
Abstract
Subsequent to wounding, keratinocytes must quickly restore barrier function. In vitro wound models have served to elucidate mechanisms of epithelial closure and key roles for integrins α6β4 and α3β1. To extrapolate in vitro data to in vivo human tissues, we used ultrathin cryomicrotomy to simultaneously observe tissue ultrastructure and immuno-gold localization in unwounded skin and acute human cutaneous wounds. Localization of the β4 integrin subunit in unwounded skin shows dominant hemidesmosomal association and minor basal keratinocyte lateral filopodic cell–cell expression. After wounding, β4 dominantly localized to cytokeratin-rich regions (trailing edge hemidesmosomes) and minor association with lamellipodia (leading edge). β4 colocalizes with α3 within filopodia juxtaposed to wound matrix, and increased concentrations of β4 were found in cytoplasmic vesicles within basal keratinocytes of the migrating tongue. α3 integrin subunit dominantly localized to filopodia within basal keratinocyte lateral cell–cell interfaces in unwounded skin and both cell–cell and cell–matrix filopodic interactions in wounded skin. This study indicates that β4 interacts with the extracellular environment through both stable and transient interactions and may be managed through a different endosomal trafficking pathway than α3. α3 integrin, despite its ability to respond to alternate ligands after wounding, does so through a single structure, the filopodia.
E
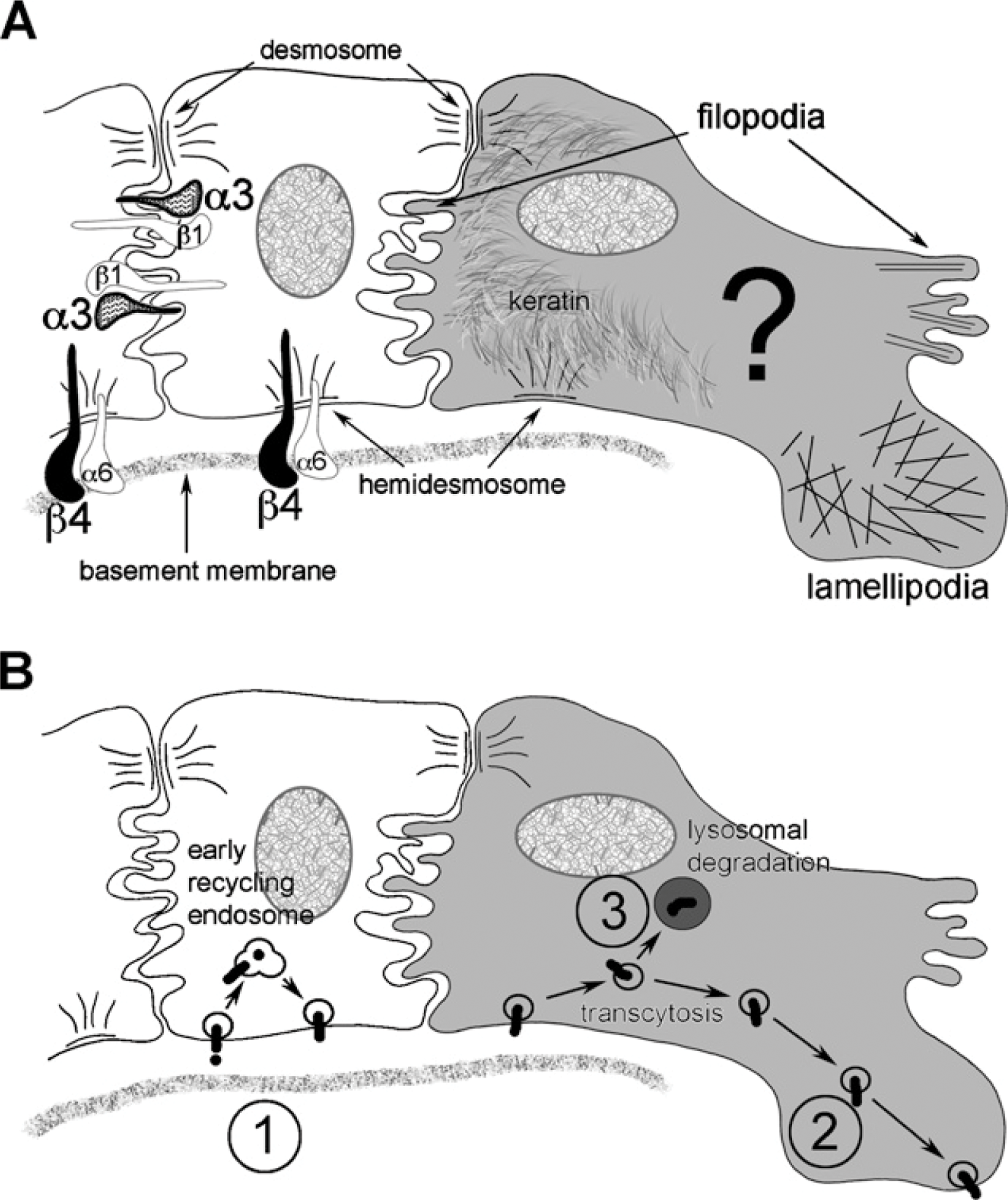
Illustration of keratinocyte features. (
Integrins are heterodimeric transmembrane receptors that play a key role in the maintenance of the quiescent epidermis and epithelialization during wound healing (Grinnell 1992; De Luca et al. 1994; Larjava et al. 1996; O'Toole 2001; Alahari et al. 2002; Martin et al. 2002; Watt 2002). For the purpose of this study, we will divide integrin functionality into two categories: (a) adhesion/signaling (cell–cell or cell–matrix) (Burgeson and Christiano 1997; Singer and Clark 1999; O'Toole 2001; Miranti and Brugge 2002) and (b) intracellular trafficking to manage these adhesions and provide the cell with information about the extracellular environment (Caswell and Norman 2006; Pellinen and Ivaska 2006).
In the quiescent epidermis, integrin α6β4 is associated with the hemidesmosome complex, forming a link in the stable anchorage between cytokeratins and the underlying dermal matrix (Carter et al. 1990b; Borradori and Sonnenberg 1999). The hemidesmosomal complex contains many integral proteins required for functional anchorage. The cytoplasmic tails of α6β4 link with the bullous pemphigiod antigens BPAG1 (BP230) and BPAG2 (BP180) and the intermediate filaments plectin and keratin. The extracellular domains of α6β4 link with the extracellular matrix basement membrane containing their dominant ligand laminin 5 (LM-332). The combination of the β4 subunit and bullous pemphigiod antigen BPAG1 (BP230) can be used, through colocalization, to identify hemidesmosomes within the wound environment (Gipson et al. 1993; Ishiko et al. 1998). Integrin α3β1, on the other hand, associates with actin and forms lateral basal keratinocyte cell–cell focal adhesions (Carter et al. 1990a, b). The story, however, increases in complexity on wounding, where the coordinated change from stable anchorage to migration and wound closure requires dynamic responses from keratinocytes (Laplante et al. 2001; Mercurio et al. 2001; O'Toole 2001; Hackam and Ford 2002; Santoro and Gaudino 2005; Usui et al. 2005). The basic morphological features of the migrating keratinocyte shows polarization in which cytokeratins concentrate toward the trailing edge (Paladini et al. 1996), and thin actin associated cellular projections, filopodia (Gupton and Gertler 2007), and broad cytoplasmic projections, lamellipodia (Choma et al. 2004), form toward the leading edge. (Alberts et al. 2002) (Figure 1A).

