
Abstract
During organ differentiation, cell-extracellular matrix (ECM) interactions are required. The components of the ECM, such as glycosaminoglycans, fibronectin, laminin, and collagens, change in relation to cytokine and enzyme activity. Moreover, glycosaminoglycans (GAGs) are components of the ECM that play an important role in both cytokine regulation and cell activities. In this work we studied the accumulation of hyaluronic acid and chondroitin sulfate and heparan sulfate proteoglycans (PGs), β-N-acetyl-
Keywords
L
For this purpose, we have studied the changes in ECM composition, β-NAG enzyme activity, and TGFβ2 and IL-1 distribution in lung buds during bronchial branching at different stages of development. Moreover, we have analyzed the total GAG and single classes of GAG in the whole lung bud at the same stages.
Materials and Methods
Hubbard fertilized eggs provided by Selice Incubator (Bubano, Imola, Italy) were incubated at 38C and 60% relative humidity. Chick embryo lungs at 5, 7, 9, 11, 14, and 18 developmental days (20 eggs per stage), according to the Hamburger-Hamilton tables (Hamilton 1965), were removed, fixed in 10% formalin, cut in serial sections at 4–7 μm, and stained for histochemical and immunohistochemical data. At the same lung development time, GAG and β-N-acetyl-
Histochemical Technique
Alcian Blue Staining. The histochemical study was performed on homologous sections of intraclavicular air sac, ectobronchi, parabronchi, and entobronchi on the basis of previous indications of the spatial GAG distribution and tridimensional reconstruction of epithelial lung branching in vivo (Becchetti et al. 1988; Stabellini et al. 2002). GAGs were identified by critical electrolyte concentrations at which the polyanions changed from binding Alcian to Mg++ (Scott and Dorling 1965). Alcian stained polyanions with increasing selectivity as the MgCl2 concentration in the staining solution increased: at MgCl2, all GAG as well as nucleic acid and sulfate glycoproteins; at 0.3 M MgCl2, the only macro-molecules stained positively were the GAGs chondroitin sulfate (CS), dermatan sulfate (DS), keratan sulfate (KS), and heparan sulfate (HS). The Alcian blue technique, used to distinguish different GAGs has been described previously (Becchetti et al. 1988). Briefly we used 1% Alcian blue 8GX staining (AB) (Sigma-Aldrich; St Louis, MO) in 0.1 M acetate buffer, pH 5.8, in the presence of 0.025 M or 0.3 M or 0.65 M MgCl2 solution (Sigma-Aldrich) for 2 hr. For enzymatic digestion, the sections were incubated with testicular hyaluronidase (Merck, Darmstadt, Germany; 1 mg/ml in 0.1 M phosphate buffer, pH 7.6, 6 hr at 37C). Control sections were incubated in buffer alone. The action of specific enzymes on the section, followed by Alcian blue staining, allowed us to determine the distribution of individual gly-cosaminoglycans. Digestion with testicular hyaluronidase, in particular, selectively removed HA and CS. GAG values were obtained by connecting a Zeiss Axioplane Microscope to a Kontron Electronic Scanner using Vidas Software with a specific canal that converted the blue color into gray scale (arrangement: black = 0, white = 1). We prepared three slides of whole lung sections for samples. The values are expressed as relative optical density and were the mean ± SD of five determinations per slide. The sections were stained with hematoxylin-eosin for morphological examination.
β-NAG Localization
For histochemical localization the samples were fixed in 3.7% formaldehyde in 0.1 M PBS, pH 7.25. They were frozen in liquid nitrogen and sectioned with cryostat in 7-μm sections. The β-NAG staining was assayed according to the method previously described (Stabellini et al. 2002). Briefly, sections were immersed in 0.1 M Na-citrate buffer, pH 5.2, containing 0.25 mg/ml Fast-Garnet GBC (Sigma-Aldrich; F-8761) and 0.2 mg/ml naphthol AS-BI N-acetyl-β-
Immunohistochemistry
CS and HA Localization. For CS analysis, a monoclonal mouse antibody (Bio Makor, Rehovot, Israel; code 6505) diluted 1:100 was used. The secondary antibody against mouse IgG was conjugated with alkaline phosphatase (Sigma, A-3688). For HA analysis, we used a probe constituted of hyaluronectin, a glycoprotein extracted from brain by Delpech et al. (1991), which is able to bind to HA and not to any other GAG. The hyaluronectin conjugated with alkaline phosphatase was provided by Girard Nicole (Centre Henry Becquerel-Roven) and utilized as described by Marret et al. (1994). For HS analysis, a primary polyclonal rabbit anti-HS antibody diluted 1:250 (kindly supplied by Patricia Simonn-Assmann; Iserm, Strassburg) was used.
Cytokine Localization. For TGFβ2 localization, we used an Rb34 antibody obtained from rabbits immunized against a synthetic peptide consisting of the first 29 amino acids of TGFβ, supplied by Celtrix Laboratories (Palo Alto, CA), diluted 1:40. IL-1 was localized by a rabbit anti-IL-1 antibody (Genzyme; Milan, Italy), diluted 1:40. The secondary antibody against the primary rabbit antibody was a biotinylated goat anti-rabbit IgG (Bio-Division, Milan, Italy, R001–63), diluted 1:250. The third molecule conjugated with peroxidase was streptavidin (Bio-Division, G014–63). For peroxidase visualization, diaminobenzidine was used (DAKO; Carpinteria, CA; S 3000). For alkaline phosphate, Fast Red (Sigma; F-5146) and phosphate naphthol AS-TR (Sigma; N-8518) were used.
