
Abstract
Multiple myeloma (MM) is an incurable B-cell malignancy that arises in the bone marrow (BM). The malignant cells within the BM have extensive interaction with the structural components of their microenvironment. It has been previously shown that the interactions between MM cells and the BM extracellular matrix (ECM) proteins contribute to drug resistance. To understand the underlying causes of adhesion-mediated drug resistance in MM, the components of human BM ECM available for interactions with MM cells must be characterized. We analyzed the expression and localization of fibronectin, laminin, and collagens I and IV in the core biopsies of normal donors and patients with monoclonal gammopathy of undetermined significance (MGUS) or MM. In addition, we compared the patterns of ECM expression in MM patients with low-, mid-, and high-level plasmacytosis of the BM. Although expression of laminin was the same for all groups tested, levels of fibronectin and collagen I were reduced in MM patients with high-level plasmacytosis. Expression of collagen IV in the BM of MGUS and MM patients was higher than in the BM from normal donors. Compared with the plasma cells isolated from the patients with low- and mid-level plasmacytosis, sorted CD138+ plasma cells from MM patients with high-level plasmacytosis overexpressed collagen IV. Our findings show that, compared with normal controls, the ECM composition of the bone, endosteum, and BM is aberrant in patients with MM, further establishing ECM as a key player in the MM disease process.
Keywords
M
As a site of hematopoiesis, BM has a complex organization with multiple cell types occupying distinct niches. A discrete extracellular matrix (ECM) microenvironment within the bone marrow helps to separate endosteum, an interface between bone and BM, from the central marrow. BM ECM, a proteinaceous matrix of mainly fibronectin, laminin, and collagens, is responsible for maintaining the BM architecture by providing a scaffold for the cellular compartments occupying the BM (Kibler et al. 1998; Gu et al. 2003). The ECM composition of the BM is crucial for normal tissue homeostasis, because the chemical treatment or irradiation of the stroma has been shown to lead to tumor formation in otherwise non-malignant epithelium (Barcellos-Hoff and Ravani 2000; Maffini et al. 2004).
The BM microenvironment can influence the therapeutic efficacy by conferring drug resistance (Vincent and Mechti 2005). Human myeloma and leukemia cell lines exhibited a drug-resistant phenotype when grown on fibronectin (Dalton et al. 2004; Hazlehurst et al. 2006) or in direct contact with BM stromal cells (Damiano et al. 1999; Nefedova et al. 2003; Schmidmaier et al. 2004). Interestingly, increased resistance to tumor necrosis-related apoptosis-inducing ligand–induced apoptosis was seen in MM cell lines directly attached to BM stomal cells but not when grown on fibronectin, suggesting that various components of the BM micro-environment confer resistance to different drugs, establishing a multidrug resistance phenotype (Damiano et al. 1999; Yanamandra et al. 2006). Expression of adhesion molecules was higher in patients receiving chemotherapy than in chemo-naïve patients, and their expression increased with additional chemotherapy rounds. In the same study, multidrug-resistant patients expressed higher levels of adhesion molecules than patients who responded to treatment (Schmidmaier et al. 2006). Cells grown in autologous ECM were shown to have an improved capacity to repair radiation-induced DNA lesions and to restore clonogenic capacity compared with cells grown on biologically unrelated ECM (Fuks et al. 1992), suggesting that the ECM composition may be crucial in maintaining the balance between drug-sensitivity and drug resistance that allows cells to repair drug-induced damage.
Hematopoietic progenitors cultured without stromal support proliferated significantly more than progenitors adherent to stroma, and proliferation of colony-forming cells was reduced after specific adhesion to stroma (Hurley et al. 1995), implying that stromal elements may maintain the their quiescence. Because essentially all MM patients ultimately relapse, drug-resistant cancer stem cells must escape current therapies (Pilarski et al. 2000, 2002, 2008; Reiman et al. 2001; Donnenberg and Donnenberg 2005). Similarly to BM-mediated quiescence of hematopoietic progenitors, it seems likely that microenvironment plays a central role in maintaining MM cancer stem cells in a quiescent state, perhaps delaying relapse or alternatively preserving the malignant clone throughout therapy. However, because adherent MM cells are drug resistant, new therapies are needed to combat adhesion-mediated drug-resistance. The combination of chemotherapy with peptides blocking cell adhesion to fibronectin holds promise for eradicating drug-resistant malignant cells in the BM of MM patients (Matsunaga et al. 2008).