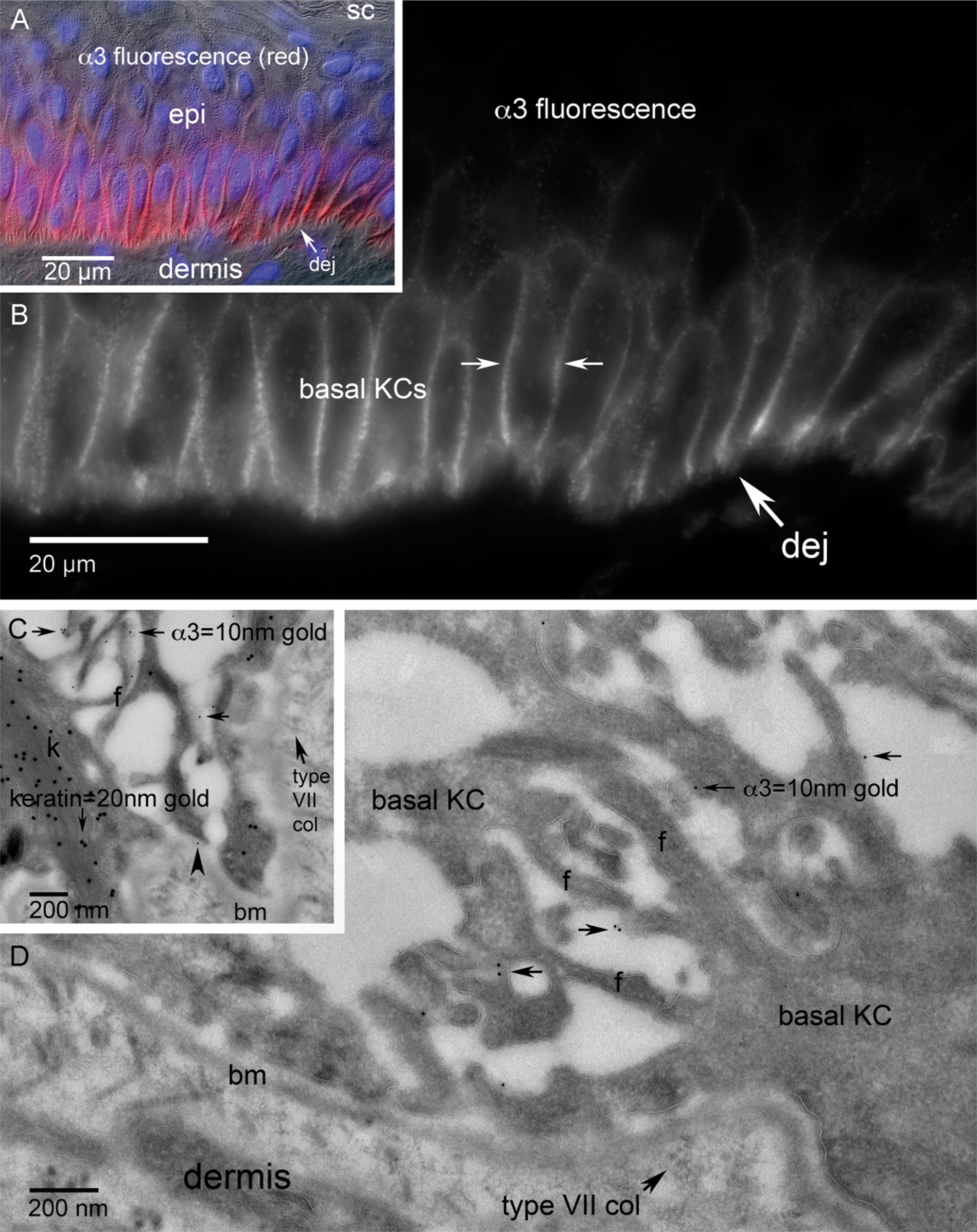
Integrin subunit β4 localization in normal adult human skin. (
In vitro data suggest that, subsequent to wounding, protein kinase C α (PKCα)–mediated phosphorylation of serine residues within the cytoplasmic tail of β4 is integral to α6β4-associated hemidesmosome disassembly (trailing edge) (Gipson et al. 1993; Mercurio et al. 2001; Hackam and Ford 2002; Rabinovitz et al. 2004), decreasing the cytoskeletal/laminin 5 (LM-332)–dependent stable attachments in favor of an actin-rich lamellipodic/filopodic migratory motif at the leading edge (Nguyen et al. 2000b; Santoro and Gaudino 2005). It is believed that, in response to wounding, α3β1 shifts from the Velcro effect of numerous lateral cell–cell focal contacts between the quiescent basal keratinocytes to a preferential ligation with newly deposited precursor laminin 5 (LM-3A32) at the interface between the leading edge keratinocytes and the wound matrix (Nguyen et al. 2000a). It is clear from in vitro studies that this ligation is central to the regulation of cell migration and polarity (Nguyen et al. 2001; Frank and Carter 2004), as well as the stable formation of a α3β1/Rac 1–mediated lamellipodium at the leading edge (Kreidberg 2000; Choma et al. 2004). Light level in vitro data have shown α3β1 to be present on the leading edge of migratory keratinocytes and considered part of the lamellipodium (Choma et al. 2004). Other data have shown that α3β1's interaction with newly deposited LM-3A32 over a collagen matrix occurs toward the trailing edge of the migrating cell (Frank and Carter 2004). What is not so clear is how the morphological location within a stratified in vivo wound relates to α6β4 or α3β1 within the keratinocytes of a monolayer scratch wound model.
The changes in integrin function and/or location are facilitated through intracellular vesicular trafficking, and each integrin subunit has the potential to involve single or multiple pathways in response to a rapidly changing microenvironment (Pelham 1999; Mellman and Warren 2000; Miranti and Brugge 2002). In vitro data would suggest that activated keratinocytes at the leading edge of a wound exhibit increased integrin trafficking to exert minute by minute control of migration polarity, adhesion, and signaling (Caswell and Norman 2006). A brief summary of the possible fates of integrins shows that, subsequent to pinocytosis through clathrin-coated pits (Schekman and Orci 1996; Schmid 1997) or caveolin-mediated non-coated caveolea (Anderson 1998), most will release their ligand in the early/recycling endosome and return to the plasma membrane ready to bind fresh ligand (Mostov et al. 2000), some will transcytose through endosomal transport to a new location in the plasma membrane (Mellman 1996; Hunziker and Peters 1998; Zacchi et al. 1998), and some will progress down a pathway through multivesicular bodies and late endosomes to eventual lysosomal degradation (Beron et al. 1995) (Figure 1B). Our light level immunolocalization of in vivo human wounds showing an increased cytoplasmic signal for β4 in the migrating tongue with no increase in cytoplasmic localization for α3 would support the idea of increased intracellular trafficking for β4 but not α3 and mechanistic differences between the two integrins.
In this study, we test the hypothesis, based on previous in vitro studies and light level immunolabeling of in vivo tissues, that β4 shifts from a dominant cytokeratin/hemidesmosomal association to dominant actin-rich lamellipodic/filopodic localization after wounding, whereas α3 remains associated with focal contacts. We also study whether the increased cytoplasmic localization of β4 seen at the light level localizes to structures indicative of vesicular trafficking.
To date, no ultrastructural evidence has been shown to elucidate the cellular management of β4 and α3 in response to wounding in human skin in vivo.
Materials and Methods
Wounds were created on the upper inner arm of normal volunteers (University of Washington Institutional Review Board approval) using a Surgicutt bleeding time device (Cardinal Health; McGraw Park, IL) (incisional) or a 1.5-mm biopsy punch (excisional). Incisional wounds were removed using a 6-mm biopsy punch at 24–30 hr, and excisional wounds were removed at 48 hr using a 3-mm biopsy punch. A 3-mm biopsy punch of unwounded skin from an adjacent region was used for comparison (Olerud et al. 1995).
For light level examination, excised wound sites were frozen in OCT (Sakura Fine Tek; Torrence, CA), cryosectioned at a thickness of 6 μm, dried onto glass slides, fixed in acetone at −20C, rinsed in Tris-buffered saline, and blocked for nonspecific binding with 2% normal goat serum in Tris-buffered saline. Indirect immunofluorescence was performed at room temperature by incubating with primary antibodies: mouse anti-α3 integrin (P1F2) (Dr. Carter) or mouse anti-β4 integrin (3D5) (Dr. Carter) at dilutions of 1:5, singly or pairwise with rabbit anti-pankeratin (Dako Corporation; Carpinteria, CA) at a dilution of 1:1000. Sections were subsequently labeled with fluorescently tagged secondary antibodies: goat anti-mouse Texas Red (1:200; Jackson ImmunoResearch, West Grove, PA) and goat anti-rabbit Alexa Fluor 488 (1:800; Invitrogen/Molecular Probes, Eugene, OR). Tissue sections were counterstained with 4′,6-diamidino-2-phenylindole (DAPI), allowing visualization of nuclei by the emission of blue fluorescence.
