
Abstract
Thirteen mammalian aquaporin (AQP) isoforms have been identified, and they have a unique tissue-specific pattern of expression. AQPs have been documented in the reproductive system of both male and female humans, rats, and mice. However, tissue expression and cellular and subcellular localization of AQPs are unknown in the female reproductive system of pigs. In this study, AQP1 immunoreactivity was detected in the capillary endothelium of the ovary. Distinct immunolabeling of capillary endothelium was also observed in the oviduct and uterus. AQP5 was expressed in flattened follicle cells of primordial follicles, granulosa cells of developing ovarian follicles, and muscle cells of the oviduct and uterus. Staining of AQP5 was also observed in the epithelial cells of the oviduct and uterine epithelium. AQP9 immunoreactivity was observed in granulosa cells of developing follicles. AQP9 was also localized in the luminal epithelial cells of the oviduct and uterine epithelia cells. This is, to our knowledge, the first study that shows tissue expression and cellular and subcellular localization of AQPs in the reproductive system of the female pig. Moreover, these results suggest that several subtypes of the AQPs (AQP1, 5, and 9) are involved in regulation of water homeostasis in the reproductive system of gilts.
A
The first confirmation of AQP in the female reproductive system was obtained by isolating and sequencing the cDNA encoding a water channel from the human uterus (Li et al. 1994). Afterward, Li et al. (1997) found AQP1 mRNA in the rat uterus. To date, based on protein expression, at least nine AQP iso-forms have been shown to be expressed in the female reproductive system of humans, rats, and mice (reviewed by Huang et al. 2006). Their specific expression pattern suggests that they play a role in water movement between the intraluminal, interstitial, and capillary compartments. There is also evidence indicating that ovarian steroid can regulate the expression of several AQPs (Jablonski et al. 2003; Branes et al. 2005; Lindsay and Murphy 2006). Nevertheless, data concerning the expression and role of AQPs in female reproductive tissues are still very limited and not available in relation to farm animals.
The aim of this study was therefore to examine the tissue expression and cellular and subcellular localization of AQPs in the reproductive system of gilts.
Materials and Methods
Experimental Animals
All experiments were performed in accordance with the principles and procedures of Animal Ethics Committee of the University of Warmia and Mazury in Olsztyn. Tissue samples were recovered from mature cross-bred gilts on days 17-19 of the estrous cycle (follicular phase). The animals were studied during the third estrous cycle, which was controlled with vasectomized boars, and additionally, the stage of the cycle was confirmed, as previously shown (Akins and Morrissette 1968).
Primary Antibodies
In this study, affinity-purified polyclonal antibodies (SulfoLink Kit; Pierce, Rockford, IL) to the following AQPs were used: AQP1 (Terris et al. 1996), AQP2 (Nielsen et al. 2006), AQP3 (Kim et al. 2005), AQP4 (Vajda et al. 2002), AQP5 (Nielsen et al. 1997), AQP7 (Nejsum et al. 2000), AQP8 (Elkjaer et al. 2001), AQP9 (Carbrey et al. 2003), and AQP11 (Gorelick et al. 2006).
SDS-PAGE and Immunoblotting
After isolation, the tissues were immediately placed in icecold dissection buffer (0.3 M sucrose, 25 mM imidazol, 1 mM EDTA in ddH2O, pH 7.2) containing 8.4 μM leupeptin and 0.4 mM pefabloc (Skowronski et al. 2007). The tissue samples were homogenized using an ultra Turrax T8 homogenizer (IKA Labortechnik; Staufen, Germany) and centrifuged at 4000 × g for 15 min at 4C. The supernatant diluted in SDS buffer contained a final concentration of 62 mM Tris (hydroxymethyl)-amino-methane, 0.1 M SDS, 8.7% glycerol, 0.09 mM bromophenol blue, and 0.04 M dithiothreitol (DTT), pH 6.8. The protein samples were heated for 5 min at 90C and stored in a refrigerator for further analysis.
