
Abstract
Stratified epithelia are exposed to abrasive forces and are required to respond rapidly to injury to minimize fluid loss and the risk for microbial infection. Healing involves a cell migratory phase to reestablish barrier function and cell proliferation to re-stratify the epithelium. Cell migration during re-epithelialization involves cell sliding, termed sheet movement, during which cells retain their cell-cell junctions while dynamically altering their shape and cell-substrate interactions to permit movement across the exposed wound bed. Proteins of the integrin family of receptor molecules modulate cell shape, cell migration, and signal transduction in many cell types. In epithelial cells, integrins of the β1 family have been implicated in regulating cell proliferation and differentiation. α9βb1 is one of the newer members of the integrin β1 family and has been recently shown to function as a tenascin receptor. Although little is known about its function in vivo, studies in developing mouse cornea and eyelid suggest that it may play a role in epithelial differentiation. Using a debridement wound model in the mouse cornea, we show in this study that (a) in response to small debridement wounds that close without cell proliferation, α9 integrin protein and mRNA are not induced during migration but are induced during restratification, (b) larger debridement wounds that require cell proliferation to generate the cells necessary for sheet movement result in a dramatic induction of α9 protein and its mRNA during both migration and restratification, and (c) tenascin, an α9β1 ligand, accumulates beneath epithelial cells during restratification but not during cell migration. Therefore, α9 integrin protein production and tenascin accumulation are dynamically regulated in response to corneal epithelial injury.
I
Recently we reported that among the β1-family integrins expressed in the epithelial cells of the skin and cornea, only one, the α9 subunit, localized differently in the skin compared to the cornea (Stepp et al. 1995). α9 integrin forms heterodimers with the β1 integrin subunit and in adults localizes to the basal cells of the epidermis, skeletal muscle, hepatocytes, to the airway epithelium lining the lungs, and to the basal cells of the corneal limbus. The only well-characterized ligand for α9β1 integrin is tenascin (Yokosaki et al. 1994). Tenascin is a large hexameric protein with multiple functional domains, whose expression has been correlated with development and wound healing (Lightner 1994; Erickson 1993).
In the epidermis, α9 integrin is localized to the lateral, apical, and basal cell membranes of the epithelial basal cells. In the cornea, α9 is localized to the basal cells within the limbus, a region of the cornea at its periphery that forms its border with the conjunctiva. Stem cells in skin are distributed in clusters and comprise approximately 10% of the cells in the basal cell layer (Potten and Morris 1988). The stem cells that produce the central corneal epithelial cells are located at the limbus (Zieske et al. 1992, 1994; Cotsarelis et al. 1989; Schermer et al. 1986). Proliferation of the limbal basal cells yields daughters which are either stem cells or transit-amplifying (TA) cells. Initially, TA cells have high proliferative potentials. In the normal cornea, TA cells migrate away from the limbus along the basement membrane towards the center of the cornea. As new cells are needed to replace squames shed from the surface of the cornea, TA cells proliferate, generating daughter cells that tend to terminally differentiate into cells that leave the basement membrane. The observation that the epithelial stem cells in the cornea are anatomically compartmentalized has made the cornea a useful model for study of stem cell proliferation and differentiation (Chung et al. 1995; Stepp et al. 1995; Zieske et al. 1992, 1994; Cotsarelis et al. 1989; Schermer et al. 1986).
In the cornea, the confinement of the α9 integrin sub-unit to the corneal epithelial limbal basal cells, coupled with studies demonstrating a role for β1 family integrins in epidermal proliferation and differentiation (Hotchkin et al. 1995; Jones et al. 1995; Jones and Watt 1993; Watt et al. 1993), suggested that we determine if conditions that create an increased demand for epithelial cell proliferation and differentiation alter α9 integrin protein and mRNA expression and expression of the α9β1 extracellular matrix ligand tenascin. The experimental model used is a mouse corneal epithelial debridement wound in which small (30–40% of the epithelial surface) or large (∼95% of the epithelial surface) central regions of the corneal epithelium are removed under anesthesia. The manual removal of the epithelium leaves the underlying basement membrane intact.