Integrin subunit α3 localization in normal human adult skin. (
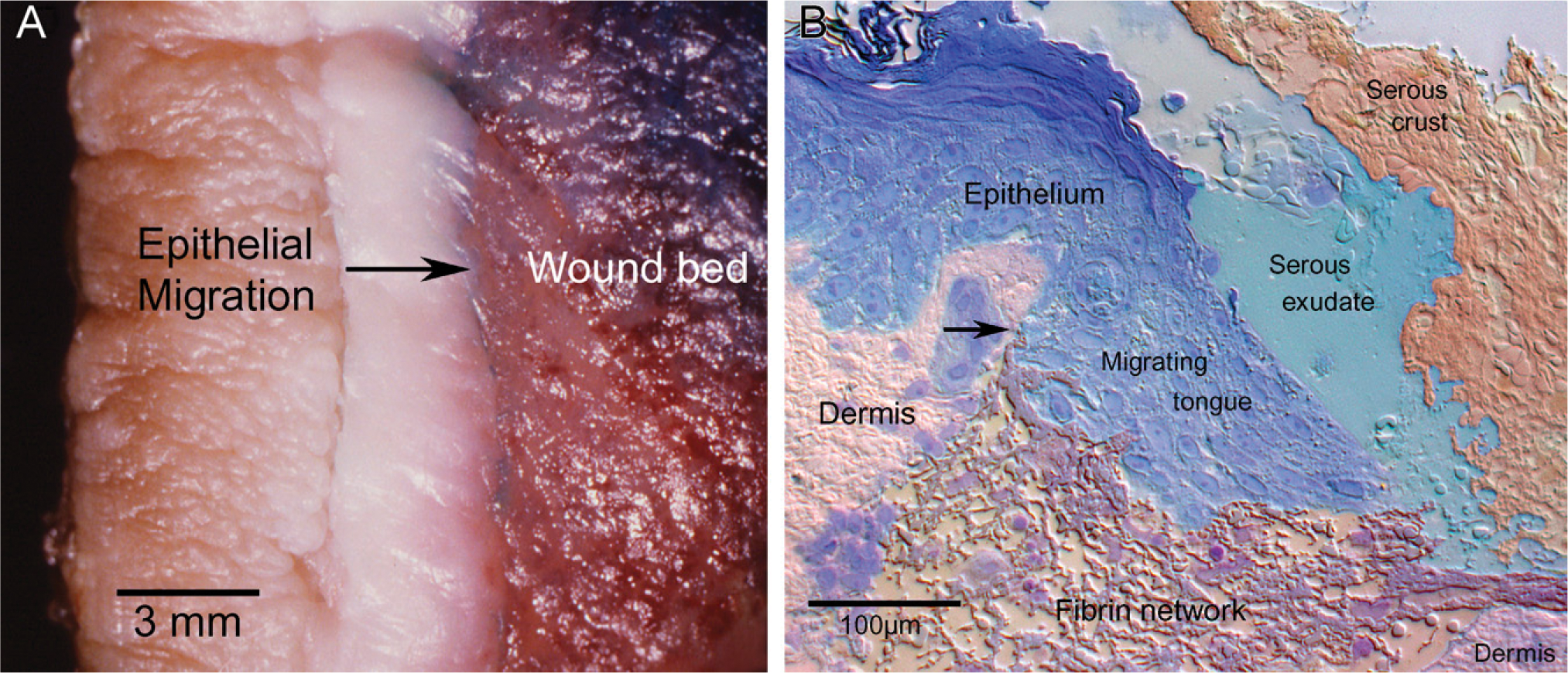
Topography of human cutaneous wounds. (
Specimens were visualized using a Nikon Microphot-SA upright microscope equipped with both transmitted and epi-fluorescence illumination (Nikon Instruments; Melville, NY). Semithin sections (350–500 nm) of specimens prepared for ultrathin cryomicrotomy were Richardson's stained and imaged using brightfield illumination. Differential interference contrast was used to provide tissue topography for both semithin and immunolabeled 6-μm cryosections as needed. Images were captured on a monochrome Photometrics Sensys digital camera with a grade 1 KAF 1400 CCD (Roper Scientific/Photometrics; Tucson, AZ). Accuracy of lateral spatial registration of sequential fluorescence images using multiple probes was maintained using a Ludl filter wheel (Ludl Electronics; Hawthorne, NY) to change excitation wavelengths at the illumination source, whereas the emission was imaged through a single stationary filter cube (Chroma Technology; Brattleboro, VT). Image acquisition and filter selection was controlled through IPLab Spectrum software (Scanalytics; Fairfax, VA) running on a Power Mac G4 dual processor computer.
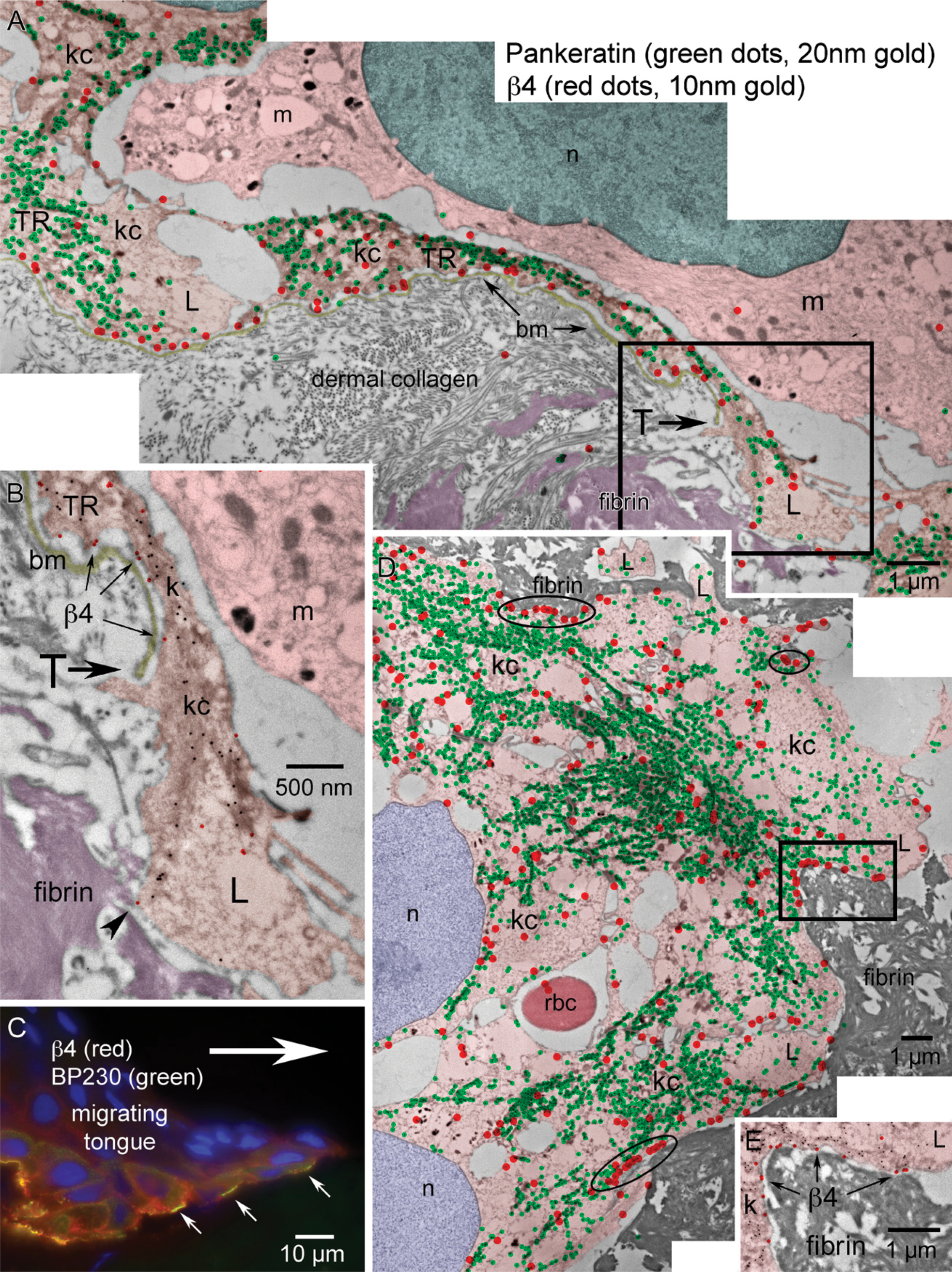
Global view of the distribution of β4 within the epithelial migrating tongue of an incisional wound after 30 hr of healing. (
Ultrathin cryomicrotomy and immunogold labeling were carried out according to the technique described by Tokuyasu (1973, 1980, 1989). Tissues were lightly fixed in 4% paraformaldehyde or a mixture of 4% paraformaldehyde combined with 0.1% glutaraldehyde in Sorenson's buffer for 2 hr on ice and further dissected into 0.5- to 1-mm tissue blocks, allowed to fix for 18 hr at 4C, and rinsed in PBS three times for 10 min each. Quenching of residual aldehydes was achieved by incubating tissue in PBS containing 0.05 M glycine for 30 min. Tissues were rinsed in PBS and infused with 20% polyvinyl-pyrrolidone (PVP)/1.7 M sucrose from 3 hr to overnight for cryoprotection. Infused tissues were mounted on cryospecimen pins in the appropriate orientation for cross-sectioning and quick-frozen in liquid nitrogen. Pins containing the frozen samples were taken from liquid nitrogen storage and mounted in a Leica Ultracut UCT ultramicrotome equipped with the Leica EM FCS cryochamber (Leica Microsystems; Bannockburn, IL). The block face was trimmed at a temperature of −85C to <0.3 mm in width. Semithin sections of 500 nm thickness were collected on droplets of phosphate-buffered 2.3 M sucrose, transferred to glass slides, and histochemically stained (Richardson's) for evaluation of the region of interest at the light level. Ultrathin cryosections of 70–90 nm thickness were subsequently cut using a 35° cryodiamond knife (Diatome; Fort Washington, PA) at −110C, collected on droplets of phosphate-buffered 2.3 M sucrose, and transferred to 200-mesh nickel grids coated with a thin support film of formvar and carbon. Grids were stored face down on droplets of PBS containing 0.05 M glycine and 1% BSA at 4C for subsequent immunolabeling.
Diluent for immunogold incubations and intermediate rinses was PBS containing 1% BSA. The entire procedure was carried out at room temperature. Grids containing the cryothin sections were incubated on droplets of diluent for 30 min and transferred to droplets of protein-specific primary antibodies: mouse anti-integrin subunit α3 (P1F2) or mouse anti-integrin subunit β4 (3D5) targeting the cytoplasmic domain (Dr. William Carter), rabbit anti-BP230 (Dr. John R. Stanley, University of Pennsylvania), and rabbit anti-pankeratin (Dako Corporation) in diluent, at dilutions of 1:1, 1:1, 1:1000, and 1:1000, respectively. Incubations were allowed to continue overnight (18 hr). Grids were rinsed three times for 20 min each and transferred to droplets of species-specific secondary antibodies: goat anti-mouse conjugated to 10-nm gold and goat anti-rabbit conjugated to 20-nm gold (Ted Pella; Redding, CA) diluted 1:40 in diluent for 1 hr. Double labeling of integrin subunit with pankeratin was accomplished through pooling of both primary antibodies into a single incubation and subsequent pooling of both secondary antibodies. Grids were rinsed in diluent for 15 min and then three times in PBS for 5 min each. Grids were placed for 15 min on droplets of half-strength Karnovsky's fixative (filtered through a no. 42 Whatman filter) and rinsed three times for 10 min each in distilled water.