The samples were warmed to 37C and were loaded into 12.5% polyacrylamide gels, and proteins were separated by electrophoresis. The total protein amount in each sample was adjusted by staining with Gelcode Coomassie Blue Stain Reagent (Bie and Berntsen; Åbyhøj, Denmark) to calculate equal loading. The proteins of studied gels were electrotransferred onto nitrocellulose membranes (Hybond ECL RPN3032D; Amersham Bioscience, Little Chalfont, UK) for 1 hr at 100 V. The membranes were blocked with 5% milk in PBS-T (80 mM Na2HPO4, 20 mM NaH2PO4, 100 mM NaCl, pH 7.5, and 0.1% v/v Tween 20) for 1 hr. After washing, the membranes were incubated overnight at 5C with anti-AQPs antibody.
The membranes were washed and incubated with horseradish peroxidase (HRP)-conjugated goat antirabbit IgG secondary antibody (DAKO; Glostrup, Denmark) in PBS-T for 1 hr. After washing with PBS-T, the sites of antibody-antigen reaction were visualized with HRP-conjugated secondary antibodies (P448, diluted 1:3,000; DAKO) with an enhanced chemiluminescence (ECL) system (Amersham Bioscience) and exposed to photographic film (Hyperfilm ECL, RPN3103K; Amersham Bioscience).
IHC
Tissues were fixed by immersion in 4% paraformal-dehyde for 24 hr (Skowronski et al. 2007). For preparation of paraffin-embedded tissue sections (4 μm thickness), the tissues were dehydrated in ethanol, followed by xylene and finally embedded in paraffin. The sections were dewaxed and rehydrated. For immunoper-oxidase labeling, endogenous peroxidase was blocked by 0.5% H2O2 in absolute methanol for 10 min at room temperature. To reveal antigens, the sections were submerged in 1 mM Tris solution (pH 9.0) supplemented with 0.5 mM EGTA and heated in a microwave oven. After the treatment, the sections were left for 30 min in the buffer for cooling. Nonspecific binding of IgG was eliminated by incubating the sections in 50 mM NH4Cl for 30 min, followed by blocking in PBS supplemented with 1% BSA, 0.05% saponin, and 0.2% gelatin. The sections were incubated overnight at 4C with primary antibodies diluted in PBS supplemented with 0.1% BSA and 0.3% Triton X-100. The sections were rinsed with PBS supplemented with 0.1% BSA, 0.05% saponin, and 0.2% gelatin and incubated with HRP-conjugated secondary antibody (DAKO). Labeling was visualized by 0.05% DAB. The microscopy was carried out using a DMRE light microscope (Leica; Heidelberg, Germany).
Results
IHC analysis using antibodies against nine AQPs (AQP1, 2, 3, 4, 5, 7, 8, 9, and 11) was performed to examine whether these water channel proteins are expressed in the female pig reproductive system. The analysis confirmed the expression of AQP1, AQP5, and AQP9. In contrast, no staining for other examined AQPs (AQP2, 3, 4, 7, 8, and 11) was observed in tested tissues (data not shown). The data obtained are presented in Figures 1-3 and summarized in Table 1.
AQP1 immunoreactivity was detected in the capillary endothelium of the ovary (Figure 1A). An identical labeling pattern was observed in the oviduct (Figure 1B) and uterus (Figure 1C). As a positive control, AQP1 labeling was seen in the apical and basolateral plasma membrane domains in the proximal tubule cells of pig kidney (Figure 1D), consistent with the previous findings in the kidneys of mice, rats, and humans (Nielsen et al. 1995; Maunsbach et al. 1997; Ma et al. 1998). Moreover, immunoblotting showed that anti-AQP1 antibody recognized a 29-kDa band in the ovary (Figure 1F, Lane 1), oviduct (Figure 1F, Lane 2), and uterus (Figure 1F, Lane 3) of the pig.