Previous studies have detected tenascin, laminin, fibronectin, and collagen types I, III, V, and VI in the BM of MM patients (Kibler et al. 1998); however, no attempt has been made to evaluate the differences in the ECM expression between the normal, premalignant, and malignant conditions. To determine whether or not the BM ECM composition is aberrant in MM, we examined the expression of fibronectin, laminin, and collagens I and IV in BM from normal human donors and patients with MGUS or MM. Because of the skewed cellular composition of the BM in patients with MM (Kuehl and Bergsagel 2002), we hypothesized that the composition of the ECM would reflect these cellular aberrations. We show that fibronectin and collagens I and IV proteins had differential expression between normal, MGUS, and MM BM. In addition, the ECM expression in the BM samples from patients with MM was dependent on the degree of BM plasma-cytosis, with a loss of expression of fibronectin and collagen I and an increase in expression of collagen IV corresponding to the increase in plasmacytosis. These data suggest that, in MM, malignant PCs overexpress collagen IV; this hypothesis was confirmed by real-time quantitative RT-PCR (RT-rqPCR) analysis.
Materials and Methods
Patients and Sample Processing
After approval from the Health Research Board (University of Alberta) and the Alberta Cancer Board, with informed consent in accordance with the Declaration of Helsinki, BM core biopsies and BM needle aspirate samples were obtained from patients undergoing BM biopsies at the Cross Cancer Institute. Sample populations were normal subjects, MGUS patients, and MM patients with low-, mid-, or high-grade plasmacytosis. BM core biopsies were EDTA decalcified and embedded in paraffin per standard protocol and were used for subsequent IHC studies. Mononuclear cells were isolated from BM aspirates by Ficoll-Paque gradient centrifugation per the manufacturer's instructions. CD138+ and CD138- BM cells were sorted on an EPICS ALTRA (Beckman Coulter; Mississauga, ON, Canada) based on the fluorescent staining with anti-CD138-PC5 (Beckman Coulter). Cell pellets were frozen in Trizol (Invitrogen; Burlington, ON, Canada), followed by RNA purification and RT-rqPCR analysis.
IHC
Paraffin blocks from five to six individual core biopsies per sample population group were sectioned using standard processing. Sections (5 μm) were stained for fibronectin, laminin, collagen I, and collagen IV expression using a standard avidin-biotin-peroxidase technique. Negative controls omitting primary antibodies were included to verify specificity of the staining. Mouse anti-fibronectin, laminin, collagen IV, and rabbit anti-collagen I antibodies were purchased from Millipore (Temecula, CA), and sample processing and staining were performed by the Department of Laboratory Medicine at the Cross Cancer Institute. The blinded slides were independently scored by two observers for the intensity of expression of fibronectin, laminin, collagen I, and collagen IV. There were no significant differences between the scores obtained by the individual observers. Imaging was done at x400 magnification on a Zeiss Axiovision microscope (North York, ON, Canada) equipped with a color camera.
RT-rqPCR
RNA Isolation
Total RNA was isolated with TRIzol reagent per the manufacturer's instructions from CD138+ and CD138- sorted BM cells from three to six patients from each MM sample population.
cDNA Synthesis
RNA (1 μg) was mixed with 1 μl of 10 μM dT15 primer to a final volume of 12 μl in diethylpyrocarbonate-H2O. Samples were incubated at 70C to anneal the primers. After the above pre-RT step, 4 μl of 5x buffer, 2 μl of dithiothreitol, 1 μl of 10 mM dNTPs, and 1 μl of Superscript (Invitrogen) were added to each sample tube. RT cycle conditions were as follows: 42C for 60 min, 99C for 3 min, and 4C hold. cDNA was diluted at 1:10 in H2O for immediate PCR analysis or stored at −80C. All incubations and cycling were performed in the GeneAmp 9700 thermo-cycler (Applied Biosystems; Streetsville, ON, Canada).
