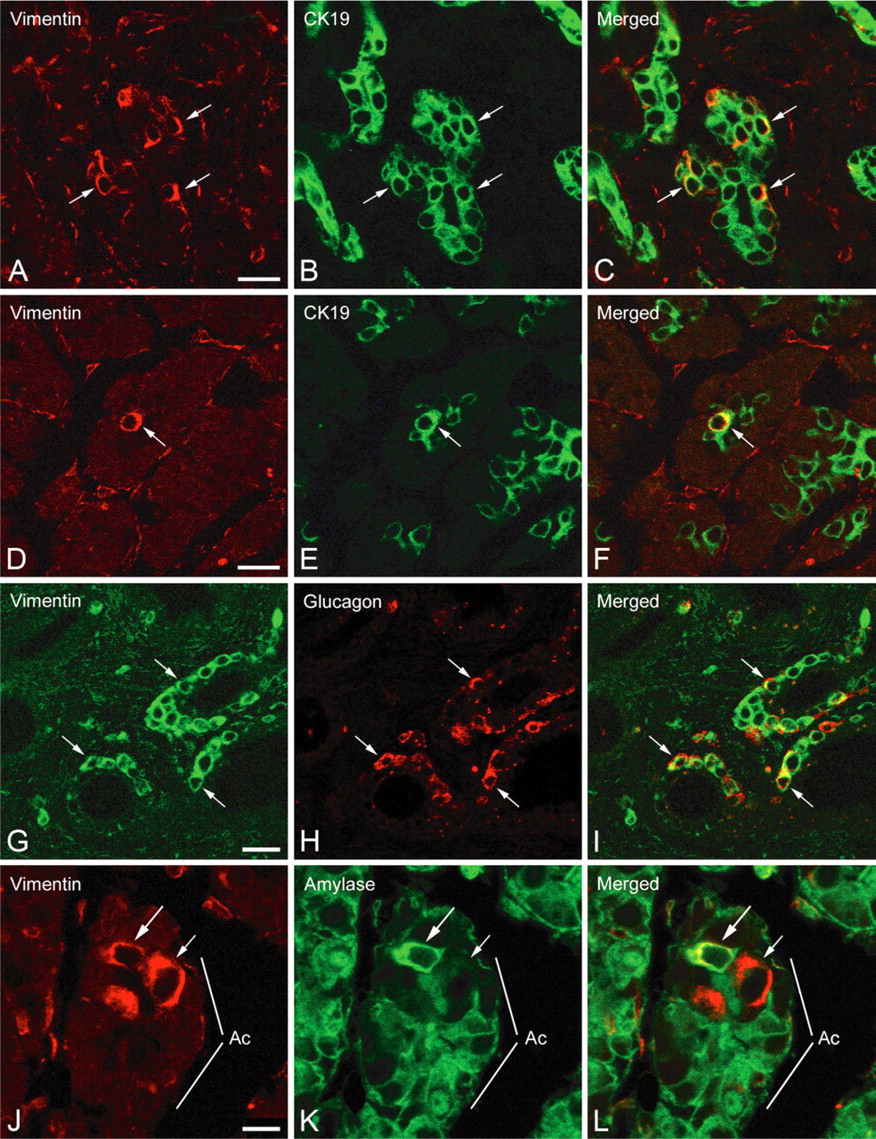
Expression of vimentin in ductal and acinar cells of pancreases from type 2 diabetic donors. (
In summary, the demonstration of cells coexpressing epithelial and mesenchymal markers in human pancreatic ducts argues in favor of the notion that the EMT observed for pancreatic exocrine cells in vitro is not solely a consequence of culture, but may represent a physiological step in the tissue regeneration and/or the renewal of human pancreatic cells.
Footnotes
Acknowledgements
We thank Mr. F. Stéfani and Mr. C. Baritaud for their technical assistance. We acknowledge the use of the confocal microscopy imaging facilities of the Center of Developmental Biology (CBD-IFR109, Paul Sabatier University, Toulouse, France).