Sequential Staining for HA or CS and Glycosidase Reaction. According to previous studies on the spatial distribution of GAGs in chick embryo lung (Becchetti et al. 1988), we also performed a sequential reaction for β-NAG and for HA and CS in 9-day-old lung sections. Control sections for each reaction were also performed without primary antibody.
Biochemical Procedures
GAG Analysis. For biochemical determination of GAG, lung rudiments were removed at 4C from 5-, 7-, 9-, 11-, 14-, and 18-day-old embryos, pooled (30 for each experiment), sonicated for 1 min at 21 Hz (MSE instrument, Model 44), lyophilized, dissolved in distilled water, and further sonicated for 30 sec. This was done for maximum solubility. The GAGs were isolated according to a method previously described (Evangelisti et al. 1984). Individual GAGs were separated by two-dimensional electrophoresis on cellulose acetate plates and identified by comparing them with standard GAG (HA, CS, DS, HS; Sigma) and by their specific ennzymatic susceptibility. Lyophilized samples were digested with bovine hyaluronidase (EC 3.2.1.35) (Miles Italiana; Milan, Italy) and with Streptomyces hyaluronidase lyase (EC 4.2.2.1) (Streptomycetes hyaluroniticus; Seikagaku Kogyo, Tokyo, Japan) at 37C for 24 hr with 30 and 5 enzyme units, respectively, or with chondroitin AC-II lyase (EC 4.2.2.5) (Arthrobacter aurescens; Seikagaku Kogyo) for 24 hr with 0.5 and 0.01 enzyme units (Conrad et al. 1977). The single classes of GAG were quantified using the microcolorimetric methods of Bertold and Page (1985).
β-N-acetyl-
Statistical Analysis
The statistical analysis was performed using the Student's t-test for paired and unpaired data. The differences were considered significant at p = 0.05.
Results
Histochemistry
Alcian Blue. Developing lung rudiments show a progressive increase of second-order tubules emerging from the bronchus, such as ento, ecto, laterobronchi, and then of third-order tubules, the parabronchi (P), always surrounded by mesenchyme. The mesobronchus (Ms) is covered by a pluriseriate epithelium, the second- and third-order bronchi instead by a simple cylindrical or cubic epithelium. After AB reaction at 0.025, 0.3, and 0.65 M MgCl2 concentrations (Figure 1), 5-day-old lung rudiments show alcianophilia around the Ms, which is completely removed by testicular hyaluronidase and chondroitinase ABC, corresponding to the presence of HA and CS, respectively. The mesenchyme areas where the entobronchi emerge show greater alcianophilia at 0.025 and 0.3 M MgCl2, totally removed by testicular hyaluronidase and chondroitinase (HA and CS). In lungs incubated for 14 days (Figure 1) at the same molarity, alcianophilia significantly increases till day 9 of incubation (appearance of laterobronchi), and then decreases (p<0.01) from day 11 (appearance of parabronchi), reaching the lowest point at day 14 of chick embryo lung development (complete bronchial branching). The Alcian blue reaction is lower (p<0.01) around the ectobronchi, laterobronchi, and parabronchi than the entobronchi and very low around the air sac. The alcianophilia at 0.025 and 0.3 M MgCl2 completely disappears after hyaluronidase and chondroitinase digestion (HA and CS).
β-NAG Reactivity. We examined the localization of β-NAG in lungs at developmental days 5, 7, 9, 11, 14, and 18. The enzyme is generally more active in cell compartments than in the extracellular component. The highest degree of β-NAG reactivity was observed in the more recently developed areas, especially on the growth front of epithelial lung branching and in the basal lysosomal compartment within the membrane surrounding the mesenchyme, where the epithelium of second order bronchi grew first, at 5 (Figure 2A) and 7 (Figure 2C) days, followed by that of parabronchi at 9 (Figure 2E) and 11 days (Figure 3A). Histochemical controls of β-NAG reactions were negative (Figures 2B, 2D, 2E, 3A, and 3C).