Grids containing the immunogold-labeled sections were placed face down on droplets of ultracentrifuged 1.5% polyvinyl alcohol (PVA)/0.4% uranyl acetate for 2 min, transferred to fresh droplets for 8 min, and picked up using a platinum wire loop. Excess embedding solution was carefully wicked off using Whatman no. 40 filter paper, and the grids were allowed to air dry.
Both the plastic-embedded and immunogold-labeled cryothin sections were imaged using a JEOL 1200 transmission electron microscope operating at 80 Kv. Films were digitized at 1600 DPI using an Epson 1600 flatbed scanner equipped with a transparency adaptor.
Results
Within normal quiescent human skin, the light level staining pattern for integrin subunit β4 shows dominant linear dermal–epidermal junction fluorescence and minor basal keratinocyte lateral cell–cell interface localization (Figures 2A and 2B). Subcellular localization using ultrathin cryosections at the TEM level showed the specific nature of the pattern where β4 localized along the basal plasma membrane of basal keratinocytes, dominantly associating with the hemidesmosome complex along the dermal–epidermal junction (Figures 2C and 2D). β4-positive filopodia were observed as the dominantly labeled feature along the basal keratinocyte lateral cell–cell interface (Figure 2D). β4-positive filopodic contacts gave way to β4-negative desmosomal contacts toward the apical region of the basal cell (data not shown). The frequency of β4 immunogold localization in relation to filopodia seemed to show an inverse relationship to desmosomal density. The dual immunogold label showing both pankeratin (20-nm gold) and β4 (10-nm gold) showed the strong keratin/β4/hemidesmosome association prevalent along the keratinocyte/extracellular matrix interface (Figure 2C).
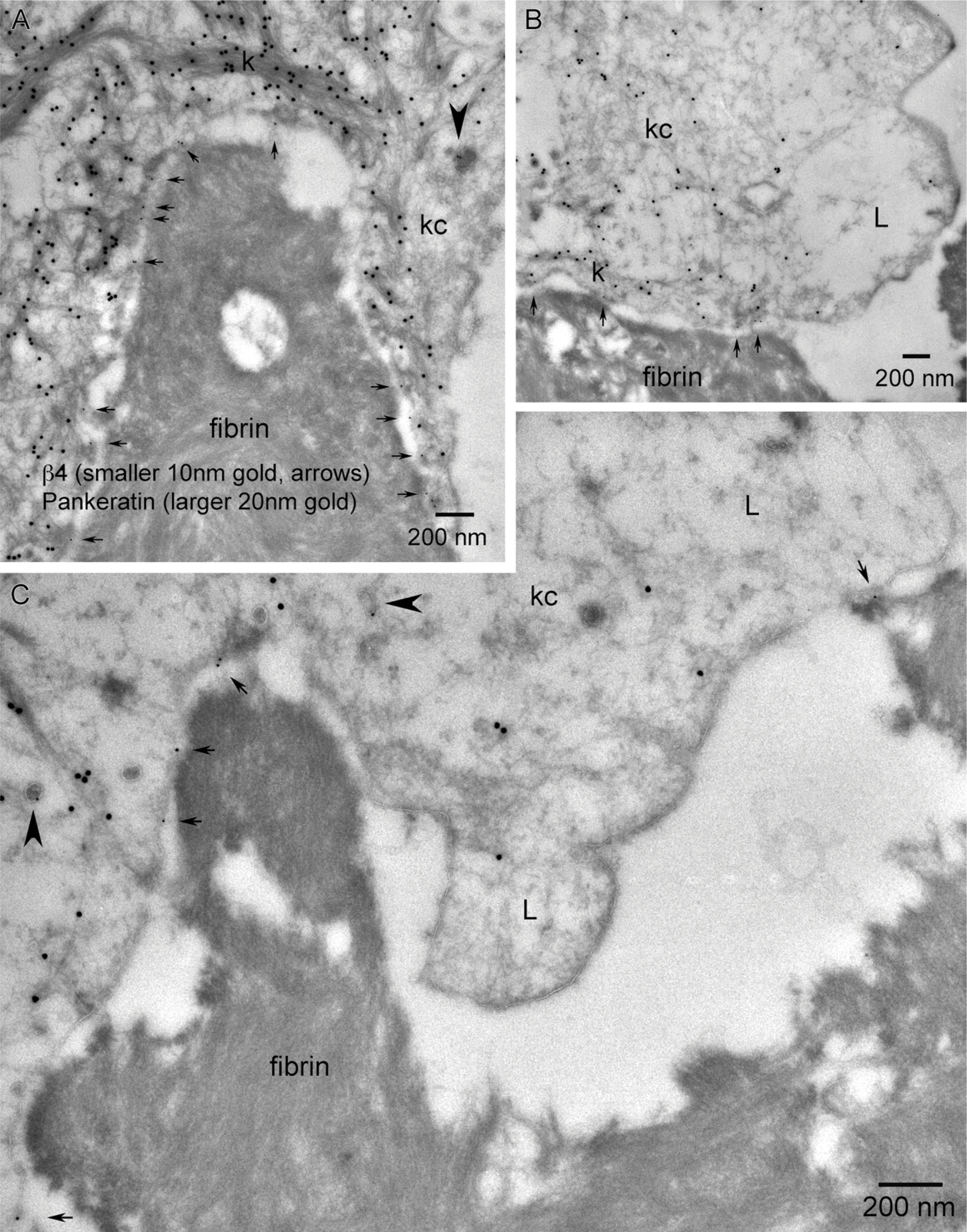
Higher-magnification TEMs within the epithelial migrating tip shows β4 localization in relation to the keratinocyte's interaction with the fibrin network of the wound matrix (trailing vs leading edge). (
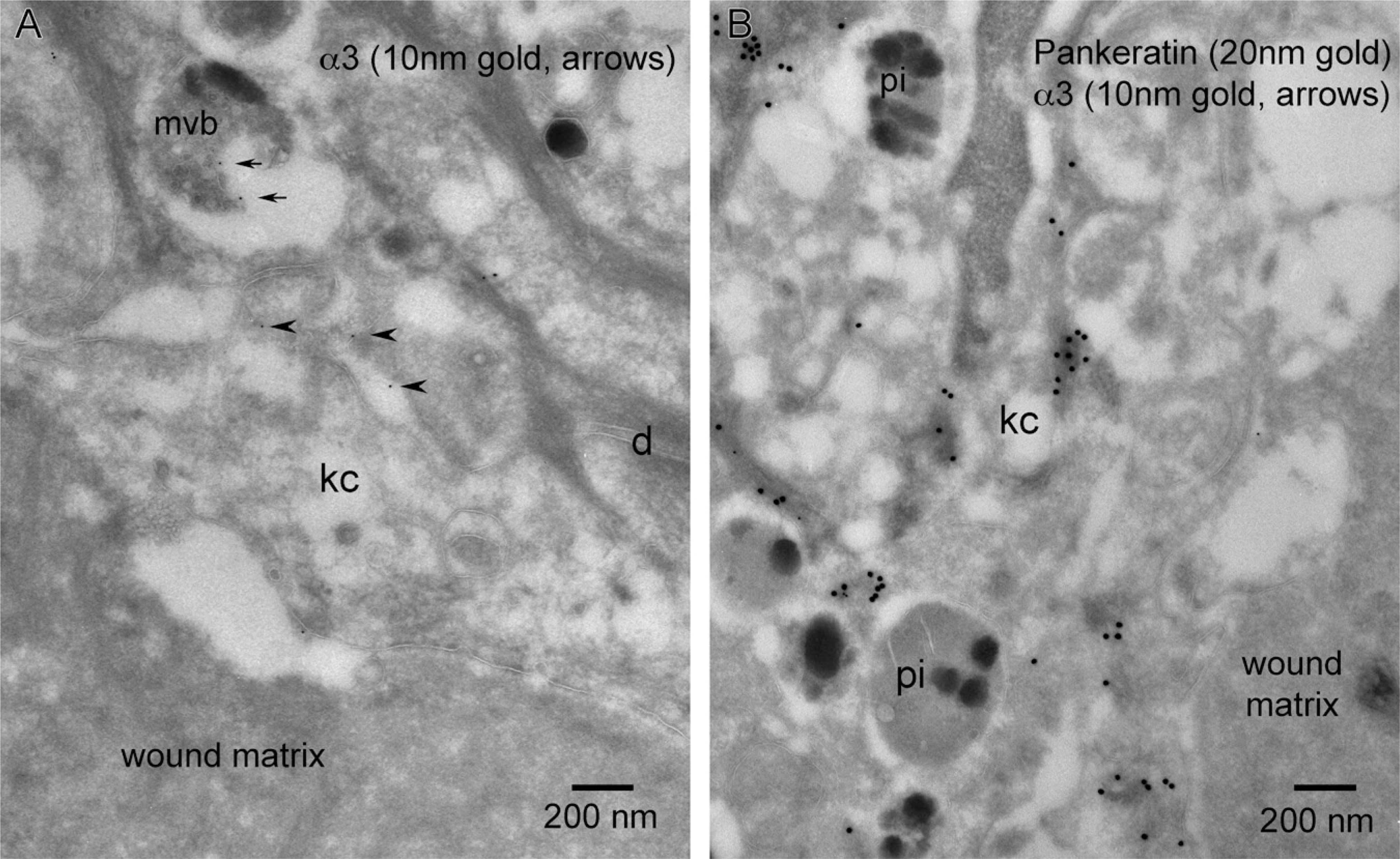
Light level immunofluorescence labeling for integrin subunit α3 showed dominant basal keratinocyte lateral cell–cell interface localization with little or no signal along the dermal–epidermal junction (Figures 3A and 3B). TEM level examination showed α3 (10-nm gold) localizing to the filopodia of the lateral cell–cell interfaces of basal keratinocytes similar to β4 and showing what appeared to be an inverse relationship to desmosomal density (Figures 3C and 3D). We also observed a very sparse frequency of α3 labeling along the basal plasma membrane of basal cells without a topographical relationship to keratin (20-nm gold) or hemidesmosomes (Figure 3C, arrowhead).
