
Abstract
The tetra-membrane-spanning protein CD9 forms a complex with a membrane-anchored heparin binding epidermal growth factor-like growth factor (HB-EGF) and integrin α3β1 in some human and monkey cell lines. We show here the immunohistochemical distribution of CD9, HB-EGF, and integrin α3β1 in normal human tissues. Distribution of CD9, HB-EGF, and integrin α3β1 was similar in various tissues, including transitional epithelium, squamous epithelium, thyroid follicular epithelium, adrenal cortex, testis, smooth muscle, and stromal fibrous tissue. However, distribution of the three proteins did not coincide in some tissues, such as lung, liver, kidney, gastric and intestinal epithelium, pancreas, salivary gland, and ovary. In striated muscle, including cardiac muscle, CD9 was present not in the muscle cells themselves but in the endomysium and perimysium, whereas HB-EGF was distributed in the muscle cells themselves. CD9 was distributed in the myelin, but HB-EGF was found in the axon of the peripheral and central nervous systems. Coincident distribution of integrin α3β1 with others was not observed in muscles and neural tissues. In conclusion, there is a possibility of complex formation and functional cooperation of CD9 with HB-EGF and/or integrin α3β1 in several tissues.
CD9 and members of the tetra-membrane-spanning superfamily, including CD37, CD53, CD63, CD81, CD82, and CD151, are implicated in cell growth, cell motility, and cell adhesion in various cells (Boucheix et al. 1991; Horejsi and Vlcek 1991; Lanza et al. 1991; Mitamura et al. 1992; Sincock et al. 1997). CD9 was originally identified as a surface antigen of lymphohemopoietic cells (Boucheix and Benoit 1988). However, it has also been detected in a wide variety of nonlymphohemopoietic tissues, including tissues of the central and peripheral nervous systems (Kersey et al. 1981; Komada et al. 1983; Nakamura et al. 1996; Schmidt et al. 1996; Kagawa et al. 1997). It has been shown that expression of CD9, originally called motility-related protein-1 (MRP-1), is reduced in some neoplastic tissues and is inversely related to the prognosis of patients with neoplasms (Miyake et al. 1991; Ikeyama et al. 1993; Huang et al. 1998). Our recent study of CD9 null mice has demonstrated that CD9 is a crucial factor for fertilization in mice oocytes. Therefore, CD9 knockout females are mostly infertile because of the failure of the sperm and egg fusion step in fertilization (Miyado et al. 2000).
It has been shown that CD9 uniquely associates with a membrane-anchored form of heparin binding epidermal growth factor-like growth factor (proHB-EGF) and upregulates the biological activity of proHB-EGF (Iwamoto et al. 1994; Higashiyama et al. 1995). The soluble form of HB-EGF (sHB-EGF) is a potent mitogen for epithelial cells, fibroblasts, and vascular smooth muscle cells (Higashiyama et al. 1992). CD9, like other tetra-membrane-spanning proteins, forms complexes with many other membrane proteins, including multiple integrins such as α3β1 and α6β1 (Nakamura et al. 1995; Berditchevski et al. 1996). Similarly, integrin α3β1 forms complexes with tetra-membrane-spanning proteins other than CD9, such as CD63, CD81, CD82, and CD151 (Berditchevski and Odintsova 1999; Sincock et al. 1999). The integrin family of cell adhesion receptors is important for the regulation of cell adhesion, cell proliferation, cell migration, and tumor cell metastasis (Schwartz et al. 1995). Therefore, CD9 may function to interconnect between the membrane-anchored growth factor and the adhesion molecule to facilitate intercellular signaling with neighboring cells (Higashiyama et al. 1995; Nakamura et al. 1995).
We report here the distribution of the three proteins, CD9, HB-EGF, and one of the integrins (integrin α3β1), in human tissues using immunohistochemistry.