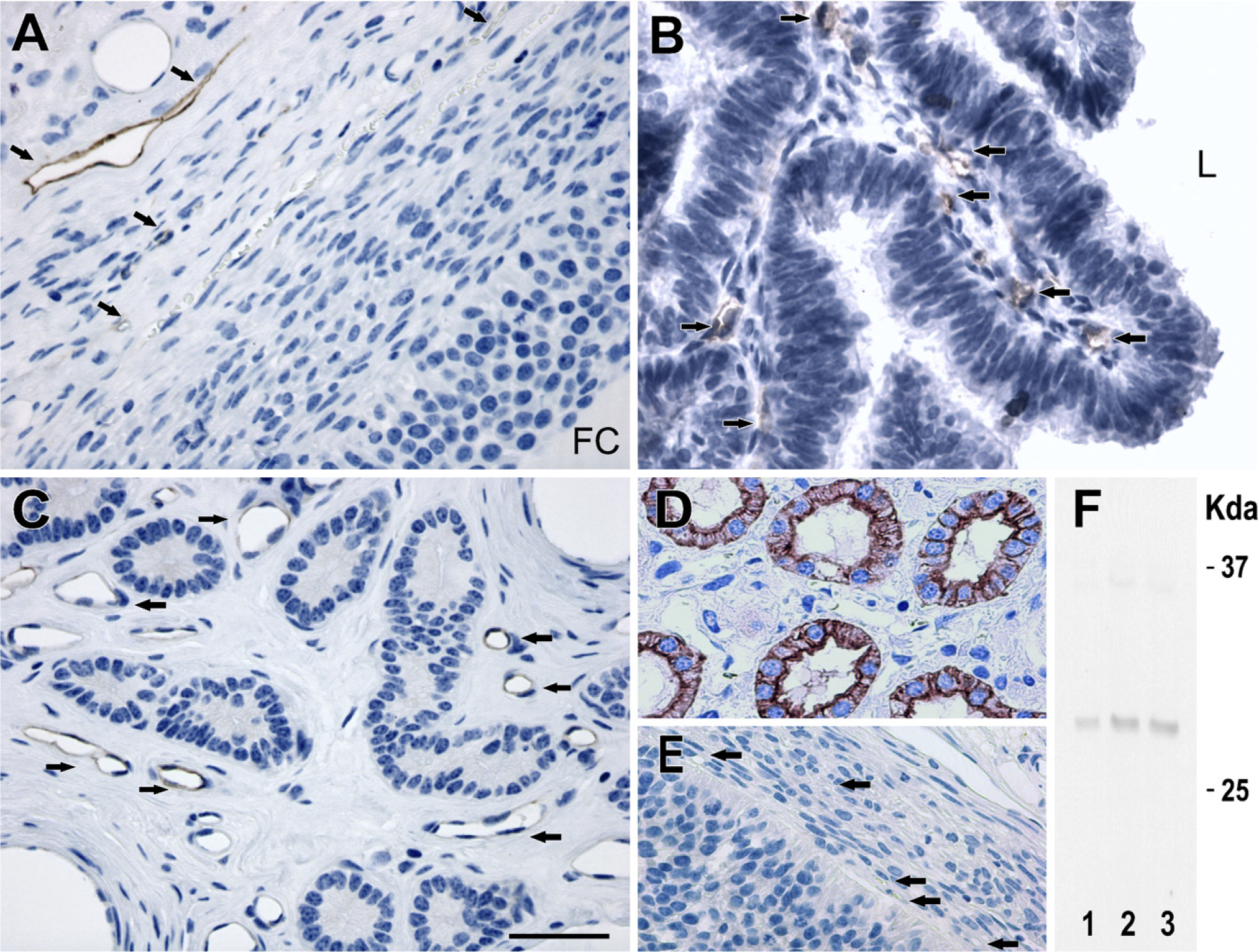
IHC staining of aquaporin (AQP)1 in paraffin-embedded sections of the ovary and uterus from pigs. Anti-AQP1 antibody labels capillary endothelium of the ovary (
AQP5 was localized in granulosa cells and flattened follicle cells of the primordial follicles in the ovary (Figures 2A and 2B). In the oviduct, anti-AQP5 antibody labeled the cells of the muscles layer (Figure 2C) and luminal epithelial cells (Figure 2D). In the uterus, AQP5 was found in the smooth muscle cells (Figure 2E) and luminal and glandular epithelial cells (Figure 2F). As a positive control, AQP5 antibody noticeably stained (Figure 2G) the apical plasma membrane of the type I pulmonary epithelial cells of the pig, consistent with the previous findings seen in the lung tissues of mice, rats, and humans (Nielsen et al. 1997; Krane et al. 2001; Kreda et al. 2001). Imunoblotting showed that anti-AQP5 antibody recognized a 28-kDa band in the porcine ovary (Figure 2I, Lane 1), oviduct (Figure 2I, Lane 2), and uterus (Figure 2I, Lane 3).