The biochemical and morphological response of the epithelial cells in the rodent cornea to injury has been studied in detail (Zieske and Gipson 1986; Gipson et al. 1984; Gipson and Kiorpes 1982). Previous studies have shown that closure of smaller types of corneal debridement wounds occurs without cell mitosis. A wave of cell proliferation is initiated after re-epithelialization to permit restratification of the epithelium (Cotsarelis et al. 1989; Zieske et al. 1989; Hanna 1966). For larger wounds, closure involves the rapid induction of cell proliferation of the TA cells near the limbus to provide enough cells to cover the wound site, with proliferation continuing for some time after closure to permit restratification (Chung et al. 1995).
We show here that production of α9 integrin protein and mRNA are enhanced during restratification in smaller epithelial debridement wounds and during both migration and restratification in larger epithelial debridement wounds. We further show that tenascin accumulates beneath the α9-positive epithelial cells during restratification regardless of wound size, but is much more abundant in the larger wounds.
Materials and Methods
In Vivo Wound Healing
All investigations described herein conformed to the regulations of the George Washington University Medical Center Institution Animal Care and Use Committee and are in voluntary compliance with the Statement for the Use of Animals in Ophthalmic and Vision Research established by the Association for Research in Vision and Ophthalmology. Adult male Balb/c mice weighing 18–22 g were anesthetized with 250 μl of a 1:10 dilution of a 1:1 mixture of Ketaset (100 mg/ml) (Aveco; Fort Dodge, IA) and Rompun (20 mg/ml) (Miles; Shawnee Mission, KS). The eyes of anesthetized animals were treated with the topical anesthetic Ophthetic (Allergan America; Hormigueros, PR) several times until the blink response was absent. A 1.5-mm-diameter (smaller wound) or a 3-mm-diameter (larger wound) central corneal area was demarcated with a dulled trephine, and the epithelium within this region was removed by gentle scraping with a dulled scalpel. Immediately after wounding the eyes were treated with erythromycin ophthalmic ointment (E. Fougera; Melville, NY). After 12, 24, and 48 h (for the smaller wound) or 2, 3, 6, and 10 days (for the larger wound), mice were sacrificed by ip injection of 0.5 ml of Fatal-Plus solution containing sodium pentobarbital (Vortech Pharmaceuticals; Dearborn, MI). The corneas were dissected and processed for immunohistochemistry, in situ hybridization, or for in situ cell proliferation as described below.
Preparation of Probes
The murine α9 cDNA clone (209 base pairs), generously provided by D. Sheppard (Lung Biology Center; UCSF, San Fransciso, CA), was linearized with BamH1 and Xho1. Anti-sense and sense probes were transcribed from linearized plasmids using an in vitro RNA transcription kit (Ambion MAXI Script; Austin, TX) in the presence of [35S]-UTP (Du-Pont NEN; Boston, MA). The transcription reaction was carried out with T7 (anti-sense) and T3 (sense) polymerases for 30 min at 37C. Approximately 60–80% of the [35S]-UTP was incorporated, to yield a final specific activity of 1 × 108 cpm/μg RNA.
In Situ Hybridization
In situ hybridization with radiolabeled RNA probes was performed as described previously (Stepp et al. 1993) with the following change made to accommodate frozen tissues. Before pretreatment, silane-coated slides with 6–8 unfixed frozen 6-μm thick sections per slide were dried overnight at 37C and fixed for 5 min in 3% paraformaldehyde in PBS, pH 7.4. Slides were viewed and photographed in brightfield microscopy. No fewer than two corneas from two different animals and two slides containing multiple sections were used for each labeled probe and for each time point studied.
Immunohistochemistry
The polyclonal antiserum against the cytoplasmic domain of α9 integrin was also provided by D. Sheppard. The monoclonal anti-mouse tenascin antibody (T3413) was obtained from Sigma Immunochemicals (St Louis, MO).
Indirect immunofluorescence was performed as previously described (Stepp et al. 1993). Briefly, 6-μm cryostat sections were placed on poly-L-lysine-coated slides and dried at 37C. These slides were rehydrated in PBS and washed in PBS containing 1% (w/v) bovine serum albumin (BSA) (Sigma) and 1% horse serum (Sigma) for 10 min. The primary antibody was then applied for 1 hr at room temperature (RT) in a moist chamber. The slides were rinsed with PBS, followed by 10 min in PBS with 1% BSA and 1% horse serum, and the secondary antibody applied for 1 hr at RT in a moist chamber. After a PBS wash, coverslips were mounted with FITC-guard (Testog; Chicago, IL), and sealed. Negative control tissue sections (primary antibody omitted) were run with each antibody binding assay. The sections were viewed and photographed with an Olympus Vanox ASHBS3 microscope (Opelco; Arlington, VA).