Materials and Methods
Seven adult autopsy cases (31–75 years old) and three neonatal cases (27 weeks, 35 weeks, and 41 weeks of gestation) comprised the material of this study. Fresh tissues such as lung, heart, liver, kidney, esophagus, GI tract, pancreas, adrenal gland, thyroid gland, salivary gland, urinary bladder, testis, ovary, brain, spinal cord, and sciatic nerve were frozen-sectioned (6 μm thick) and fixed briefly in acetone (5 min) for immunohistochemistry. After freeze-sectioning, the rest of the tissues were fixed in 20% formalin and embedded in paraffin. Some tissues were immediately placed in liquid nitrogen for protein extraction.
Histological Staining
Formalin-fixed, paraffin-embedded sections were stained with hematoxylin and eosin. Some tissue sections were stained with special stainings such as Azan for collagen fiber, elastica van Gieson for elastic fibers and muscle cells, PAS for epithelial mucin, silver impregnation for reticulin fibers, Bodian silver stain for neurofilaments, and Luxol Fast Blue for myelin.
Antibodies
A mouse monoclonal antibody (MAb) 007 was prepared as described previously (Iwamoto et al. 1991). A rabbit polyclonal antibody H6, which recognizes the extracellular domain of HB-EGF, was prepared as described previously (Iwamoto et al. 1994). A mouse MAb against human integrin α3β1 was also commercially obtained (Chemicon International, Temecula, CA; catalogue number MAB1992). The MAb recognizes an epitope related to VLA-3 complex (α3β1). Both CD9 and integrin α3β1 MAbs were purified IgG class antibodies.
Other commercially available antibodies were used as markers for normal tissues and cell types, such as pan-cytokeratin (Boehringer–Mannheim; Indianapolis, IN) and epithelial membrane antigen (DAKO; Carpinteria, CA) for epithelial cells, myoglobin (DAKO), desmin (DAKO), and α-smooth muscle actin (Novo Castra; Newcastle, UK) for muscle tissues, neurofilament protein (DAKO) for neuronal fibers, myelin basic protein (DAKO) for myelin, S-100 protein (DAKO) for peripheral nerves, vimentin (DAKO) and fibronectin (DAKO) for mesenchymal cells including perimysium and endomysium, and collagen Type IV (DAKO) for basement membranes.
Immunohistochemistry
The freeze-sectioned and acetone-fixed specimens were sequentially incubated at room temperature with various diluted antisera for 20 min, 3% hydrogen peroxide for 5 min, biotinylated secondary antisera (rabbit or mouse; BioGenex, San Ramon, CA) for 20 min, and streptavidin–enzyme conjugate (BioGenex) for 20 min. Each incubation was followed by three 5-min washes in PBS. Sections were exposed to the chromogen reaction solution (3-amino-ethyl carbazole; Lipshaw, Detroit, MI) for 10 min. After washing in tapwater, specimens were briefly counterstained. Immunohistochemistry with specific markers for various tissues was done using serial tissue sections.
Dilutions of primary antibodies were 1:50 (H6) and 1:100 (007, integrin α3β1, and all other commercially available antibodies).
As negative controls, primary antibodies were omitted or normal serum substituted.
Western Blotting
Tissues stored in liquid nitrogen were dispersed with the polytron and lysed with lysis buffer (10 mM HEPES, pH 7.0, 0.5 M NaCl, 1% Triton X-100, 0.02% SDS, 3 mM PMSF). After centrifugation at 200, 000 × g for 30 min, the supernatants were used for immunoblotting. An aliquot containing 20 μg protein was boiled with SDS-PAGE sample buffer. Samples were electrophoresed in an SDS-polyacrylamide gel (15%), transferred to an Immobilon membrane, and blotted with 007 and H6 as described previously (Iwamoto et al. 1991). Immunopositive bands were detected by ECL Western blotting kit (Amersham; Poole, UK).
Results
Immunohistochemistry
The distribution of each antigen was carefully evaluated. A summary of the immunoreactivities is shown in Table 1.