AQP9 immunoreactivity was observed in the granulosa cells of the developing follicles in the ovary (Figure 3A). In the oviduct, AQP9 immunoreactivity was detected in the epithelial cells (Figure 3B). The anti-AQP9 antibody labeled the luminal and glandular epithelial cells of the uterus (Figure 3C). As a positive control, in agreement with the previous observation in the liver tissues of mice and rats (Carbrey et al. 2003; Rojek et al. 2007), AQP9 staining was seen at the sinusoidal surfaces of hepatocyte plates in the pig liver (Figure 3D). Immunoblotting also showed that the anti-AQP9 antibody recognized a 32-kDa band in the ovary (Figure 3F, Lane 1), oviduct (Figure 3F, Lane 2), and uterus (Figure 3F, Lane 3) of the pig.
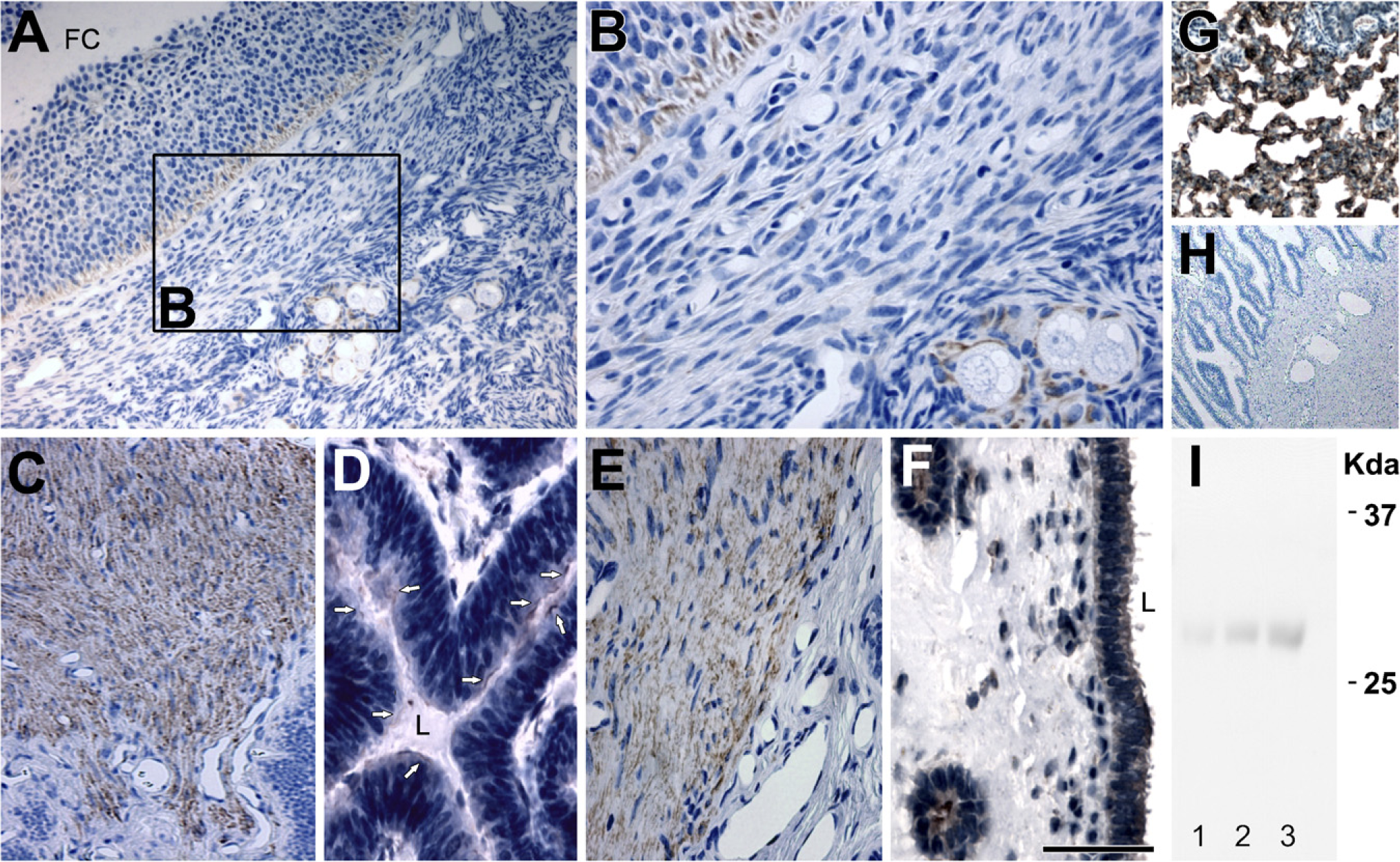
IHC staining of AQP5 in paraffin-embedded sections of the ovary (
Negative controls for all IHC analyses were performed by both omitting the primary antibodies specific to each AQP (Figures 1E, 2H, and 3E) and using non-immune IgG (data not shown), and no specific immunostaining was observed.
Discussion
In this study, IHC and immunoblotting showed that three isoforms of water channel proteins (AQP1, 5, and 9) are expressed in the pig ovary, oviduct, and uterus. In contrast, IHC showed no immunostaining for other examined AQPs (AQP2, 3, 4, 7, 8, and 11).
AQPs in Ovary
This study showed AQP1, 5, and 9 protein expression in the pig ovary: AQP1 in the capillary endothelium; AQP5 in the flattened follicle cells of primordial follicles and the granulosa cells of developing follicles; and AQP9 in the granulosa