Confocal microscopy was performed in the Center for Microscopy and Image Analysis at GWU Medical Center as previously described (Stepp et al. 1995). Briefly, sections labeled with both fluorescein and Texas red conjugate were sequentially analyzed by a confocal laser scanning microscope (Bio-Rad MRC 1000; Richmond, CA). The typical Z-series is composed of optical sections in the x-y optical plane. Each image was taken at 1-μm intervals with a X 20 objective. Z-series images were projected simultaneously to obtain the final images. Images are presented in pseudo-color based on fluorochrome intensity. Merged images were computer-generated from two optical sections in the same focal plane recorded from the fluorescein or Texas red photomultiplier tubes. The laser color and offset are identical in all panels presented. No fewer than three corneas from three different animals and three slides for each antibody tested were used for each time point studied.
In Situ Cell Proliferation
The in situ cell proliferation kit AP (Boehringer Mannheim; Indianapolis, IN) was used to identify epithelial cells that were engaged in DNA synthesis at the time of sacrifice. Briefly, after in vivo wound healing, dissected corneas were placed in dishes containing media (DMEM; Gibco LTI, Bethesda, MD) supplemented with bromodeoxyuridine (BrdU) labeling solution at a final concentration of 10 μm BrdU for 30 min at 37C in a humidified atmosphere (5% CO2). After incubation, corneas were frozen in tissue embedding medium and 6-μm sections obtained. BrdU was visualized with an anti-BrdU serum conjugated with alkaline phosphatase, as suggested by the manufacturer. After color development, slides were briefly stained with Toluidine blue and were viewed and photographed in brightfield. For quantitation, the number of labeled cells per unit area of the basal membrane surface of the basal cells was determined by morphometry. No fewer than five visual fields were counted per time point. At least two corneas from two different animals and two slides for each time point studied were used for these studies.
Results
Localization and Expression of α9 Protein and mRNA Are Altered in Corneal Epithelium During Restratification After Small (1.5-mm) Debridement
Shown in Figure 1 are data from a representative experiment involving control mice or mice at 12, 24, or 48 hr after small (1.5-mm) debridement. As we have reported previously (Stepp et al. 1995), control mouse corneal epithelium expresses α9 primarily in the basal cells of the limbus (Figures 1A and 1B). At 12 hr after injury there was no detectable α9 in the cells at the leading edge, nor was there a change in the localization of α9 within the limbus (Figures 1C and 1D). In this model, re-epithelialization is complete at 19–22 hr. At 24 hr, there was upregulation of α9 protein in some of the basal cells of the central corneal epithelium (Figures 1E and 1F). By 48 hr after injury, there was less α9 in the basal cells of the central cornea (Figures 1G and 1H). At each time point studied, the distribution of α9 within the limbal basal cells remained unaltered.
To determine whether the patchy localization of α9 protein observed in the central corneal epithelial basal cells at 24 and 48 hr was due to alterations in the steady-state levels of α9 mRNA, in situ hybridization analyses were performed using mouse α9 35S-labeled anti-sense RNA probes. Frozen sections from control mice and mice at 12, 24, or 48 hr after debridement were used in these procedures. The data presented in Figure 2 are brightfield photomicrographs taken of sections after slides were dipped in emulsion, exposed in the dark, developed, and stained with Toluidine blue. In control unwounded mouse cornea, α9 mRNA was detected in both the limbus and the central cornea (Figures 2A and 2B; sense control inset in 2B). In fact, there was no restriction of the α9 mRNA to the limbus, as might be expected from the localization of the protein to the limbus (compare Figures 2A and 2B with Figures 1A and 1B). During migration, there was no apparent change in the localization of α9 mRNA at the leading edge (Figures 2C and 2D). At 24 and 48 hr after injury (Figures 2E-2H), there appeared to be more α9 mRNA expressed in the basal cells of both the limbus and central cornea especially at 48 hr after wounding (compare Figures 2G and 2H, with Figures 2A and 2B).