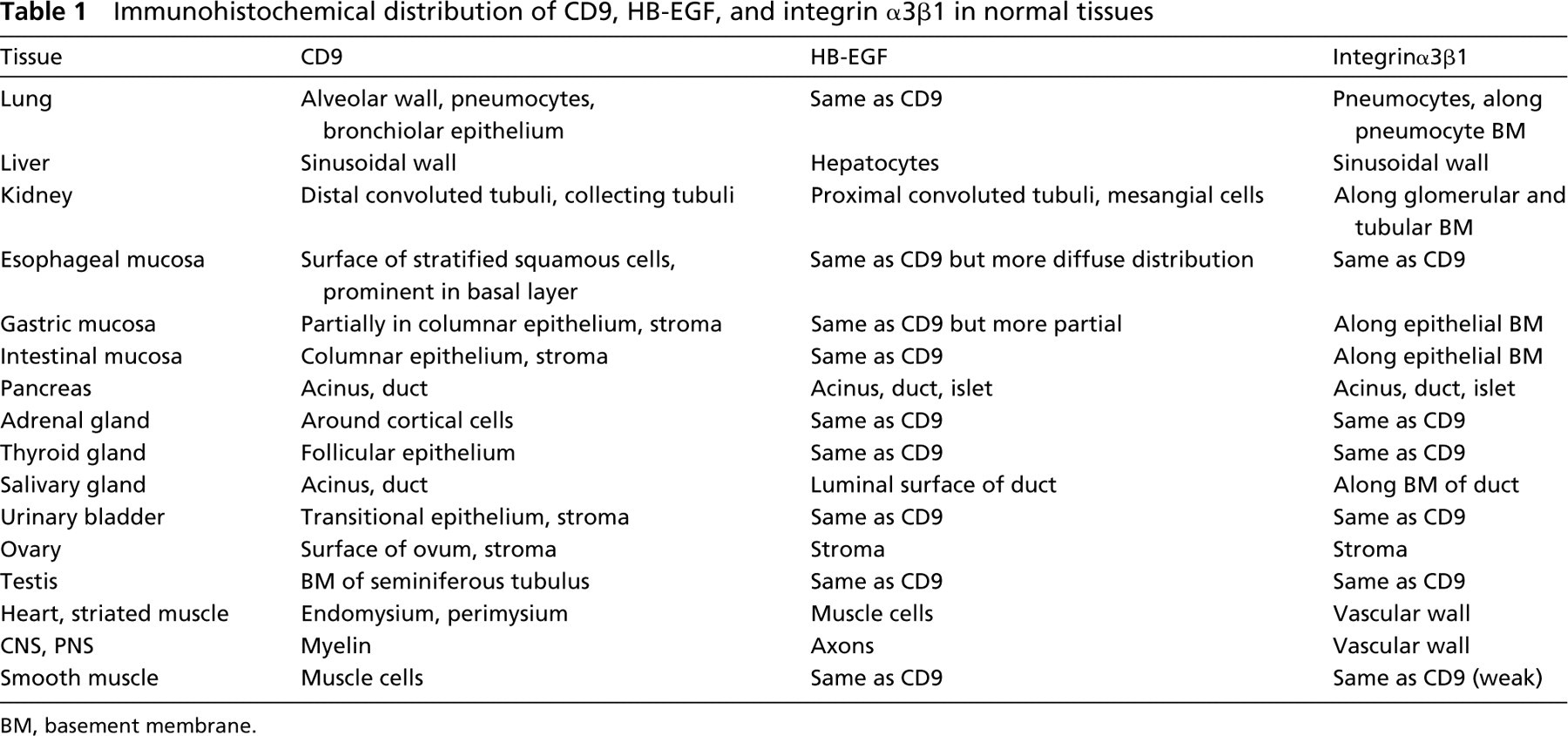
The distribution of CD9, HB-EGF and integrin α3β1 was similar in various tissues, such as transitional epithelium, squamous epithelium, thyroid follicular epithelium, adrenal cortex, testis, smooth muscle, and stromal fibrous tissue, in all neonatal and adult cases. CD9, HB-EGF and integrin α3β1 were positively stained along the cell surface of the transitional epithelium in the urinary bladder and ureter. They were present around the cortical epithelial cell surface in the adrenal gland, follicular cells in the thyroid gland, and along the basement membrane (BM) of the seminiferous tubulus in the testis. They were all positive along the cell surface of the smooth muscle cells (Figure 1), which were positive for desmin and α-smooth muscle actin, and stromal fibrous tissue, which was positive for vimentin and/or fibronectin.