A macro view of epithelialization during cutaneous wound healing can be graphically seen in the textural contrast of the epithelial migrating tongue from a burn patient (Figure 4A). Light microscopic examination using semithin cryosections (350 nm) of an experimentally produced 30-hr incisional wound shows a migratory epithelial tongue that can extend 50–400 μm from the original cut edge without contacting the opposing epithelial tongue (Figure 4B). The wound beds are a heterogeneous milieu in their distribution of wound matrix components in juxtaposition to the migrating epithelial tongue. Morphologically, dermis can be seen proximal to the original cut edge, whereas fibrin, collagen, elastin, red blood cells, inflammatory cell infiltrate, serous exudate, and serous crust can be seen within the new wound matrix. We observed variations in ratio and location of wound matrix components from wound to wound (Figure 4B). Although the actual proteinaceous components of the wound matrix are extensive and beyond the scope of this study, the different surfaces of a single keratinocyte may encounter a very different extracellular environment.
Immunolocalization of wounds at the LM level showed β4 predominantly staining basal keratinocytes extending along the advancing wound margin in juxtaposition to the wound matrix (Figure 5C). β4 localization changed from the dominant linear staining along the dermal-epidermal junction and minor lateral basal cell-cell contact staining pattern in adjacent skin (Figure 2) to a discontinuous “beads on a string” (Harper et al. 2005) basal aspect pattern and increased cytoplasmic staining within the basal keratinocytes of the migrating tongue (Figure 5C, arrows). A global TEM morphological view (Figure 5A) using overlapping micrographs and color enhancement to help visualize morphological features and immunogold localization showed a transition zone (T) where the lamina densa of the basement membrane disappears and keratinocytes can be observed migrating in direct juxtaposition to the new wound matrix. Keratinocytes along this region of the transition point (T) are polarized with keratin (20-nm immunogold color enhanced green) concentrated at the trailing edge of the basal keratinocyte, whereas the keratin-depleted lamellipodia (L) can be seen at the leading edge of each keratinocyte in the direction of migration (Figures 5A and 5B). β4 localization (10-nm immunogold color enhanced red) is concentrated in the keratin-rich, trailing edge regions (TRs) along the basement membrane and cytoplasm of basal keratinocytes (Figures 5A and 5B). This pattern of β4 regional keratin-rich concentration and paucity of label within the lamellipodia is conserved within the migrating tip in juxtaposition to the fibrin network (Figures 5D and 5E). The dual immunogold labeling of pankeratin (20-nm gold) and β4 (10-nm gold) further defines the progressive formation of keratinocyte/fibrin-rich matrix contacts within the leading edge of the migrating epithelial tongue. Highest concentrations of β4 are associated with the keratin-rich trailing edge (Figure 6A). β4 remains associated with matrix contacts and bands of keratin along the basal aspect of keratinocytes adjacent to the leading edge lamellipodia (Figure 6B, arrows). Keratin-depleted lamellipodia do, however, exhibit the ability to form discrete β4-positive contacts between a leading keratinocyte and the wound matrix (Figure 6C, arrows). Immunogold localization of newly deposited LM-3A32 at the interface between keratinocytes and wound matrix (Figure 7A, arrows), colocalization of BP-230 and β4 (Figures 7B and 7C), and the presence of anchoring filaments (Figure 7B, arrowhead) define the nature of these contacts as hemidesmosomes, showing that new hemidesmosomes form between keratinocytes and the fibrin-rich wound matrix.
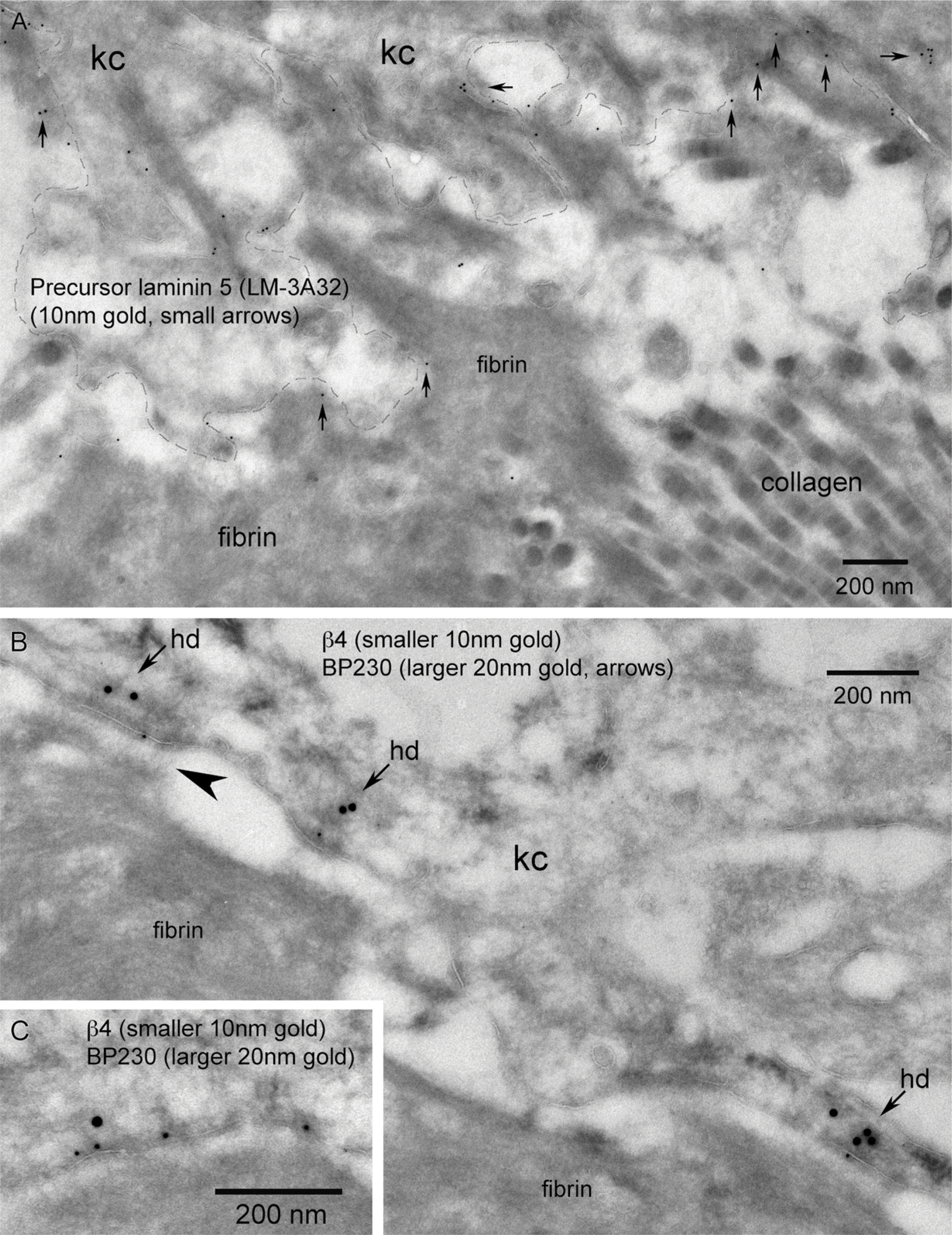
The presence of laminin 5 and BP-230 defines the hemidesmosomal nature of the β4-associated contacts between keratinocytes and the fibrin-rich wound matrix. (
In contrast to β4, LM observation of the α3 staining pattern within the keratinocytes of the migrating tongue in juxtaposition to a fibrin-rich wound matrix showed an overall reduction in signal and no colocalization with BP-230 (Figure 8C). TEM observations showed that the distribution of α3 remains strongly localized to filopodic cell–cell interfaces between keratinocytes adjacent to the migrating epithelial tongue (Figure 8A) and, compared with β4, shows an overall sparse distribution between keratinocytes migrating over the fibrin-rich wound matrix (Figure 8B). α3 concentrations, being proportional to the prevalence of filopodia, is reduced in the global view using overlapping micrographs of the migrating tip of the wound, showing few filopodia between keratinocytes (Figure 8D, circle) and a correspondingly minor distribution of α3. We show a paucity of α3 localization along the trailing edge keratinocyte/basement membrane interface adjacent to the wound (Figures 8A and 9A). This paucity of α3 is also exhibited along the basal aspect of the leading or trailing portions of keratinocytes migrating over the fibrin-rich wound matrix (Figure 9B). There is an absence of α3 within leading edge lamellipodia that are actively forming BP-230–positive hemidesmosomal contacts at the tip of epithelial migration (Figure 9C). However, in contrast to this lack of α3 along the fibrin-rich interface, both α3 and β4 localize to filopodic processes at the keratinocyte/collagen-rich wound matrix interface at the leading edge of the wound epithelial tongue (Figures 10A and 10B).