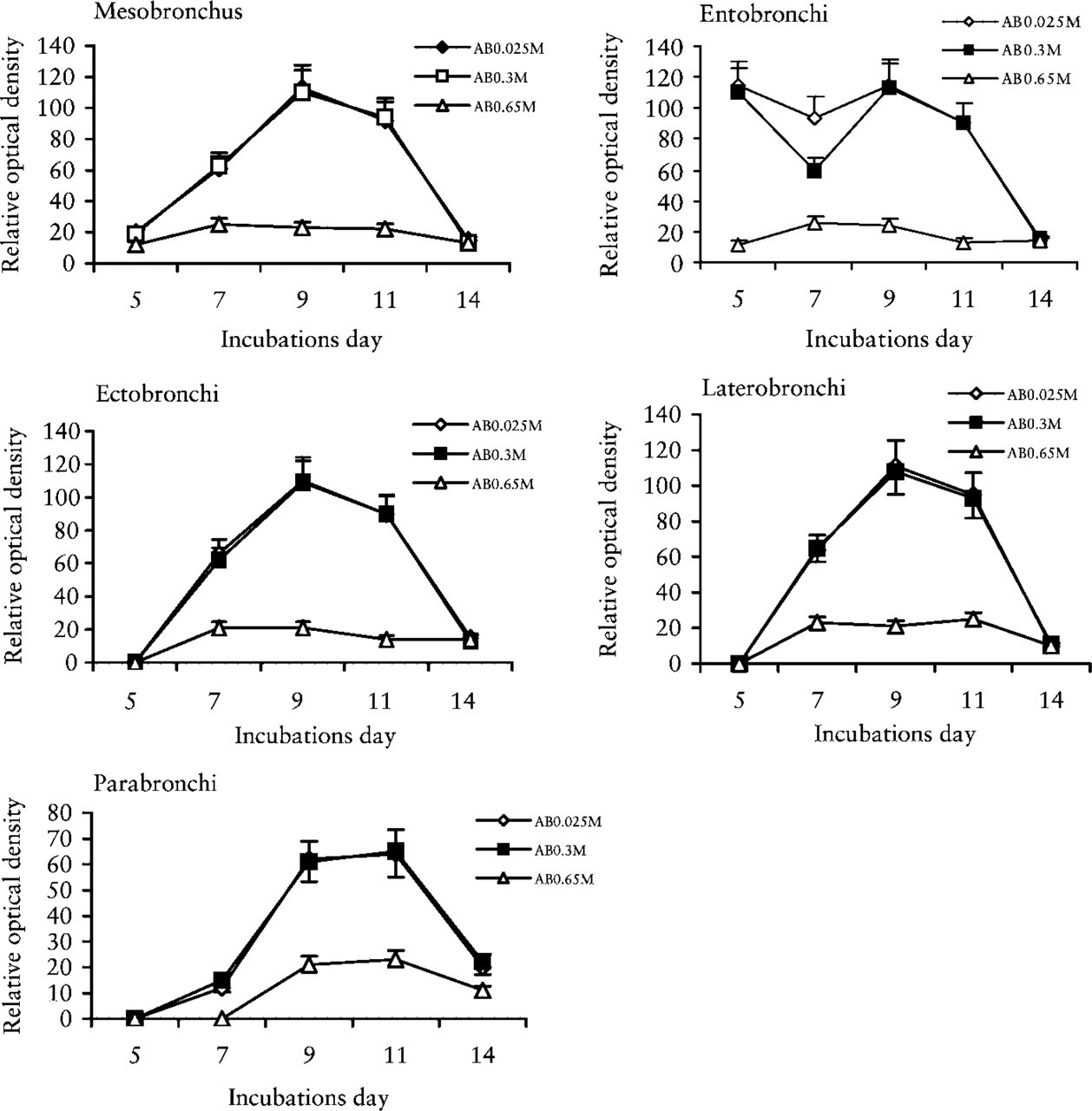
Alcian blue staining at different MgCl2 concentrations. Reactivity of mesenchyme surrounding the mesobronchus, ecto-, latero-, endo-, and parabronchi of bronchial branching in chick embryo lungs at 5, 7, 9, 11, and 14 days of incubation.
Immunohistochemistry
Glycosaminoglycans and β-NAG Localization. The 9-day-old lungs showed a positive reaction for HA and CS PGs in the mesenchyme and basal membrane. The CS PGs were particularly evident along the lateral portion of the bronchial branching (Figure 4A), whereas HA was located either in the mesenchyme or in the most external areas of the expanding lung bud (Figure 4C). HS PG appeared near the basal membrane except in the epithelial growth front (Figure 4E).
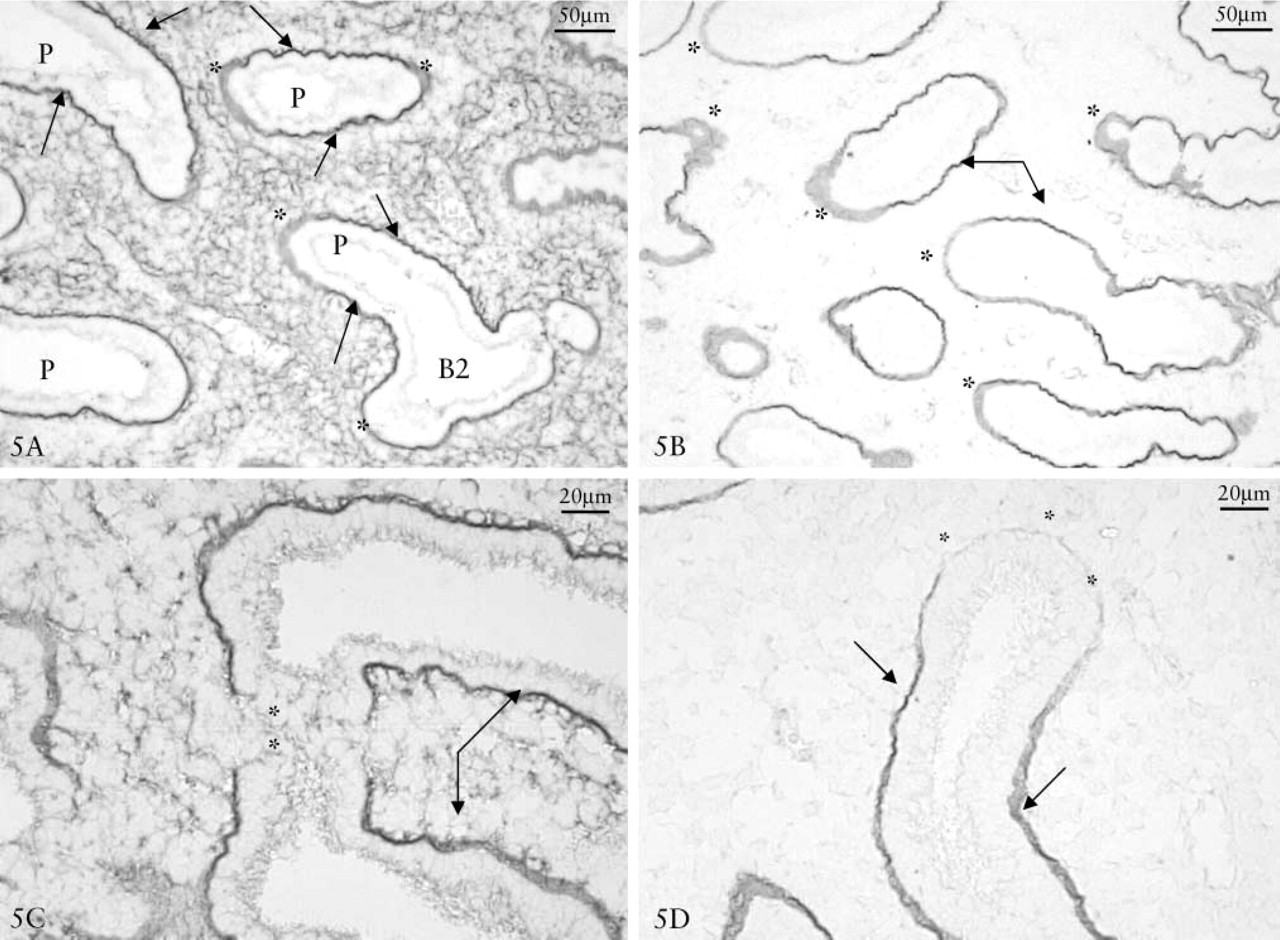
Localization of β-NAG/CS or β-NAG/HA reactions in the mesenchyme around the bronchial branching at 9 days showed that HA and CS were poorly reactive in the area where the enzyme was more concentrated (Figures 4B and 4D). Immunohistochemical controls were negative (Figure 4F). The lungs at 11 days showed that fibronectin (Figure 5A), laminin (Figure 5B), collagen type I (Figure 5C) and type IV (Figures 5D) had a strong positive reaction along the lateral part of the bronchus, whereas they were nearly negative in the growth front.