Immunohistochemical distribution of CD9, HB-EGF, and integrin α3β1 in normal tissues
BM, basement membrane.
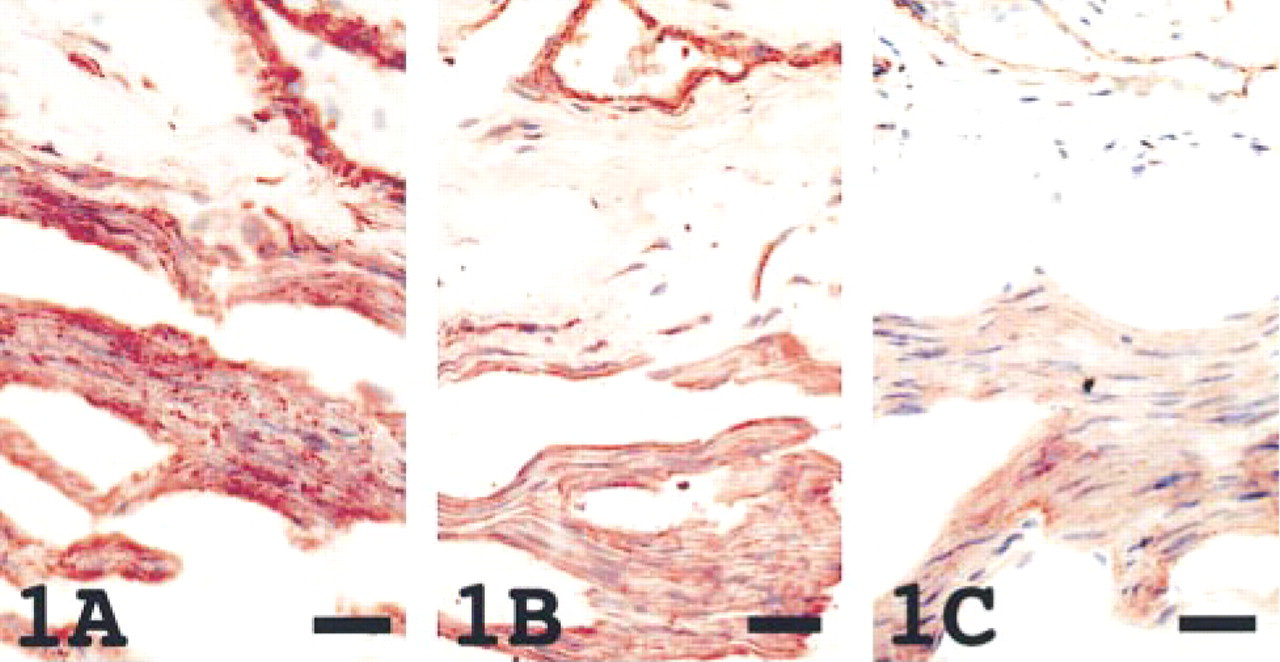
Distribution was a little different among the three antigens in tissues, including lung, liver, kidney, gastric and intestinal epithelium, pancreas, salivary gland, and ovary. In the lung, CD9 and HB-EGF were detected in the alveolar wall, pneumocytes, and bronchiolar epithelium, but integrin α3β1 was predominently positive in pneumocytes. The number of pneumocytes with positive immunoreactivity for the three antigens was greater in the neonatal lung compared to the adult lung. In the liver, CD9 and integrin α3β1 were weakly positive along the sinusoidal wall, whereas HB-EGF stained positively in small numbers of hepatocytes. Whereas CD9 was present in distal convoluted tubuli, collecting tubuli, and mesangial cells, HB-EGF was seen in proximal convoluted tubuli and mesangial cells, and integrin α3β1 along the glomerular and tubular BM in the adult kidney (Figure 2). In neonatal kidney, an immature nephrogenic zone without positive staining of the three antigens was present in the superficial cortex. In the gastric and intestinal mucosa, CD9 and HB-EGF were positive along the cell surface of the columnar epithelium, and integrin α3β1 was found along the epithelial BM. In the pancreas, HB-EGF and integrin α3β1 were present in the acinus, duct, and islet cells, but CD9 was seen only in the acinus and duct. In the salivary gland, CD9 was present in the acinus and duct, HB-EGF on the luminal surface of the duct, and integrin α3β1 along the BM of the duct. These epithelial cells were also positive for cytokeratin and/or epithelial membrane protein, the stromal cells positive for vimentin and/or fibronectin, and the basement membrane positive for collagen Type IV. Whereas the CD9 was present on the surface of the ovum, HB-EGF and integrin α3β1 were seen only in the stroma of the ovary. Integrin α3β1 was found similar to the HB-EGF, but the intensity of staining was weak.