rqPCR
The rqPCR step was performed using the SYBR green method using the DyNAmo HS SYBR Green qPCR kit (New England Biolabs; Pickering, ON, Canada) (Li et al. 2002) with product specificity confirmed by melting curve analysis (Thulien et al. 2006) per the manufacturer's instructions. In short, 1 μl of the 1:10 dilution of cDNA was mixed with 10 μl of SYBR green I master mix, 0.5 μl of 10 μM forward and reverse primers (Table 1), and 8.5 μl of H2O for a final volume of 20 μl. Fluorescence was acquired at each cycle on a DNA Agent Opticon 2 system (Bio-Rad; Hercules, CA) using the following cycling conditions: 94C for 2-min denaturation, 45 amplification cycles (94C for 30 sec, 60C for 30 sec, 72C for 30 sec), 72C for 10-min final annealing, and 4C hold. The c(t) values, analyzed using Opticon Monitor 3 software (Bio-Rad), corresponding to the number of cycles where the rqPCR curve crosses a threshold line set at the midpoint of the log fluorescence expansion, were normalized to GAPDH expression levels. The relative amounts of fibronectin and collagen IV mRNA expressed by each of the samples were reported as a percentage of GAPDH mRNA expression.
Sequences of primers used for real-time quantitative RT-PCR and PCR
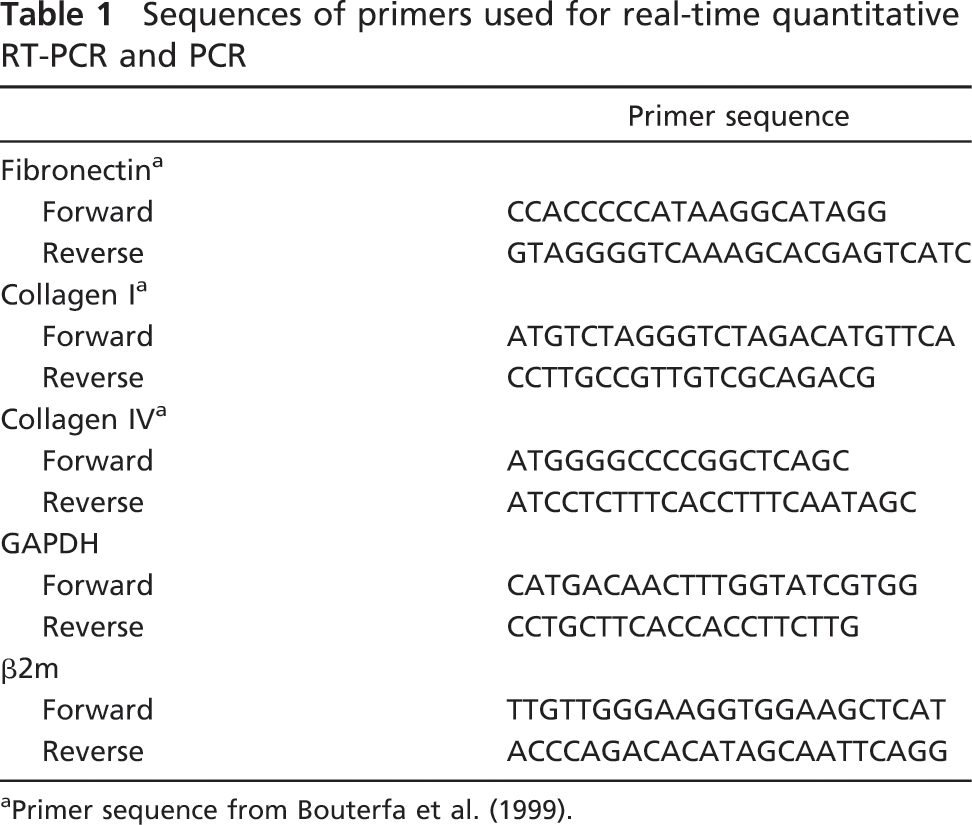
Primer sequence from Bouterfa et al. (1999).
PCR
Total genomic DNA was isolated with TRIzol reagent per the manufacturer's instructions.
Each PCR reaction contained 250 ng of DNA, 19 μl of H2O, 2.5 μl of 10x buffer, 1.0 μl of MgCl2, 0.5 μl of dNTPs, 1.0 μl of 10 μM collagen I, and β2m forward and reverse primers (Table 1) and 0.1 μl of Taq polymerase. PCR conditions were as follows: 94C for 2-min denaturation, 35 amplification cycles (94C for 30 sec, 60C for 30 sec, 72C for 30 sec), 72C for 10-min final annealing, and 4C hold. Amplification products were resolved on a 1% agarose gel stained with ethidium bromide and evaluated by densitometry using an AlphaImager HP (AlphaInnotech; San Leandro, CA) system.