cells. AQPs have not been extensively studied in the ovary thus far, and this study provides the first evidence of AQP1 and AQP5 expression in ovarian tissues. The AQP9 expression has been previously shown in rat granulosa cells (McConnell et al. 2002); however, we showed the subcellular localization of AQP9 within the granulosa cells. In turn, Edashige et al. (2000) showed the mRNA expression of AQP3 and AQP7 in mouse oocytes. The presence of AQP3 at mRNA level in mouse oocytes was recently confirmed by Meng et al. (2008). In the mammalian ovary, the proper development of oocytes depends on the conditions provided by follicles including follicular fluid accumulation. This is why the transport of water in this organ seems to be an important process for the effective reproduction. Recently, AQP7, 8, and/or 9 have been shown to participate in water influx across the ovarian follicle wall primarily through transcellular transport mechanisms in the rat (McConnell et al. 2002). Importantly, AQP9 is the member of aquaglyceroporin subgroup, which is permeable not only to water but also to glycerol, urea, and other non-electrolytes. Hence, the expression of this AQP9 protein in granulosa cells may suggest that transport of above-mentioned substances is also important for the follicle development. In fact, AQP9 might ensure sufficient supply of granulosa cells with substrates (androgens) for estrogen production. Collectively, the expression of studied AQPs in the porcine ovary is confined to various compartments that are likely to be connected with their specific functions.