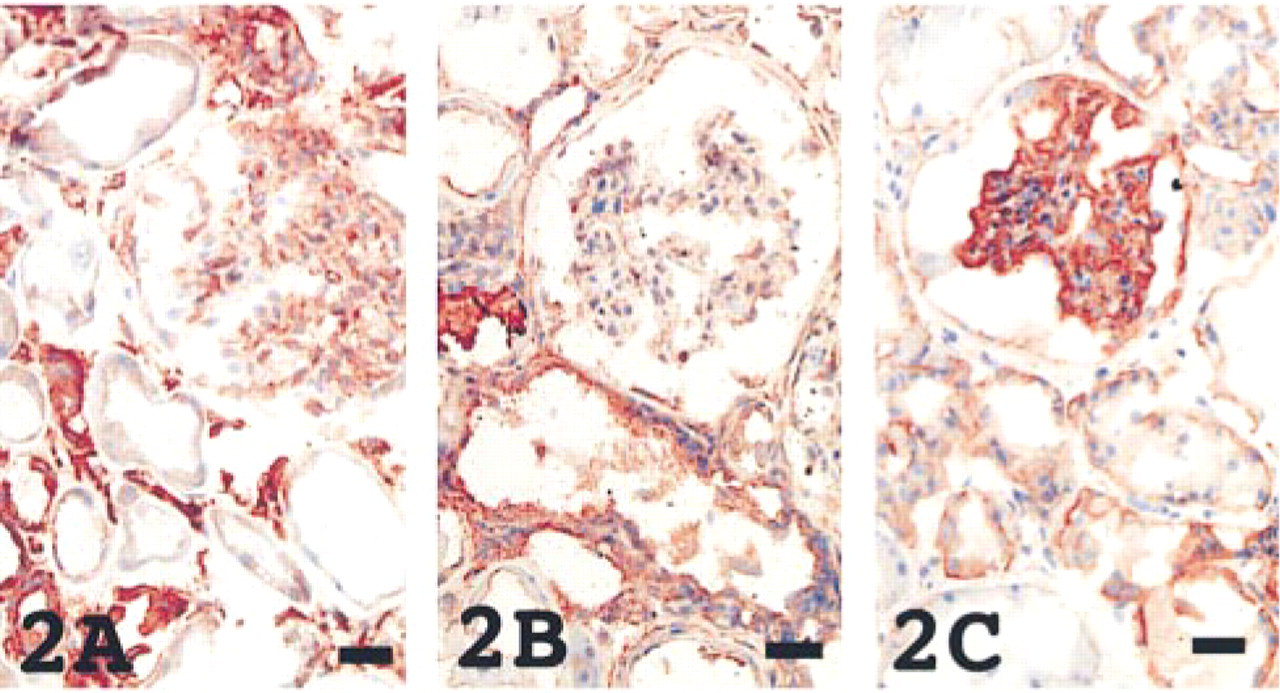
In both neonatal and adult striated muscle, including cardiac muscle, CD9 was present not in the muscle cells themselves but in the endomysium and perimysium (Figure 3) which were positive for vimentin and/or fibronectin. On the contrary, HB-EGF was distributed in the muscle cells themselves, which were positive for myoglobin. Integrin α3β1 was found only in the vascular wall.