Statistical Analysis
Data are presented as mean ± SEM. Statistical significance was measured by Student's t-test, and Spearman correlation was calculated using Prism 4 software from GraphPad Software. Differences in the mean values are reported as p values, with p<0.05 considered significant.
Results
Expression and Localization of ECM Proteins in BM of Normal Donors, MGUS, and MM Patients
Expression of ECM proteins was analyzed by IHC using the sections of archived BM core biopsies from each sample group of normal, MGUS, and MM with low (<20%), mid (40–50%), and high (>80%) levels of plasmacytosis based on clinical assessment and confirmed by the CD138 IHC staining of the cores. CD138+ cells and aggregates of CD138+ cells were uniformly distributed thorough the entire length of the core. All of the core biopsies analyzed were collected before the initiation of treatment to avoid any changes in ECM proteins that may result from therapeutic interventions. Expression of the ECM proteins was uniform throughout the entire length of the BM core. Table 2 lists the clinical features of the samples used within each group.
Clinical parameters of the BM donors
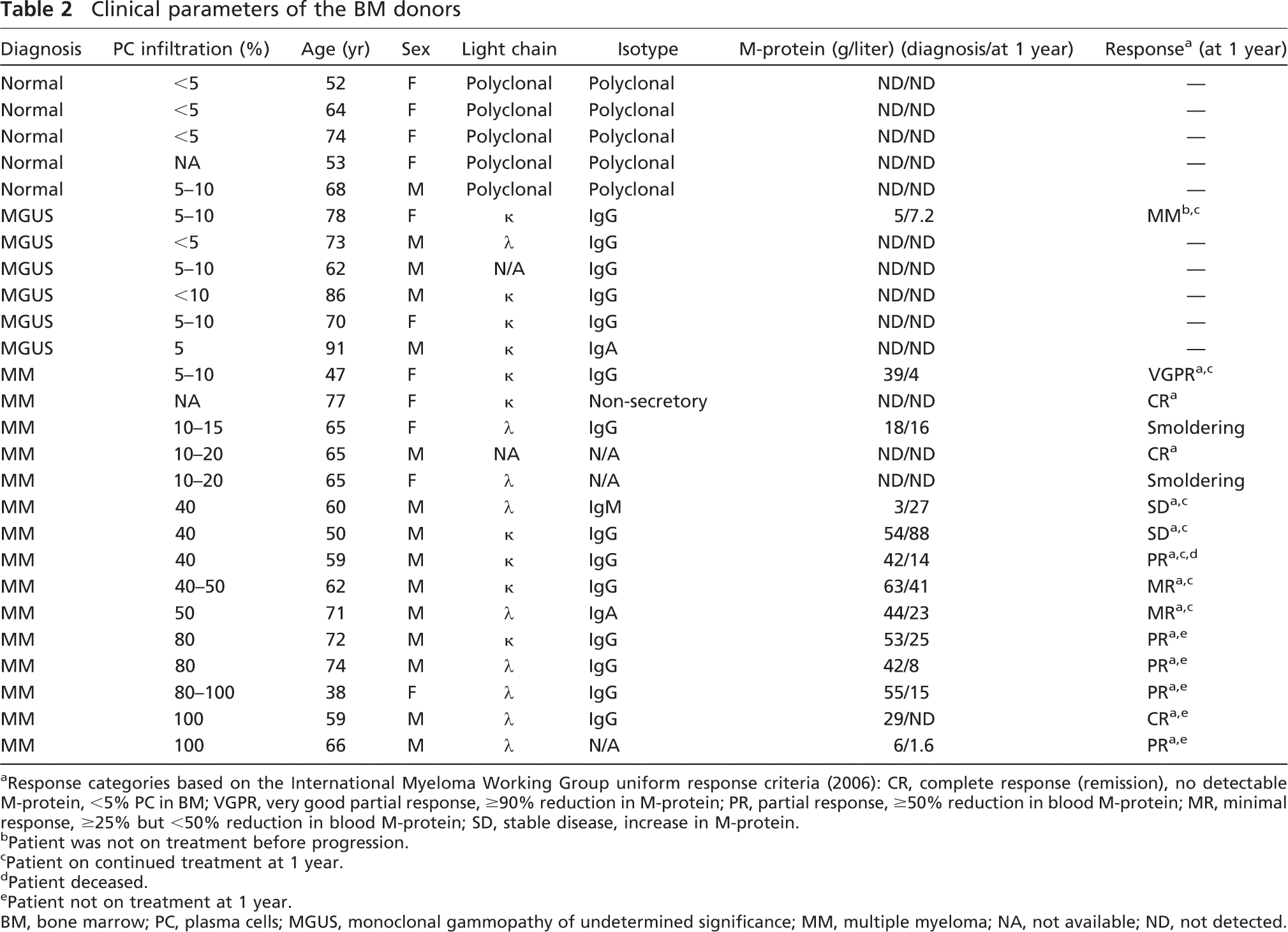
Response categories based on the International Myeloma Working Group uniform response criteria (2006): CR, complete response (remission), no detectable M-protein, <5% PC in BM; VGPR, very good partial response, ≥90% reduction in M-protein; PR, partial response, ≥50% reduction in blood M-protein; MR, minimal response, ≥25% but <50% reduction in blood M-protein; SD, stable disease, increase in M-protein.