Cytokine, Chondroitin 4–6 Sulfate and Hyaluronic Acid Immunolocalization. TGFβ2 was located in the basal membrane and particularly in the lateral subepithelial regions of second- and third-order bronchi (Figure 6B). Immunostaining of IL-1 showed the same localization, but with greater reaction in the growth front with respect to the lateral regions of the epithelial tubulus (Figure 6A). The localization of cytokines and GAGs showed that TGFβ2 and CS were expressed in the same regions (particularly in the lateral region of branching) (Figures 4A and 6B), while IL-1 was correlated with HA reaction (particularly in the growth front of branching) (Figures 4C and 6A).
Biochemical Data
GAG Evaluations. Figure 7 shows the trend of non-sulfated (HA) and sulfated GAGs (CS, DS, HS) in lungs at different incubation days. GAGs increased from 5 days and reached their greatest concentration at 11 days, then decreased to their lowest point at 18 days. In addition, single classes (HA, CS, DS, and HS) displayed the same trend, with a peak of greater concentration at the 11 day. The HA/sulfated GAG ratio showed a significant and continuous decrease from 5 to 18 days (Figure 8).
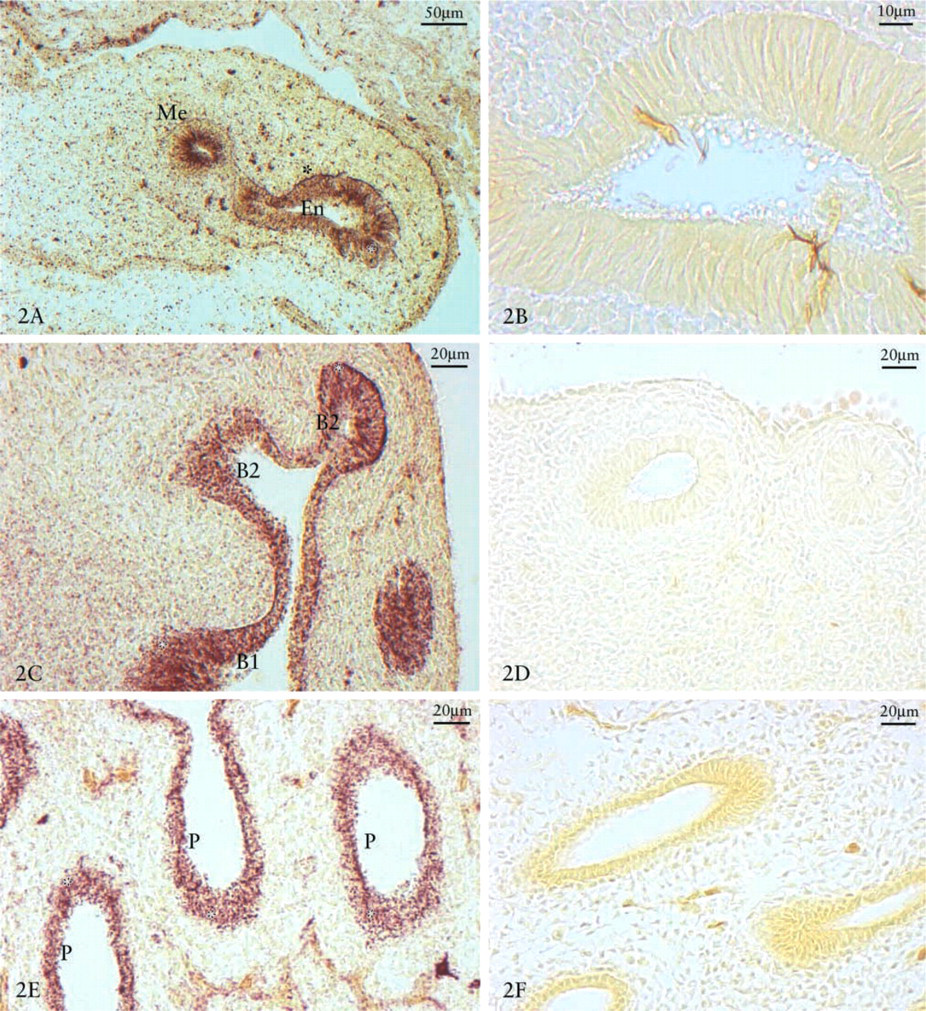
β-NAG reaction in day 5, 7, 9, 11, and 14 chick embryo lungs. Strong positivity in the epithelial growth front (asterisks) where first secondary bronchi, then parabronchi emerge. Five-day-old chick embryo lung (
β-N-acetyl-
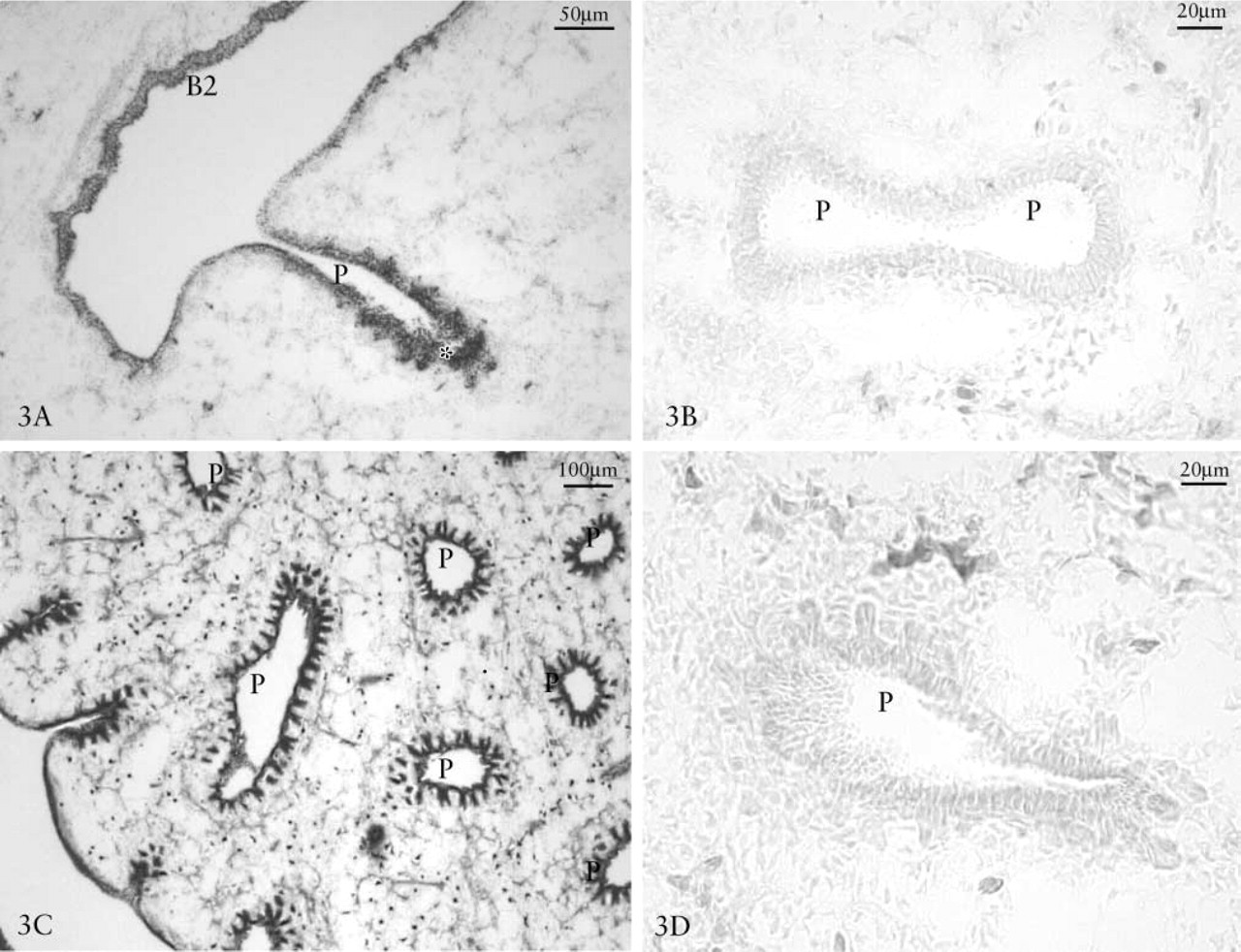
Eleven-day-old chick embryo lung (
Discussion
During chick embryo lung development, bronchial branching shows progressive complexity of ramifications from the day 6 to day 14 of incubation (Hamilton 1965). The gradual development of ento-bronchus, ectobronchi, laterobronchi, parabronchi, and air sacs is related to mesenchymal GAG changes and to cytokine and glycosidase activity. Our data show that during lung branching, when a new bronchial branch emerges, the involved mesenchyme accumulates HA. Instead, when the new branch gradually lengthens, HS PG and CS PG accumulate in the lateral regions. In the same lateral regions of bronchial tubuli, at the epithelial-mesenchymal interface, fibronectin, laminin, and collagen IV are evidenced, whereas they are strongly reduced in the growth front. Collagen I can be detected when the entobronchi