In tissues of the central and peripheral nervous systems, the distribution of CD9 and HB-EGF was characteristic. Whereas CD9 was distributed in the myelin that was positive for myelin basic protein, HB-EGF was found in axons of the peripheral and central nervous systems that were positive for neurofilaments (Figure 4). Integrin α3β1 was not associated with CD9 and HB-EGF in the neural tissue. In neonatal brains, unmyelinated nerve fibers were not positively stained with CD9 as previously reported (Nakamura et al. 1995).
Western Blotting
Anti-CD9 MAb 007 detected a single band of about 29 kD in the lung, esophagus, ovary, and salivary gland. In the heart and liver, signals were faint. In the spinal cord, doublet bands were detected as previously reported (Nakamura et al. 1996). Although the reason for the doublets is not clear, it may be due to the differences in glycosylation (Figure 5A). Anti-HB-EGF MAb H6 detected a band of about 29 kD and a nonspecific extra band with second antibody in all samples and bands of about 49 kD and 120 kD in some samples (Figure 5B).
Smooth muscle cells are positively stained in the cytoplasm with CD9 (
The kidney revealed CD9 (
The heart showed different immunoreactivities; CD9 (
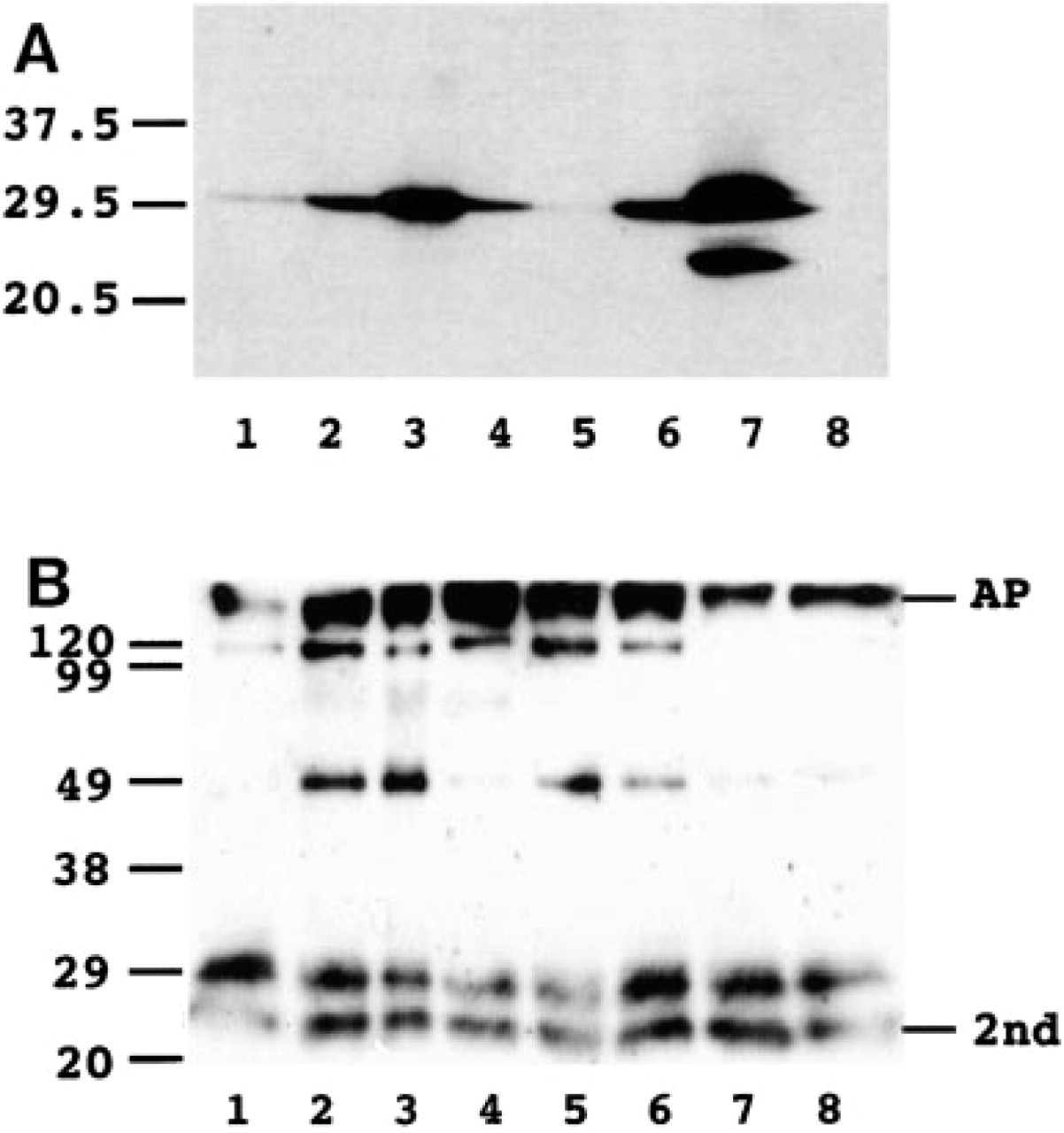
Western blotting analysis of expression of CD9 (
Discussion
This study showed that CD9, HB-EGF and integrin α3β1 were coincidently distributed in transitional epithelium, squamous epithelium, thyroid follicular epithelium, adrenal cortex, testis, smooth muscle, and stromal fibrous tissue. In these tissues, the three proteins might be interrelated as a functional unit. In epithelial tissues such as transitional, thyroid follicular, and adrenal cortical epithelium, the three proteins may be important for reciprocal epithelial cell attachment. The association of CD9 and integrin α3β1 in normal and neoplastic human keratinocytes has been reported (Jones et al. 1996; Okochi et