Patient was not on treatment before progression.
Patient on continued treatment at 1 year.
Patient deceased.
Patient not on treatment at 1 year.
BM, bone marrow; PC, plasma cells; MGUS, monoclonal gammopathy of undetermined significance; MM, multiple myeloma; NA, not available; ND, not detected.
IHC allowed the analysis of ECM in the bone/ endosteum and in the central marrow of the BM biopsies (Figure 1). The entire length of the core was examined, and ECM expression was homogeneous throughout the core. Unlike mouse bone, which was previously shown to express high levels of both fibronectin and collagen I (Nilsson et al. 1998), human bone was positive only for collagen I (Figure 1). Levels of fibronectin in the central marrow were the same in the normal, MGUS, and MM with low-level plasmacytosis and were decreased in MM samples with mid- or high-level plasmacytosis (Figure 1A). There were no obvious differences in laminin expression between sample populations for the BM or for the bone/endosteum (Figure 1B). Similar to fibronectin expression, collagen I showed a decreasing staining intensity with increased levels of plasmacytosis. The normal, MGUS, and MM with low-level plasmacytosis showed very strong expression of collagen I, MM with mid-level plasmacytosis had strong, and MM with high-level plasmacytosis had moderate intensity of collagen I expression (Figure 1C). Finally, collagen IV had weak expression in the normal BM, with increased intensity of staining in MGUS and MM samples (Figure 1D). For all proteins analyzed, the expression levels of ECM proteins at the endosteum followed the same trend as for the BM but were slightly less intense (Figure 1). Table 3 summarizes the expression levels and spatial localization of the ECM proteins for the human BM, bone, and endosteum.
Cellular Origin of ECM Proteins in the BM
To determine whether the malignant PCs contribute to changes in the expression of ECM proteins in the BM of patients with MM, we analyzed the expression of the fibronectin, collagen I, and collagen IV mRNA in sorted CD138+ and CD138- BM cells by RT-rqPCR. CD138, an antigen expressed on the surface of PCs, was used as a marker to separate the PC population from other compartments of the BM. Laminin was not included in the RT-rqPCR analysis because it was not differentially expressed in the BM of normal donors or MM patients. Because we wanted to determine whether the level of plasmacytosis affects the BM mi-croenvironment, we restricted the evaluation of the mRNA levels of ECM proteins to MM patients. Normalized to GAPDH, there was no difference in the expression of fibronectin in CD138+ and CD138- cells among the MM sample groups (p<0.05; Figure 2A). This suggests that the differential expression of fibronectin seen in IHC between low- and mid-/high-level MM plasmacytosis may occur predominantly in the stroma. In contrast, collagen IV was overexpressed by the CD138+ cells in BM with mid- and high-level MM plasmacytosis (p<0.001) compared with the MM BM with low-level plasmacytosis (Figure 2B), implying that, as malignant PCs increase in number, they also acquire an increased capability to express collagen IV. There was no difference in the expression of collagen IV in the CD138- cells from any of the groups tested (p<0.05), providing an internal control for each patient. Figure 1E shows that a large fraction of CD138+ cells also expresses collagen IV, confirming the RT-qPCR analysis. Expression of collagen I mRNA was undetectable in either CD138+ or CD138- cells for all samples tested. Genomic collagen I was successfully amplified using the same set of primers (Figure 2C), providing a positive control for the assay. Thus, collagen I does not seem to be expressed in the hematopoietic compartment of the BM, suggesting it may be derived from stromal cells.