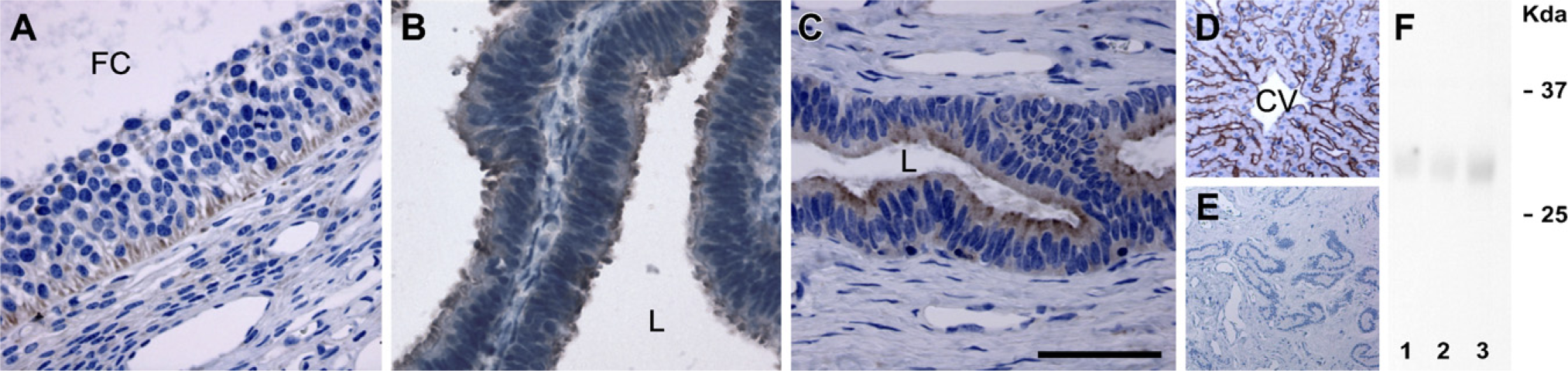
IHC staining of AQP9 in paraffin-embedded sections of the ovary (
AQPs in Oviduct
This study showed expression of AQP1, 5, and 9 in the porcine oviduct: AQP1 in the oviductal capillary, AQP5 in the muscle layer, and both AQP5 and AQP9 in the epithelial cells. In the literature thus far, there are two reports pertaining to AQP localization in oviductal tissues. Branes et al. (2005) showed by IHC the expression of AQP5, AQP8, and AQP9 in the epithelial cells of the rat oviduct. In turn, Gannon et al. (2000) found AQP1 labeling in the innermost and the inner cells of the circular muscular layer of the rat oviduct. These studies suggested that increased water movement into oviductal muscles through AQP1 could lead to muscle swelling that shuts down the lumen. These changes might be implicated in ovum transport toward the uterus. One of the hypotheses (tube locking) concerning the mechanism of ovum transport suggests that the ovum is delayed at the ampullaristhmic junction, probably as a result of muscle contraction, isthmic edema, and vascular distention (reviewed by Huang et al. 2006). In contrast, the pig oviductal muscles exhibited AQP5 expression but not AQP1. However, the expression of AQP1 was present in the pig oviductal vessels. Moreover, in oviductal luminal epithelium, AQP5 and AQP9 were localized. We therefore suggest that these three AQP isoforms (AQP1, 5, and 9) might participate in the epithelial fluid movement in the oviduct and/or the ovum transport toward the uterus.