have reached complete differentiation, while the underlying mesenchyme shows a decrease in HA and sulfated GAG (day 11 development). During lung development, the decreased HA/sulfated GAG ratio shows that ECM remodeling processes play an important morphogenetic role (Becchetti et al. 1988; Caniggia et al. 1992; Stabellini et al. 2002). At days 6, 9,11, and 14 of lung development, the accumulations of HA in the growth front and of CS in the lateral regions of epithelial branching are in agreement with β-NAG reactivity. The different distribution of degradative enzyme remodels the GAG of the ECM. In fact, a high β-NAG concentration correlates with a decrease in HA and CS. In the epithelial-mesenchymal interface of tubuli, HA co-localizes with IL-1 and CS with TGFβ2. Therefore, during bronchial branching development, GAGs are able to continuously modulate cytokine activity. In the lung the TGFβ isoforms may stimulate production of various TGFβ isoforms (Jakowlew et al. 1994; When et al. 2003). IL-1 shows greater concentration in the growth front of bronchial branching, co-localizing with HA and laminin, whereas TGFβ2 is localized mainly in the lateral regions of bronchial branching, together with CS PG and HS PG, fibronectin, laminin, and collagen IV. Because IL-1 increases synthesis of HA (Bodo et al. 1994), TGFβ1 that of CS (Locci et al. 1993), and HA blocks the effects of TGFβ1 (Locci et al. 1995), these linkages could prevent the inhibiting action of growth factor on epithelial cell proliferation.
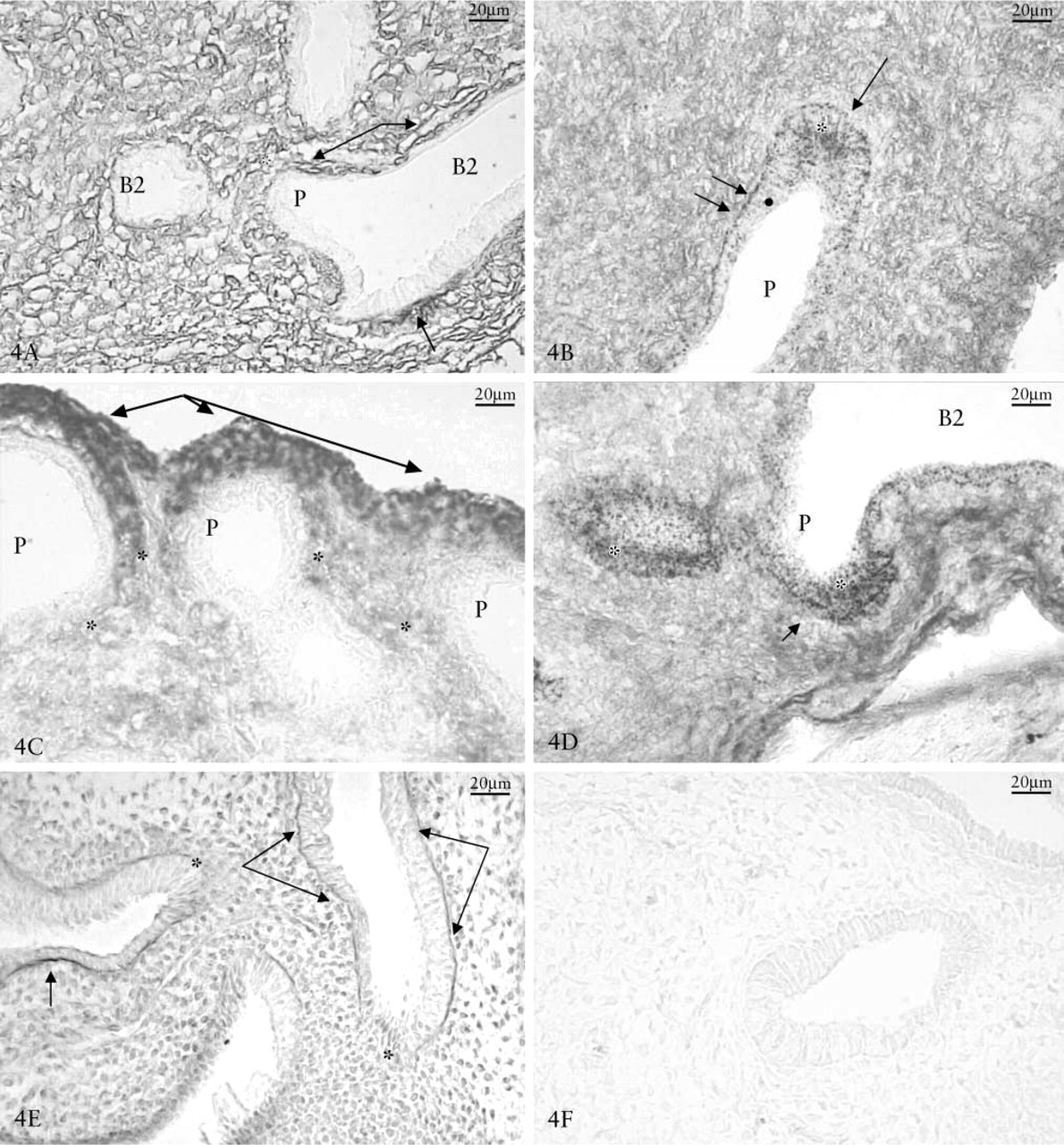
Day 11 chick embryo lungs. (
(
During development of bronchial branching (6–14 days) in the lung, mesenchyme, GAG classes, fibronectin, laminin, and collagen I and IV accumulate along the lateral region of the growing tubules. The stabilization of branching, HS proteoglycan, fibronectin, laminin, and collagen IV therefore play an important role because they intervene in the processes of cell-matrix adhesion and in the regulation of growth factor activities (Miao et al. 1997). Fibronectin is able to link both HS and collagen networks during basal membrane formation. It is present in the lateral regions, where the membranes stabilize the structures, whereas it is absent in the growth front, where HA supports cell proliferation and migration phenomena. The absence of fibronectin, laminin, and collagen IV prevents the formation of basal membrane. It is therefore impossible