al. 1997). It is also suggested that CD9 could make a complex with proHB-EGF and cooperate with it for human keratinocyte growth in a juxtacrine manner (Inui et al. 1997). Abnormalities of these proteins in neoplastic epithelial cells may lead to detachment, invasion, and metastasis. A poor prognosis has been reported in cases of CD9-negative cancers arising in the breast, colon, lung, and esophagus (Miyake et al. 1996; Higashiyama et al. 1997; Mori et al. 1998; Uchida et al. 1999). One report described the co-localization of tetraspan superfamily such as CD151, CD9 and CD63, and integrin α5β1 in some normal tissues using immunohistochemistry (Sincock et al. 1997).
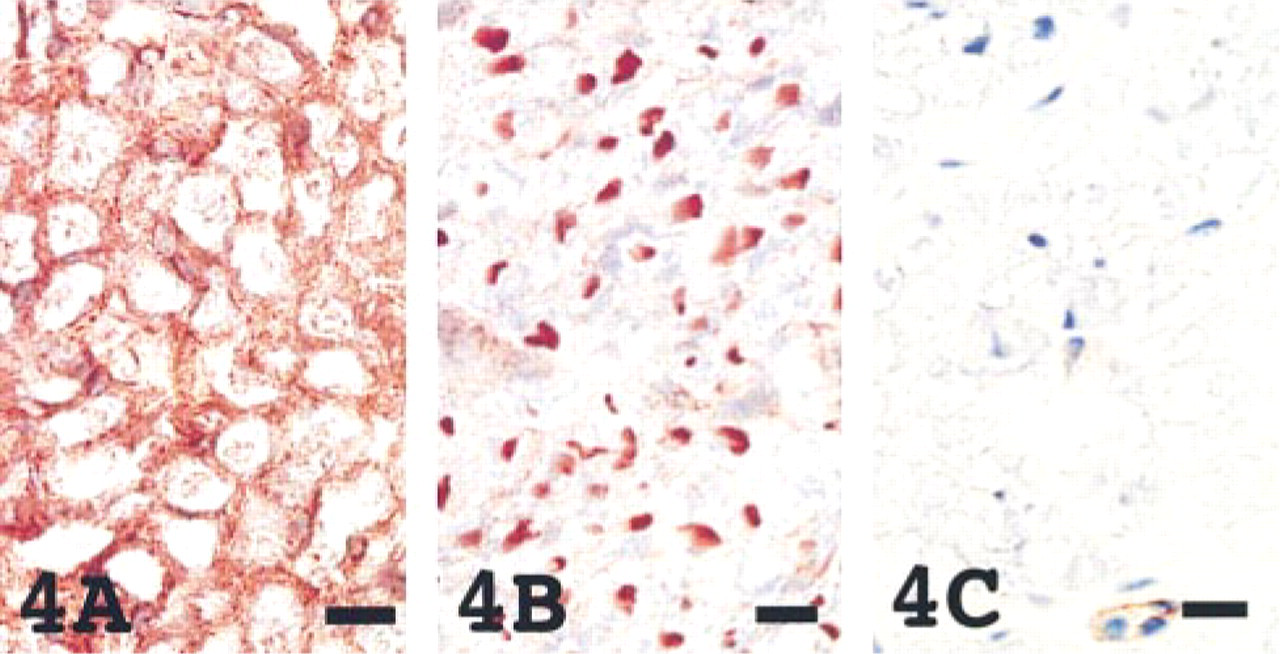
In the spinal cord, CD9 (
The CD9 was associated with HB-EGF and integrin α3β1 in smooth muscle cells, however CD9 was proved to be present solely in the endomysium and perimysium of the striated muscle cells. It is reported that HB-EGF is present in the smooth muscle of the urinary bladder and is responsible for muscle hyperplasia in pathological conditions as an autocrine smooth muscle cell growth factor (Borer et al. 1999). Although a relation to CD9 and/or integrins has not been reported, it is possible that CD9 and/or integrin α3β1 plays a role in the regulation of the function of HB-EGF on smooth muscle proliferation. One report suggested that CD9 and CD9–β1 integrin complexes are important for muscle cell fusion and support myotube maintenance (Tachibana and Hemler 1999). In this regard, CD9, which presents in the endomysium and perimysium, may be responsible for striated muscle fusion and maintenance.
In tissues such as the lung, liver, kidney, pancreas, salivary gland, and ovary, the distribution of CD9, HB-EGF, and integrin α3β1 differed, and the significance of this is unclear. We have demonstrated that CD9 expressed on the surface of the ovum is important for fertilization (Miyado et al. 2000). This study has