Expression of extracellular matrix (ECM) proteins in the human bone. IHC analysis of the core biopsy sections from normal donors (n = 5) and patients with monoclonal gammopathy of undetermined significance (MGUS; n = 6) and multiple myeloma (MM; n = 5 for each of the low/mid/high plasma-cytosis groups), showing the staining (brown) of bone marrow (BM), bone, and endosteum for (A) fibronectin, (B) laminin, (C) collagen I, and (D) collagen IV. The entire length of the core biopsy was examined, and images are shown from the representative field of view. (E) High-magnification staining of BM core biopsy serial sections. Bars: A–D = 200 μm; E = 10 μm.
Expression and spatial localization of the ECM proteins in human bone a
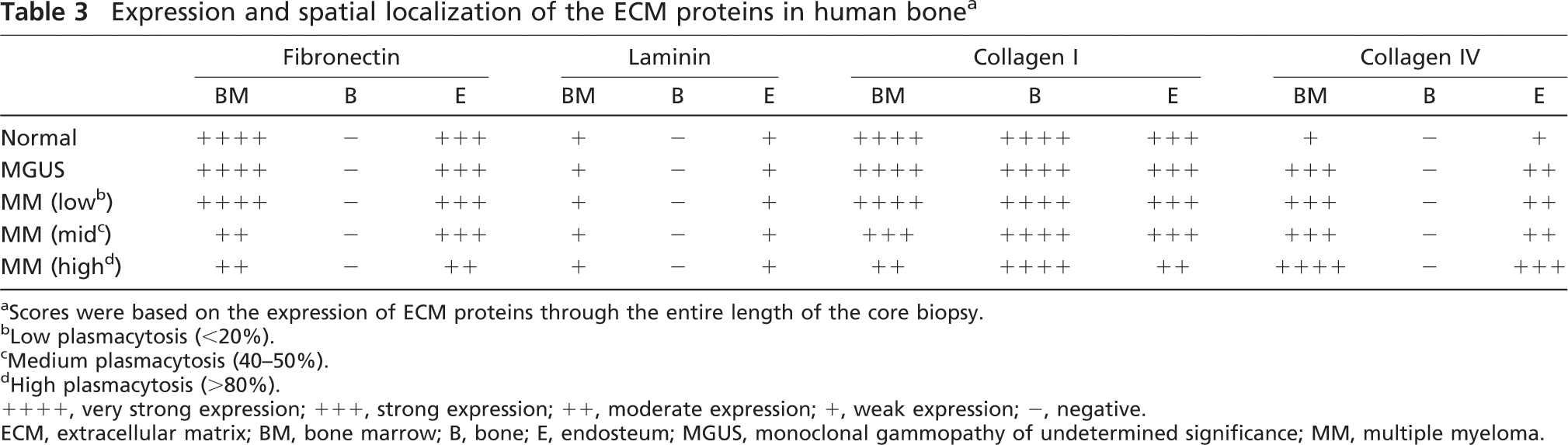
Scores were based on the expression of ECM proteins through the entire length of the core biopsy.
Low plasmacytosis (<20%).
Medium plasmacytosis (40–50%).
High plasmacytosis (>80%).
++++, very strong expression; +++, strong expression; ++, moderate expression; +, weak expression; −, negative.
ECM, extracellular matrix; BM, bone marrow; B, bone; E, endosteum; MGUS, monoclonal gammopathy of undetermined significance; MM, multiple myeloma.
Discussions
MM remains incurable, and patients develop resistance to currently available drugs and drug combinations. To better understand the elements that contribute to adhesion-mediated drug resistance, we evaluated the composition of the microenvironment from normal donors, patients with MGUS, and fully developed MM. The patients with MM were further subdivided into three categories: low, mid, and high degrees of plasma-cytosis. We found that normal human BM expresses high levels of fibronectin and collagen I and low levels of laminin and collagen IV. Our work showed that human BM differs from mouse BM, which expresses fibronectin, laminin, and collagen IV, but not collagen I (Nilsson et al. 1998). The only ECM protein expressed in the human bone was collagen I, whereas mouse bone was positive for both collagen I and fibronectin. The endosteal surface of human bone was composed of fibronectin and collagen I and to a lesser degree collagen IV and laminin. This pattern was very similar in the mouse. The similarities between the ECM composition of the endosteal surface between human and mouse bones suggest that this region may govern biological processes that are conserved between the species.