for them to bind to the cadherin of the cell surface and to form cell-matrix adhesions according to Sakai et al. (2003) in developing structures. Recent studies show that some components of the basal membrane, such as laminin and collagen IV, are able to modulate gene expression of in vitro hepatocytes (Srebrow et al. 2002). It is therefore feasible to outline a sequence of events in which β-NAG activity plays a role in ECM remodeling alongside other enzymes such as N-acetyl-β-
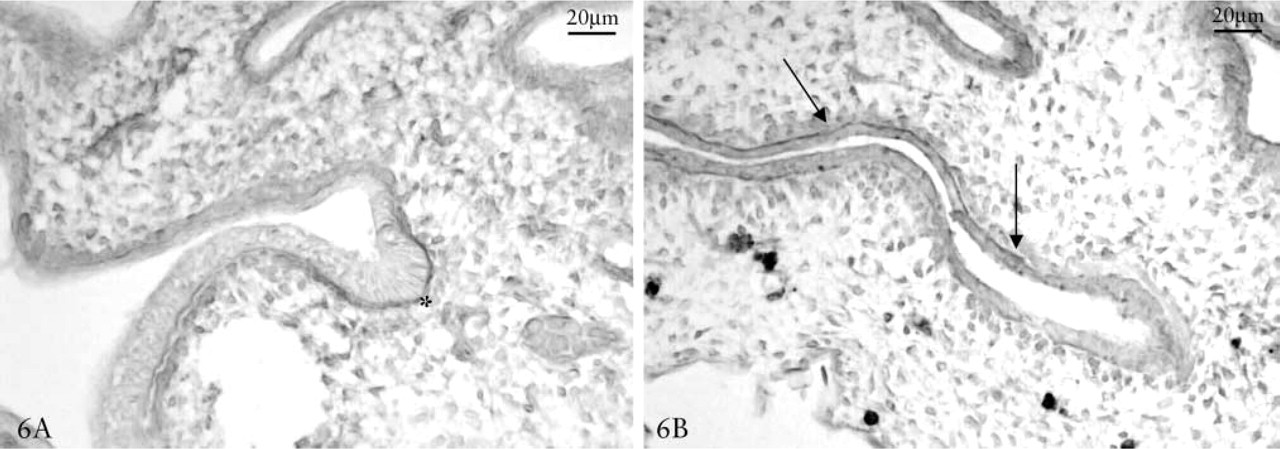
Day 11 chick embryo lungs. IL-1 and TGFβ1 immunostaining. IL-1 is mainly located in underlying basal membrane in the growth front (
In conclusion, GAG distribution changes during lung development and permits or prevents the stimulation of epithelial cells through IL-1 and TGFβ2 in basal membrane formation or in cell proliferation.
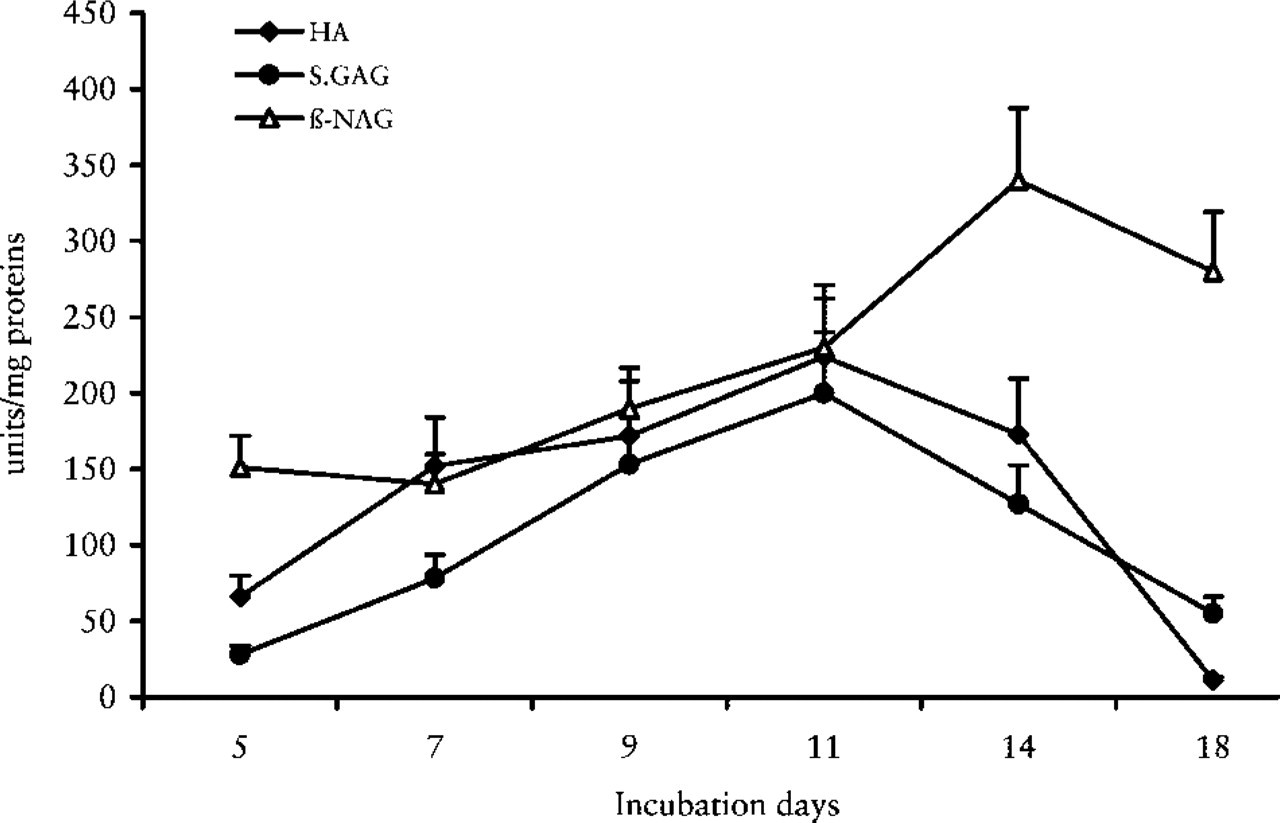
β-NAG, HA, and sulfated GAGs (CS, DS, HS) trends during chick embryo lung development.
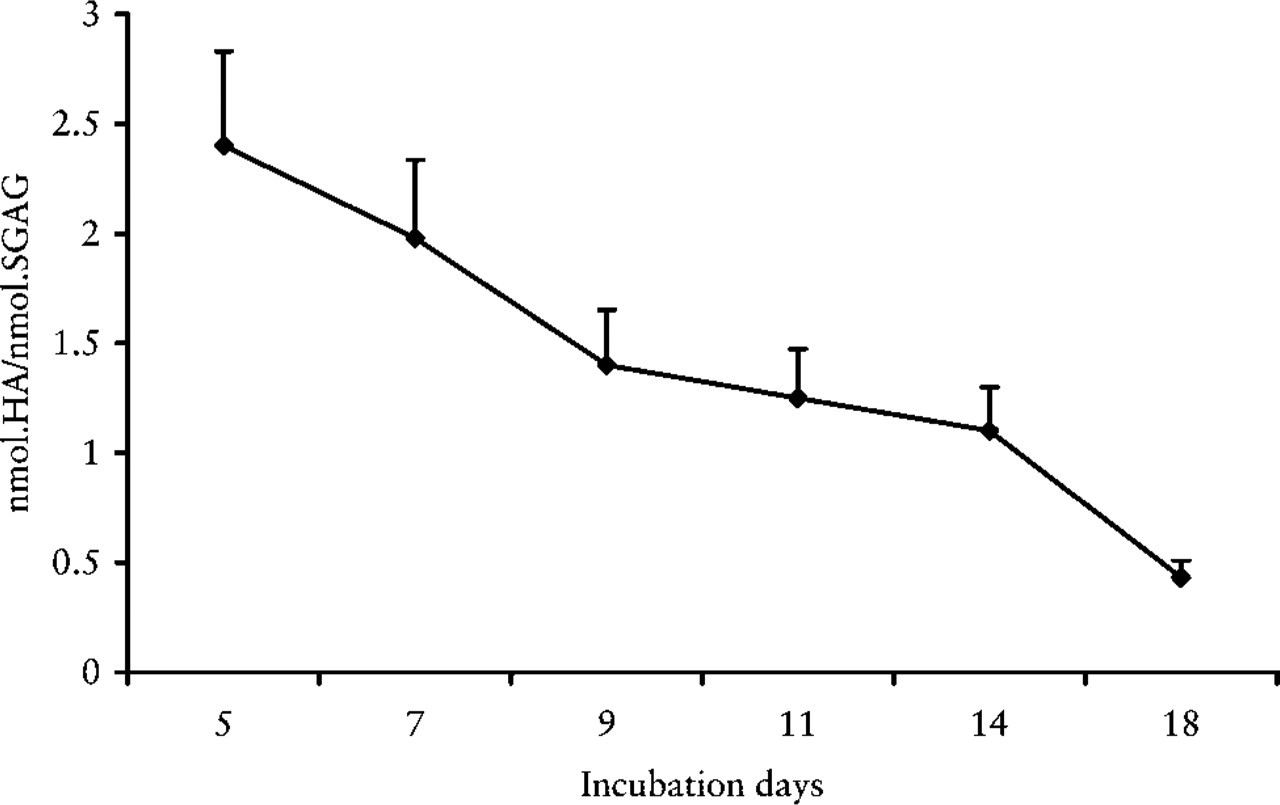
Trend of HA/sulfated GAG ratio in chick embryo lung development.
Footnotes
Acknowledgements
Supported by grants from the Ministero dell'Università e della Ricerca Scientifica e Tecnologica (MURST).