confirmed the presence of CD9, but HB-EGF and integrin α3β1 were not found on the surface of the ovum. However, on the surface of the ovum CD9 has been proved to exist with α6β1 integrin and to play an important role in fertilization (Miyado et al. 2000). In the kidney, CD9, HB-EGF, and integrin α3β1 were distributed in different portions of the tubuli. One report showed that transfection of cultured renal epithelial cells with HB-EGF and CD9 increased renal epithelial survival, suggesting that they are more resistent to disruption of cell–cell and cell–matrix interactions and acceleration of the re-establishment of these attachments (Takemura et al. 1999).
In central and peripheral neural tissues, we have demonstrated that CD9 is abundant in myelin, localizing along its outermost membrane (Nakamura et al. 1996). Although the function of CD9 in myelin has not been demonstrated, the abundant expression of CD9 suggests a unique function of CD9 along the surface of myelin. One report suggested that CD9 is associated with a neural adhesion molecule, L1, and with integrin α6β1 in mouse brain, and is implicated in neurite outgrowth and cell migration in vitro (Schmidt et al. 1996). It has been reported that both neurons and glia express HB-EGF mRNA in the rat forebrain (Mishima et al. 1996). This suggests that the HB-EGF plays an important role in the brain. This report is the first report demonstrating the presence of HB-EGF in axons of the central and peripheral nervous systems.
The distribution of CD9, HB-EGF, and integrin α3β1 in normal human tissues would provide fundamental data for study of the functional significance of complex proteins.
Footnotes
Acknowledgements
Supported in part by Health Sciences Research Grants for Specific Diseases “Intractable Hydrocephalus” (1999-SD-17) from the Ministry of Health and Welfare, Japan (to YN).
We thank the staffs of Department of Pathology, St. Mary's Hospital, for their assistance.