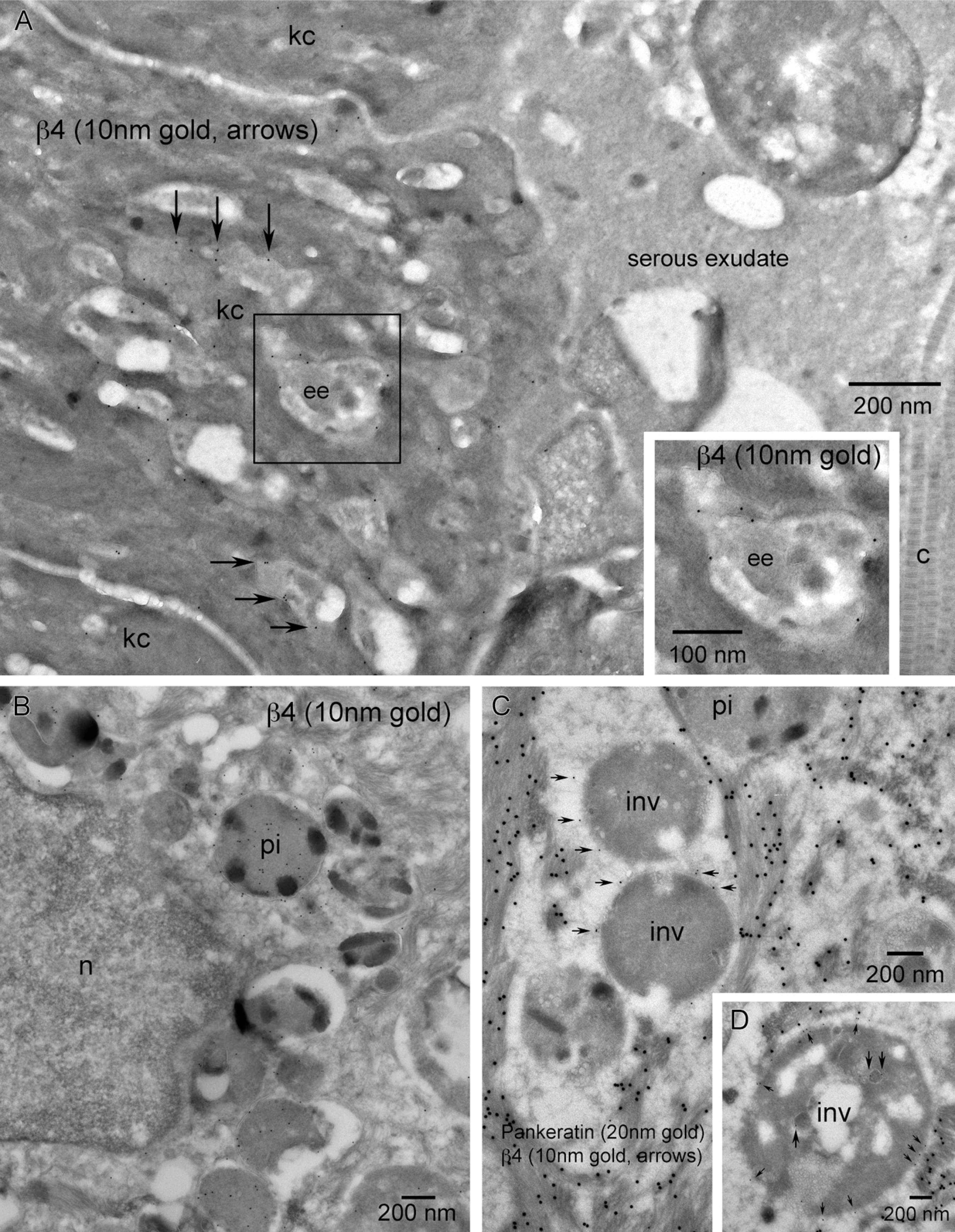
Keratinocytes along the wound margin exhibit an array of cytoplasmic features. Of notable interest in relation to cellular trafficking are vesicular features that appear morphologically in three classes: small endosomes, larger vesicles that could be early/recycling endosomes, and vesicles of the degradative pathway (late endosomes, multivesicular bodies, and phagocytic inclusions; Figure 1B). Localization of α3 within small transport endosomes was rarely observed (Figure 9B, arrowhead). Pigmented phagocytic inclusions (pi) that were often positive for β4 (Figure 11B) were consistently negative for α3 (Figure 12B). On one rare occasion, α3 was observed along the outer membrane of a multivesicular body within a wound basal keratinocyte (Figure 12A, small arrows).
Although the overall α3 vesicular labeling was a rarely observed event, all three classes of vesicles exhibited labeling for β4. Small β4-positive endosomal/transport vesicles were observed at a higher frequency in all wound vs quiescent basal keratinocytes within both the trailing and leading edge cytoplasm (Figures 6A, 6C, and 10A). We made a novel finding of β4 concentrations associated with larger cytoplasmic vesicles within a discrete subset of leading edge keratinocytes. These larger vesicles showed some internal vesicular membranes (Figure 11A). This could be a cross-sectional view through early endosomes exhibiting an undulating outer membrane or late endosomes showing some sequestration or pinching off of the external membrane as it progresses toward becoming a multivesicular body. It must also be noted that this could represent tangential cuts through the basal plasma membrane of the keratinocyte if the plasma membrane is highly undulating (Figure 11A, inset). The degradative pathway also showed β4 labeling within individual subsets of pigmented phagocytic inclusions (Figure 11B). Ultrastructural labeling that may look like perivesicular labeling of non-pigmented inclusions (Figure 11C) is most likely a tangential cut through the dermal–epidermal junction and represents labeling of the basal membrane of the basal keratinocyte. Similar tangential sections showed β4-positive endosomes or caveolae invaginating along the basal plasma membrane (Figure 11D). As a final note, we did not see β4 associated with clathrin-coated pits or clathrin-coated vesicles.
Discussions
We successfully localized integrin subunits β4 and α3 to specific subcellular anatomical structures in both quiescent and wounded skin. Integrin subunit β4 localized to hemidesmosomes and filopodia in normal unwounded skin. After wounding, we observed the additional localization of β4 to lamellipodia and subcellular vesicles ranging from small transport endosomes, possible recycling endosomes, to degradative phagocytic inclusions. Integrin subunit α3 dominantly associated with filopodia both between neighboring keratinocytes and the filopodic interface between keratinocytes and the collagen-rich wound matrix. The results of our study are summarized in illustration form (Figure 13).

Global TEM level distribution of α3 within the epithelial migrating tongue of an incisional wound after 30 hr of healing (
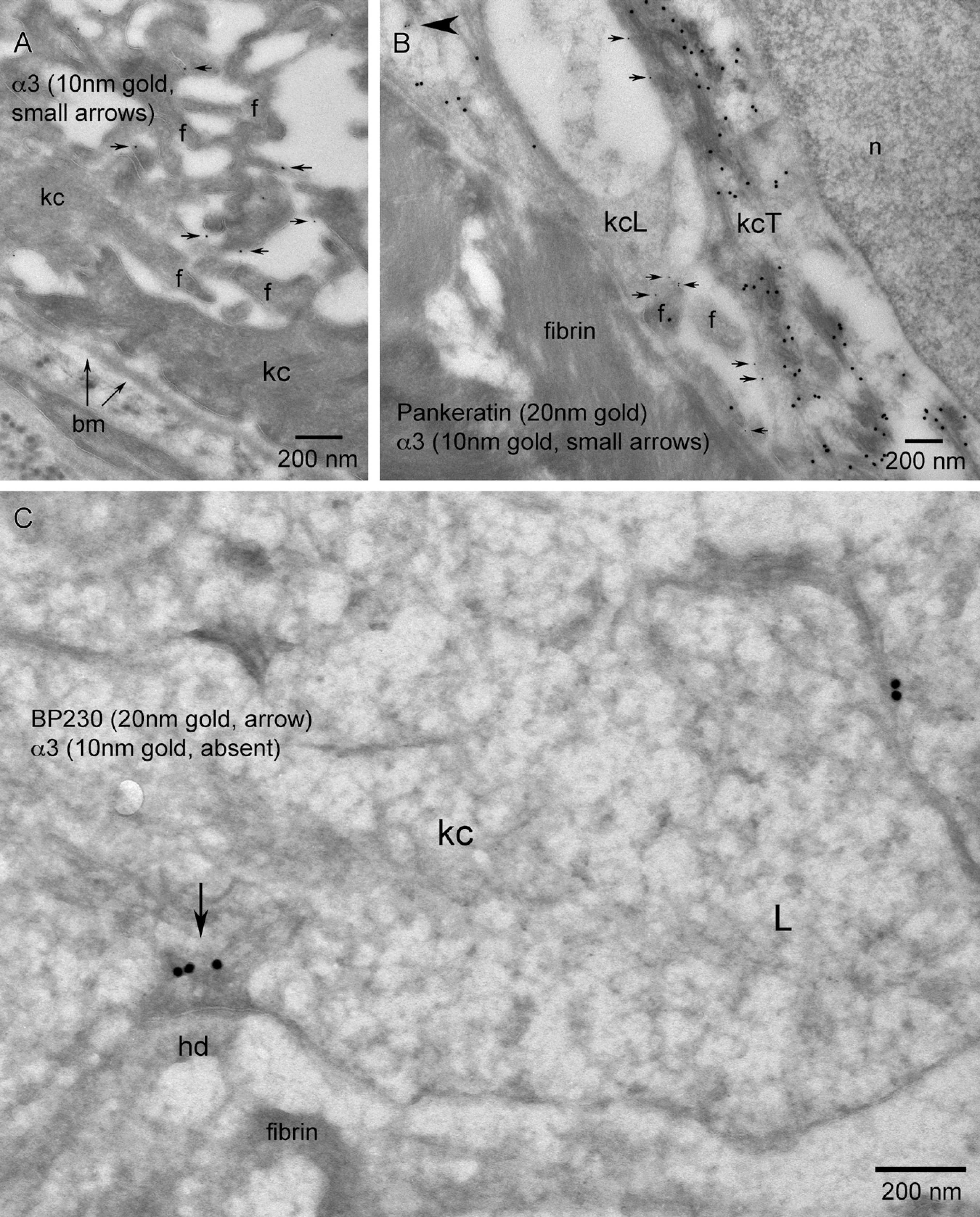
TEM level α3 localization within, first, the region of intact basement membrane adjacent to the transition zone; second, the keratinocyte interaction with the fibrin network of the wound matrix; and third, the leading edge keratinocyte including lamellipodia.