AQPs in Uterus
In this study, we showed AQP1, 5, and 9 expression in the pig uterus. AQP1 expression was present in the endothelial cells of the blood vessels, AQP5 in the cells of myometrium, and both AQP5 and AQP9 in the cells of uterine epithelia. The first confirmation of AQP1 mRNA expression in the rat uterus was reported by Li et al. (1997). In 2003, two independent groups found AQP1 expression in mouse myometrium (Jablonski et al. 2003; Richard et al. 2003). Recently, Lindsay and Murphy (2006) reported AQP1 expression in endothelial cells of the endometrium and in the inner circular layer of myometrium of rats. Consistent with this, in this study, we showed the expression of AQP1 in the endothelial cells of the uterine blood vessels, whereas no expression was observed in the pig uterine epithelia or myometrium. The presence of AQP5 in the uterine epithelia has been previously shown in ovariectomized rats (Lindsay and Murphy 2006) and pregnant rats (Lindsay and Murphy 2007), as well as in mice during implantation (Richard et al. 2003). Here, we report AQP5 expression in the myometrium and the luminal and glandular epithelium of the pig uterus. Lindsay and Murphy (2007) showed AQP9 expression in the apical plasma membrane of the glandular epithelium of the rat uterus. In this study, AQP9 protein was detected in the glandular epithelium and additionally in the luminal epithelium of the pig uterus.
Immunolocalization of aquaporin 1, 5, and 9 in the female pig reproductive system
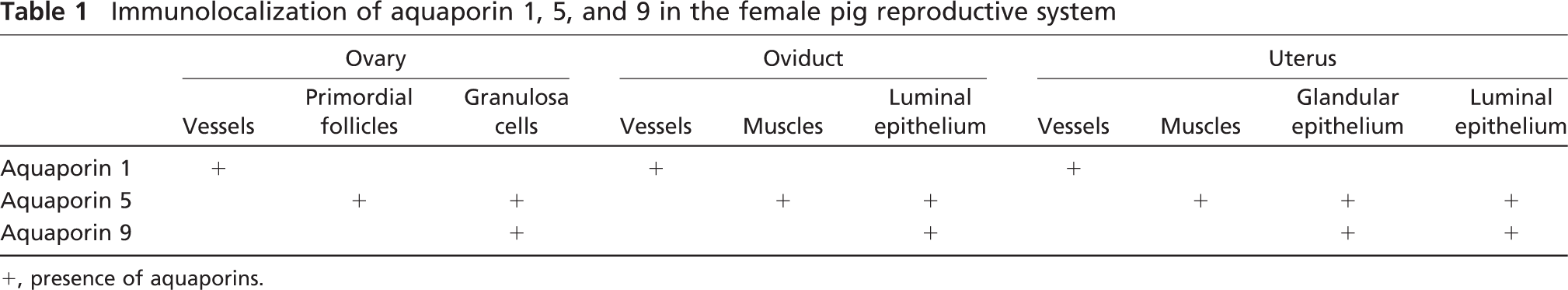
+, presence of aquaporins.
Recently, AQP2 expression was found in human endometrial cells of the uterus (He et al. 2006; Hildenbrand et al. 2006). In contrast, Jablonski et al. (2003) did not observe AQP2 in the ovariectomized mouse uterus, because this protein was strongly upregulated by estrogen in the uterine epithelial cells and myometrium (Jablonski et al. 2003). In this study, we did not localize AQP2 in the pig uterus, whereas anti-AQP2 antibody strongly labeled the apical cell membrane of inner medullary collecting ducts of the pig kidney as a positive control (data not shown). We suggest that specific localization of AQPs (AQP1, AQP5, and AQP9) in the cells of the pig uterus, particularly the presence of aquaporin (AQP5) and aquaglyceroporin (AQP9) in the uterine epithelial cells, might play an important role in the preparation of suitable environment for implantation of the embryo and nourishment of the embryo and fetus during pregnancy.
To our knowledge, this is the first IHC study pertaining to the expression of AQPs in the female reproductive system in pigs. These results suggest the role of AQPs in water homeostasis within tissues of the porcine ovary, oviduct, and uterus. Further studies are necessary to establish the physiological role of these proteins in relation to reproductive processes in the pig.
Footnotes
Acknowledgements
This research was supported by the Polish Ministry of Science and Higher Education (Grants N N308 0042 33 and 0206.0805).
The authors declare that there is no conflict of interest that would prejudice the impartiality of this scientific work. The authors thank Prof. Stanislaw Okrasa for comments and suggestions.