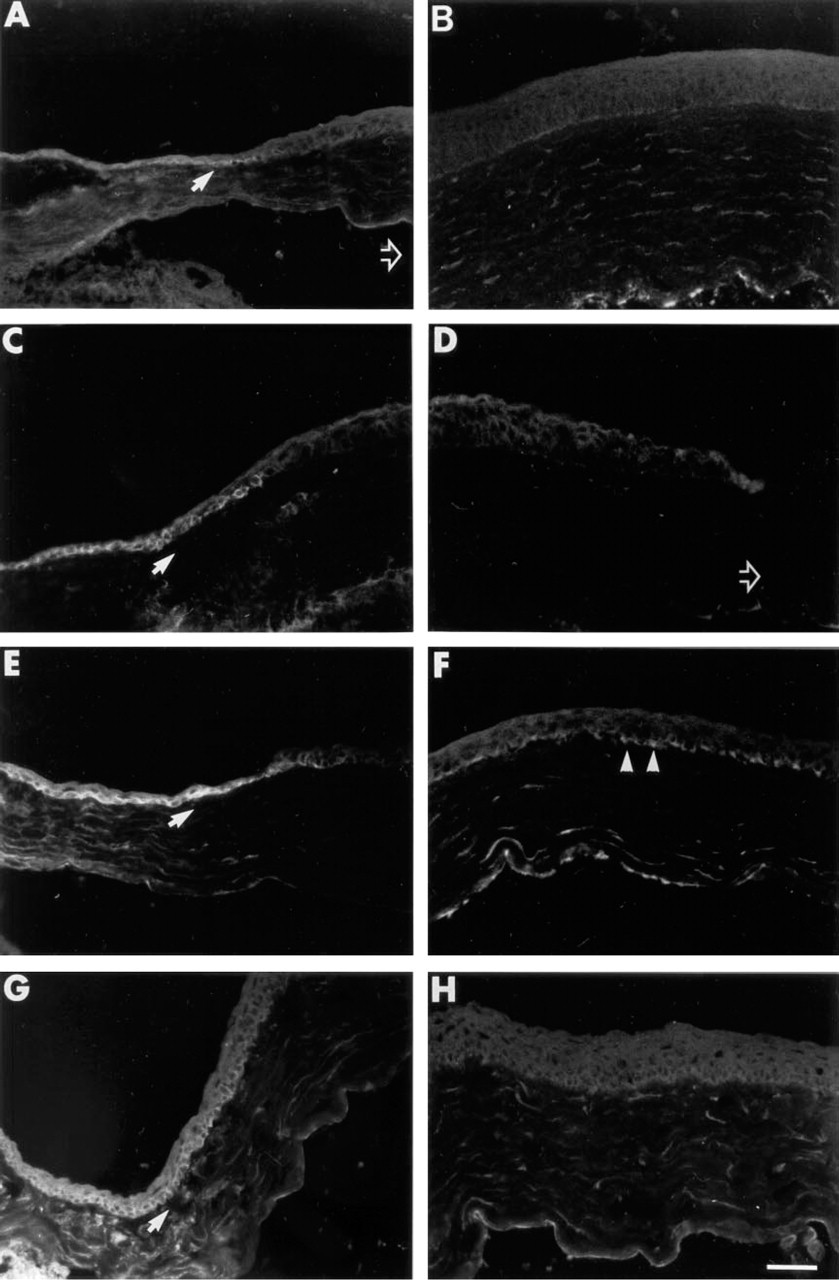
Localization of α9 integrin protein in corneal epithelium after 1.5-mm central debridement.