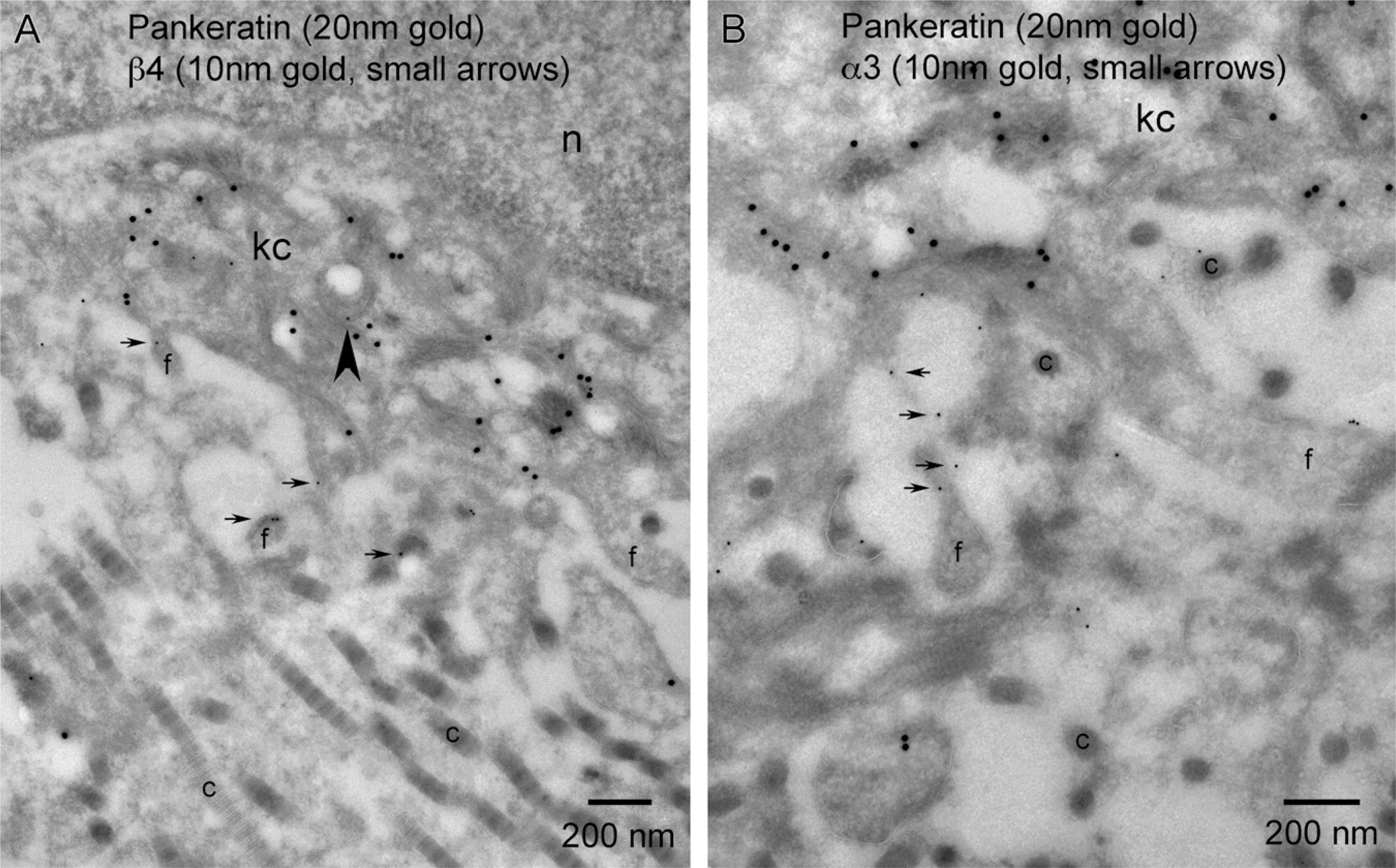
The collagen-rich wound matrix elicits an alternative response from leading edge migrating keratinocytes in regard to expression of β4
Our findings do not support the hypothesis that β4 shifts from a dominant hemidesmosomal association to a dominant filopodic/lamellipodic localization after wounding. Our data suggest that β4 remains strongly associated with keratin/hemidesmosomal adhesions in a trailing edge motif when keratinocytes are migrating juxtaposed to a fibrin-rich wound matrix and remaining associated with filopodia. Our findings also show that β4 is associated with discrete keratinocyte/fibrin contacts at the leading edge of migration within lamellipodic subcellular regions that show a paucity of keratin filaments. Despite this discrete β4 localization to the leading edge, the global concentration of β4 remains within the keratin-rich trailing edge of the migrating keratinocyte. We believe this trailing edge concentration and leading edge paucity of β4 accounts for the repeating cell to cell “beads on a string” (Harper et al. 2005) staining effect when viewing β4 localization on the migrating tongue of wounds at the light level. Our data suggest that the formation of hemidesmosomal contacts between keratinocytes and the wound matrix is a highly dynamic process concentrating traction forming adhesions toward the trailing edge while at the same time creating new stabilizing contacts at the probing leading edge.
Our TEM data support the hypothesis that the expansion of the β4 staining pattern from outer plasma membrane to include cytoplasmic localization in the keratinocytes of the migrating tongue observed at the LM level is caused by increased integrin trafficking. At this stage of our study, we cannot determine inside/out or outside/in directionality of β4 trafficking. We can only conclude that there is increased intracellular trafficking at the wound's leading edge, and this trafficking involves three categories of vesicles: small round non-coated single membrane transport endosomes, larger polymorphic vesicles that could be early or recycling endosomes, and vesicles of the degradative pathway (phagocytic inclusions). It is worth noting that we have observed a tendency of β4 localization to the larger cytoplasmic vesicles in a subset of cells that are juxtaposed to serous exudate and show little sign of actively adhering to wound matrix (Figure 11A). We have not observed β4 in similar vesicles within keratinocytes that are actively adhering through hemidesmosomal contacts or cells exhibiting filopodia juxtaposed to collagen matrix. We require a greater number of samples to see if this is a valid relationship. The overall endosomal distribution of β4 showing a strong association with the keratin-rich regions of the cytoplasm within the migrating tongue suggests a possible connection between zones of intermediate filaments and physical pathways of endosomal trafficking (Styers et al. 2005).
β4 integrin subunit is associated with various cytoplasmic features within a subset of leading edge keratinocytes.
TEM micrographs showing the association of α3 integrin with cytoplasmic vesicles of leading edge keratinocytes was a rarely seen event.