BM includes distinct niches, with hematopoietic progenitor cells (HPCs) found at the endosteal surface of the bone, more differentiated hematopoietic cells such as B cells and PCs in the central marrow, and the most differentiated cells localized furthest away from the endosteum (Lord et al. 1975; Calvi et al. 2003; Zhang et al. 2003; Kollet et al. 2006; Haylock et al. 2007). The contact between HPCs and osteoblastic cells expressing osteocalcin and N-cadherin is essential for the maintenance of the HPC niche (Calvi et al. 2003; Kollet et al. 2006); this interaction has been shown to maintain dormancy of normal HPCs (Kiel et al. 2005; Adams and Scadden 2006; Sugiyama et al. 2006; Wilson and Trumpp 2006). Therefore, disrupting the endosteal surface will disrupt the niche and may have dramatic affects on stem cell biology. However, disrupting the stem cell niche may be beneficial when trying to eliminate MM. In the ex vivo BM reconstructions, we localized putative MM cancer stem cells to an endosteal niche (Kirshner et al. 2008). Disrupting this niche may allow chemotherapeutic drugs to circumvent adhesion-mediated drug resistance of cancer stem cells. Thus, not only is the ECM crucial for maintaining the stem cell niche, it plays a central role in adhesion-mediated drug resistance and should be considered during the development of new therapeutics.
A variety of reports document that malignant cells attached to ECM are more drug resistant than cells grown without ECM support (Hazlehurst and Dalton 2001; Westhoff et al. 2008). Therefore, it is important to understand the ECM composition of the diseased tissue to evaluate potential mediators of drug resistance. It seems likely that most MM patients progressed through an early stage of monoclonal gammopathy (MGUS) that is clinically defined as non-malignant, although malignant changes have been identified in MGUS (Billadeau et al. 1996; Gutierrez et al. 2003; Martin-Jimenez et al. 2007). We found a dramatic increase in the expression of collagen IV in the BM of patients with MGUS compared with normal donors. Interestingly, we did not observe any differences between MGUS and MM characterized by low plasmacytosis, suggesting that an increase in the BM plasma cell infiltration >20% is needed to induce macroscopic affects on the BM ECM. However, we noted that the ECM composition of the BM changed in MM BM biopsies having a high level of plasmacytosis. The levels of fibronectin were decreased in mid compared with low plasmacytosis biopsies but remained at the moderate level of expression even in MM biopsies with high-level plasmacytosis. A similar pattern was seen for collagen I but with a further decrease in MM with high plasmacytosis. The extent to which these patterns are dependent on the location at which the biopsy is taken or alternatively reflect more systemic distributions remains to be determined, as does the extent to which ECM distribution patterns correlate with response to treatment.