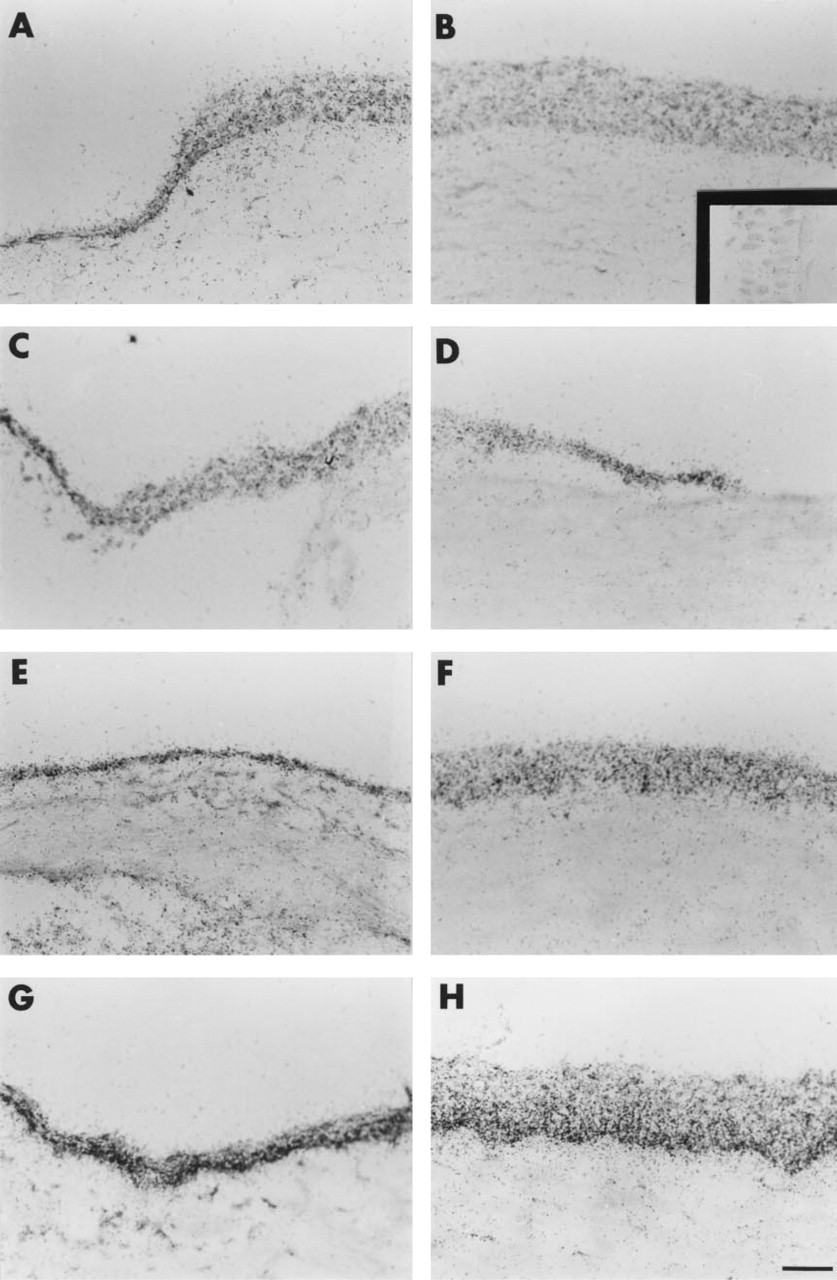
Localization of α9 mRNA in corneal epithelium after 1.5-mm central debridement. Sections were hybridized with 35S-labeled α9 integrin RNA anti-sense probes, slides dipped in photographic emulsion, counterstained with Toluidine blue, and photographed using bright-field microscopy.
The data presented in Figures 1 and 2 show that migration per se does not alter expression and localization of α9 protein and mRNA. However, the events that occur after migration is complete do result in the accumulation of α9 protein within some of the basal cells of the central corneal epithelium and appear to indicate an increase in expression of α9 mRNA in the limbus and in the central corneal epithelium.
To more directly correlate α9 expression with cell proliferation, experiments were conducted using bromodeoxyuridine (BrdU) labeling. BrdU becomes incorporated into the DNA synthesized by cells that are in S-phase. Labeled nuclei are shown in Figures 3A-3D; data are quantitated in Figure 3E. Incorporation of BrdU into DNA is visible within the nuclei of a few scattered epithelial basal cells in control unwounded cornea (Figures 3A and 3E). No change was observed in the numbers of labeled nuclei at 12 hr after 1.5-mm wounding, while the epithelium is actively migrating (Figures 3B and 3E). However, at 24 hr, when migration is complete, a significant increase in the incorporation of BrdU was observed (Figures 3C and 3E). At 48 hr, there were fewer nuclei labeled relative to 24 hr (Figures 3D and 3E). However, there were still more cells labeled than in the control unwounded cornea. These data support the assumption that migration in response to small 1.5-mm debridement wounds occurs in the absence of any increase in the rate of cell proliferation and that as soon as migration is complete, cells begin to proliferate to restratify the epithelium.