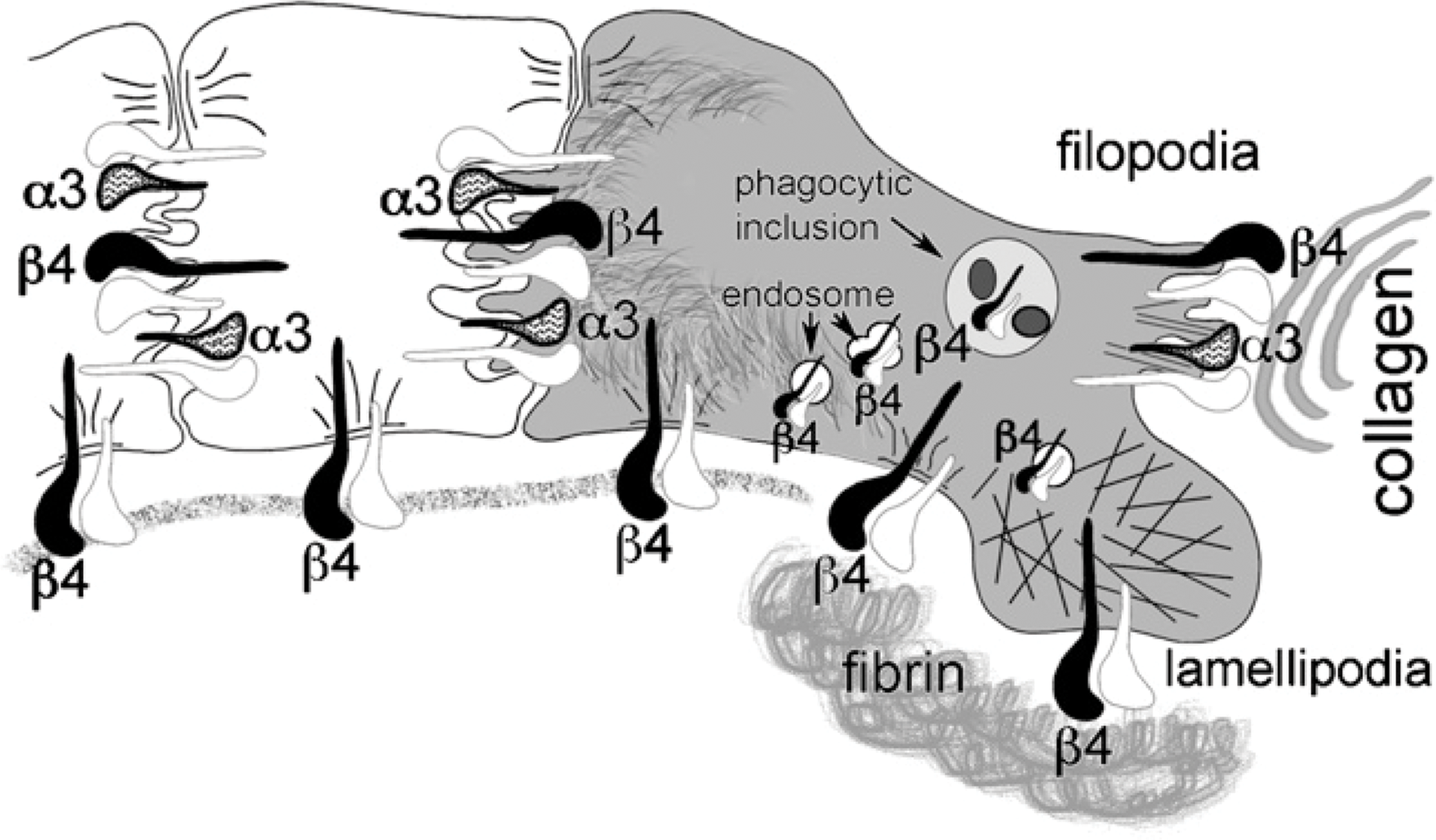
Illustration summarizing our results localizing integrin subunits β4 and α3 in human skin during quiescence (white keratinocytes to the left) and during wound healing in vivo (gray keratinocyte to the right). During quiescence, integrin subunit β4 is located within hemidesmosomal adhesions between basal keratinocytes and the underlying dermis. After wounding, keratinocytes express β4 in hemidesmosomal adhesions between the keratin cytoskeleton and the fibrin-rich wound matrix and to a lesser extent discrete contacts between the keratin-depleted lamellipodia and the fibrin-rich wound matrix. Both β4 and α3 are found on filopodic cell-cell interfaces during quiescence and filopodic cell-cell and cell-collagen-rich matrix interfaces after wounding. The association of the β4 subunit with small endosomal vesicles during quiescence was found to increase after wounding with additional localization to larger polymorphic vesicles and phagocytic inclusions within the leading edge keratinocytes. α3 subunit maintains a discrete minor expression at the keratinocyte's basal interface over both intact basement membrane and wound matrix.
Our data support the hypothesis that α3 can be associated with the interaction of leading edge keratinocytes with a collagen-rich wound matrix (Frank and Carter 2004). We have shown that filopodia express both β4 and α3 integrins wherever they are found; however, in this in vivo study, it seems that the filopodia/integrin subunit/extracellular matrix interface was only observed in the presence of a collagen-rich extracellular matrix. Integrin α3 shifts from a cell-cell interaction proximal to the wound to both a cell-cell and cell-matrix interaction along the migrating wound tongue through filopodia not lamellipodia while rarely showing localization to endosomal vesicles as was commonly observed with β4.
The major overlap between the structural localizations of β4 and α3 was found in filopodia and is suggestive of multiple roles for β4 in stable anchorage (Carter et al. 1990a), transient or traction forming adhesion, and signaling (Santoro and Gaudino 2005). However, this is not exclusive to activated migratory keratinocytes but is exhibited in the quiescent epidermis as well and may not be a shift in function subsequent to wounding but may be the same function applied to a new environment. It would suggest that if hemidesmosomes in the quiescent epidermis are considered stable adhesions exhibiting slow turnover; perhaps the same structure in the wound environment might be a transient fast turnover pinning from which lamellipodia can extend themselves.
When localizing integrins to cellular structures such as hemidesmosomes, it is important to remember that the structural relationship we visualize is in fact a relationship between an integrin and its ligand. The electron-dense contrasted matrix identified ultrastructurally as fibrin (Odland and Ross 1968; Ross and Odland 1968) is actually composed of many different components (Clark 2001, 2003). The fact that there is no receptor identified on keratinocytes for fibrin (Clark 2003; Geer and Andreadis 2003) does not prevent the cells from forming hemidesmosomal adhesions to an intermediary component associated with the fibrin rich network, possibly plasmin-mediated fibrin degradation (Geer and Andreadis 2003) and newly deposited LM-3A32 (Nguyen et al. 2000b). The presence of anchoring filaments, LM-3A32, keratin concentrations and colocalization of β4 with BP230 at the adhesion sites identifies these structures as hemidesmosomes (Ishiko et al. 1998).
The electron dense material found in vesicles within the cytoplasm of leading edge keratinocytes is likely from three origins: lipofuscin and hemosiderin, both within the degradative pathway, and naturally occurring melanin pigment. Melanocytes were frequently observed in the migrating tongue of wounds (Figure 5A). Vesicles of the degradative pathway showed a variety of morphologies suggestive of stages from late endosomes, multivesicular bodies, and phagocytic inclusions. Localization of β4 within the larger cytoplasmic vesicles of keratinocytes at the leading edge of the migrating tongue (Figure 11A) may be caused by integrin sorting within early/recycling endosomal compartments or degradative processing within late endosomal and phagocytic/lysosomal vesicles. However, it could be interpreted as a tangential cut through a region of highly undulating plasma membrane of keratinocytes interacting with adjacent collagen. No fibrin-rich matrix was observed and thus would discount the possibility of β4 concentrations associated with hemidesmosomal contacts. We believe it is clear that some of the β4 is being degraded in identifiable pigmented phagocytic inclusions (Figure 11B) as result of rho GTPase activation inducing phagocytic activity in the migratory motif (Fiorentini et al. 2001).
Engaging the challenges of using ultrathin cryomicrotomy to examine a small, heterogeneous region of tissue has helped refine our techniques and to position us to pursue the next stage of experiments. We suggest that in vivo ultrastructural snapshots using markers targeting Rab proteins demarcating the integrin endosomal/exosomal pathways will become an important avenue of discovery (Bretscher 1992; Hunziker and Peters 1998; Zacchi et al. 1998; Yoon et al. 2005), as well as degradative markers such as the degradation resistant cytosolic enzyme CuZn-superoxide dismutase (SOD) compared with the rapidly degraded carbonic anhydrase (CAIII) will help clarify the localization of β4 in the degradative pathway (Fengsrud et al. 1995). The cellular management of integrins is central to migration, polarity, and adhesion of keratinocytes after wounding (Caswell and Norman 2006) and may provide insights not only into normal healing, but the impaired non-migratory nature of chronic wounds as seen in patients with diabetes (Olerud et al. 1995). These same issues become important in the context of cutaneous interaction with percutaneous biomaterial implants (Knowles et al. 2005; Fukano et al. 2006) such as catheters where the same integrin recycling pathways are involved in forming or not forming a seal around the implant and are also used by pathogens to gain access to the living tissues (Pellinen and Ivaska 2006; Pizarro-Cerda and Cossart 2006). Our long-term goal is to better understand the migration/adhesion characteristics of keratinocytes to understand the basic biology of wound healing, to promote healing of chronic ulcers, and to encourage a pathogen barrier to form at the interface between the epidermis and a percutaneous device (Fukano et al. 2006).
“It is only the intact organism that constructs the complete network of interactions, and there is a need for good in vivo studies to compliment increasing molecular and biochemical studies” (Lindblad 2006). We believe that ultrathin cryomicrotomy coupled with immunogold localization of β4, α3, BP230, LM-3A32, and keratin has served, in a provocative way, not possible using light microscopy, to show some key subcellular features of wound healing in vivo.
Footnotes
Acknowledgements
This study was supported by The George F. Odland Endowed Research Fund, National Institutes of Health Grants DK-59221 and EB-004422, and National Science Foundation Grant EEC 9529161 (University of Washington Engineered Biomaterials).
We thank Dr. John Stanley of the University of Pennsylvania for his generous gift of the antibody directed against BP230. This article is dedicated to Frank Hutchinson, who encouraged me to look at the world in closer detail.