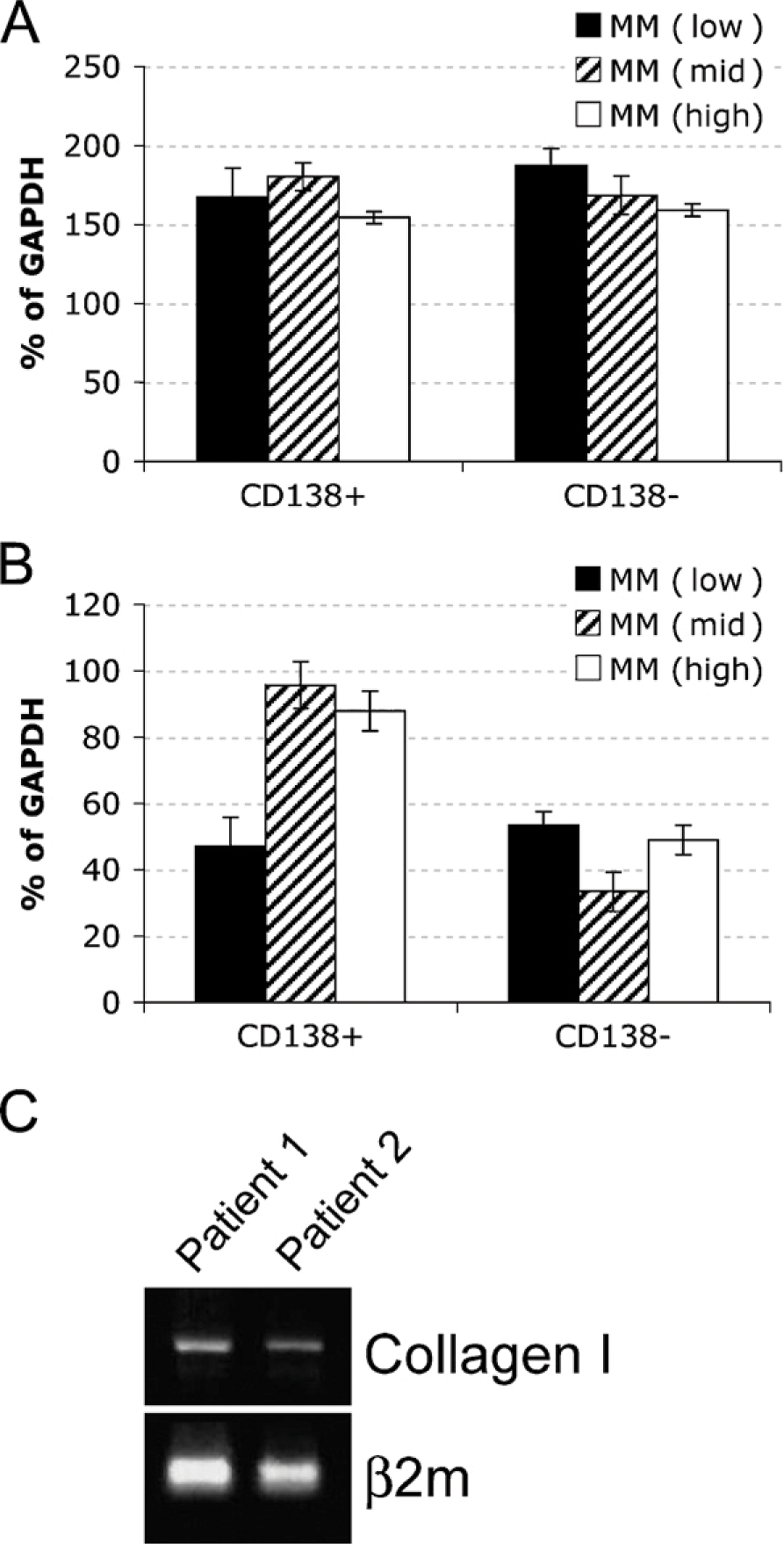
Contribution of CD138+ cells to the expression of fibronectin and collagens I and IV in patients with MM. (
Stomal compartments of the BM are normally responsible for secreting the ECM (Zuckerman and Wicha 1983); however, other cellular compartments may be directly or indirectly involved in this process. Non-stromal compartments may secrete ECM protein or may modulate the composition of the ECM though signaling, resulting in an increase or a decrease in the production of ECM components by the stroma. Here we assessed the contribution of the malignant PCs to the aberrant expression of fibronectin and collagens I and IV in the BM of MM patients. After normalization to GAPDH, collagen IV was overexpressed in CD138+ cells in the MM patients with mid/high plasmacytosis compared with the CD138+ cells from the patients with a low number of PCs in the BM. It is possible that the interaction between PCs in the mid-/high-grade BMs creates a feedback loop with more collagen IV produced by the PCs, which are in contact with other PCs. Like collagen IV, fibronectin was expressed by both CD138+ and CD138- cells, but its levels did not change with the degree of plasmacytosis. Interestingly, in long-term BM cultures, stromal cells, but not hematopoietic cells, secreted fibronectin (Reincke et al. 1982). This emphasizes the need to evaluate the tissue microenvironment in vivo, and not in culture, because cellular phenotypes change once a cell is taken out of its physiological context. Taken together, our results show that the ECM composition of the BM in patients with MM differs from that of patients with MGUS and normal donors.
Footnotes
Acknowledgements
This work was funded by grants from the Canadian Institutes of Health Research and the Alberta Cancer Board Research Initiatives Program. J.K. was funded by a fellowship award from the Alberta Heritage Foundation for Medical Research. L.M.P. is the Canada Research Chair in Biomedical Nanotechnology, and this work was funded in part by the Chairs program.
We thank the members of the Department of Laboratory Medicine at the Cross Cancer Institute for their technical expertise and assistance in IHC.
The authors declare that they have no competing financial interests.