Removal of Most of the Corneal Epithelial Cells Results in Dramatic Upregulation of α9 Protein and mRNA in the Epithelial Cells During Migration and Restratification
The data presented above showed that after a 1.5-mm debridement injury to the cornea, α9 integrin protein accumulated in the central corneal epithelial basal cells after epithelial cell migration was complete and during the time when cell division was occurring to re-populate the central corneal to its normal thickness. When larger areas of the corneal epithelium are removed, cell proliferation must occur during migration to generate new cells to cover the exposed wound bed. In the cornea, these cells are generated by the proliferation of a small population of cells located in the basal cell layer of the limbus. This is the same population of cells that express α9 in the unwounded control cornea. To determine whether cells that are required both to migrate and to proliferate alter their localization and expression of α9 integrin, 3-mm debridement wounds were made which involved removal of most of the epithelial surface. Because the Balb/c adult mouse cornea is only slightly larger than 3 mm in diameter, the 3-mm wound removes ∼95% of the corneal epithelial surface; care was taken to avoid injuring the limbus. These debridement wounds took 2.5–3 days to close; they healed without corneal scarring and neovascularization. Figure 4 shows the results obtained when these tissues were analyzed for both α9 integrin protein by indirect immunofluorescence (Figures 4A, 4C, 4E, and 4G) and α9 integrin mRNA by in situ hybridization (Figures 4B, 4D, 4F, and 4H). Compared with control unwounded tissues, the expression of α9 protein was elevated both during migration and during restratification. As a result, there was no longer evidence of differential accumulation of α9 in the basal cells of the limbus, and only the central cornea is shown in Figure 4. Expression of α9 protein in cells at the leading edge at 2 days during active migration was observed (Figure 4A). At 3 days, migration was complete and α9 was detected in all of the cells of the central cornea (Figure 4C). By 6 days, only the basal cells of the central cornea expressed α9 (Figure 4E). At the last time point presented, Day 10, expression of α9 in the cells of the central cornea was almost absent (Figure 4G).
The expression of α9 mRNA was also upregulated at all time points studied. Figure 4B shows expression of α9 mRNA at Day 2. Inset i in Figure 4B shows the sense probe control for this experiment. Inset ii shows the results of anti-sense α9 RNA probe hybridization to control unwounded corneal tissue in this experiment. Inset ii in Figure 4B and Figure 2B show α9 mRNA expression in control tissues. Comparison of these two photomicrographs shows that approximately the same number of silver grains are bound to control tissues in both experiments. The in situ hybridization results presented in Figure 4 were generated by exposure of slides to emulsion for the same length of time as those presented in Figure 2. At all time points evaluated after the larger debridement wounds, there was intense accumulation of silver grains, indicating a dramatic increase in α9 mRNA within these tissues. Interestingly, at 6 and 10 days (Figures 4F and 4H) there is evidence of enhanced accumulation of α9 mRNA in the basal cells. Protein and mRNA expression at earlier time points was also studied. α9 integrin protein and mRNA expression were elevated also at 24 hr, the earliest time point at which epithelial cells could be visualized moving over the corneal surface away from the limbus (data not shown). Therefore, the immunohistochemical and the in situ hybridization data both indicate that a significant increase in expression of α9 integrin occurs in the cells that are migrating over the wound surface and during restratification in response to the 3-mm epithelial debridement wound.
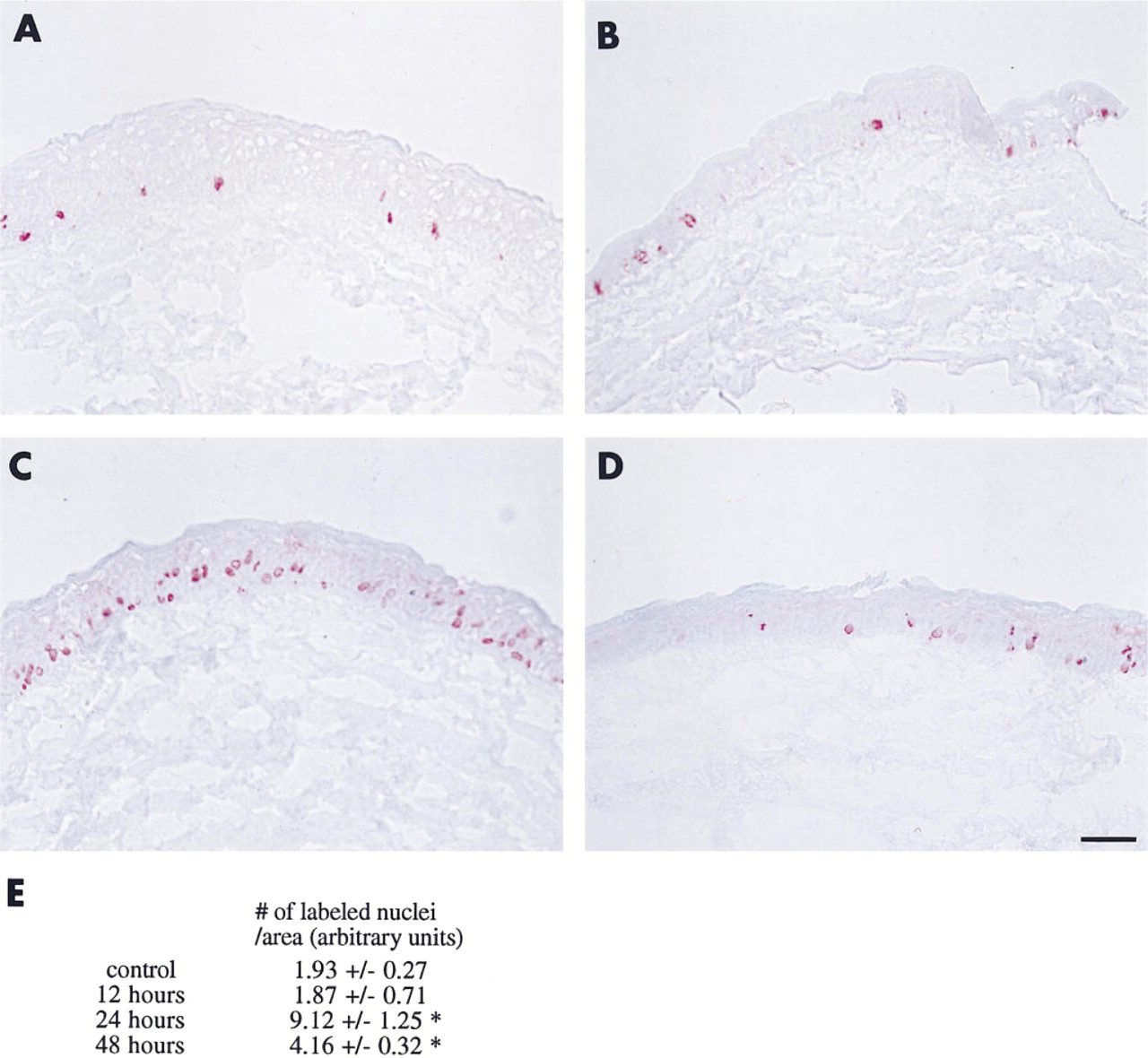
Changes in cell proliferation in response to 1.5-mm central debridement.
Tenascin Accumulates Beneath Epithelial Cells Expressing α9 Integrin
To determine the localization of tenascin, the α9 ligand, relative to that of the α9 protein in the cornea after wounding, double labeling immunofluorescence microscopy was performed on corneal tissue sections and viewed by confocal microscopy. The results are presented in Figure 5. In control tissues (Figure 5A), note the presence of tenascin in the stroma underlying the region anterior to the α9-positive limbal basal cells in the control cornea. The central region of the cornea is negative for both α9 protein and tenascin in the epithelium, at the basement membrane junction, and in the stroma (Figure 5A, inset). At 24 hr after the smaller (1.5-mm) wounds, a few α9-positive epithelial basal cells overlie patchy regions of tenascin-positive matrix. The α9-positive epithelial cells themselves are negative for tenascin protein (Figure 5B). Tenascin also begins to accumulate in the posterior stroma in the smaller wounds at 24 hr. In the larger (3-mm) wounds, α9 expression in the epithelial cells is not correlated with expression of tenascin by the epithelial cells or at the basement membrane zone during migration at Day 2. There is, however, accumulation of tenascin in the stroma (Figure 5C). By 3 days in the larger wounds, patchy areas of tenascin-positive extracellular matrix appear beneath the epithelial cells (Figure 5D) and by 6 days an intense band of tenascin-positive extracellular matrix material has accumulated beneath the epithelial cells (Figure 5E). At later time points, the expression of both α9 integrin in the epithelial basal cells and tenascin at the basement membrane zone beneath the epithelial cells begins to decrease, as shown in a representative micrograph from a cornea 10 days after injury (Figure 5F).
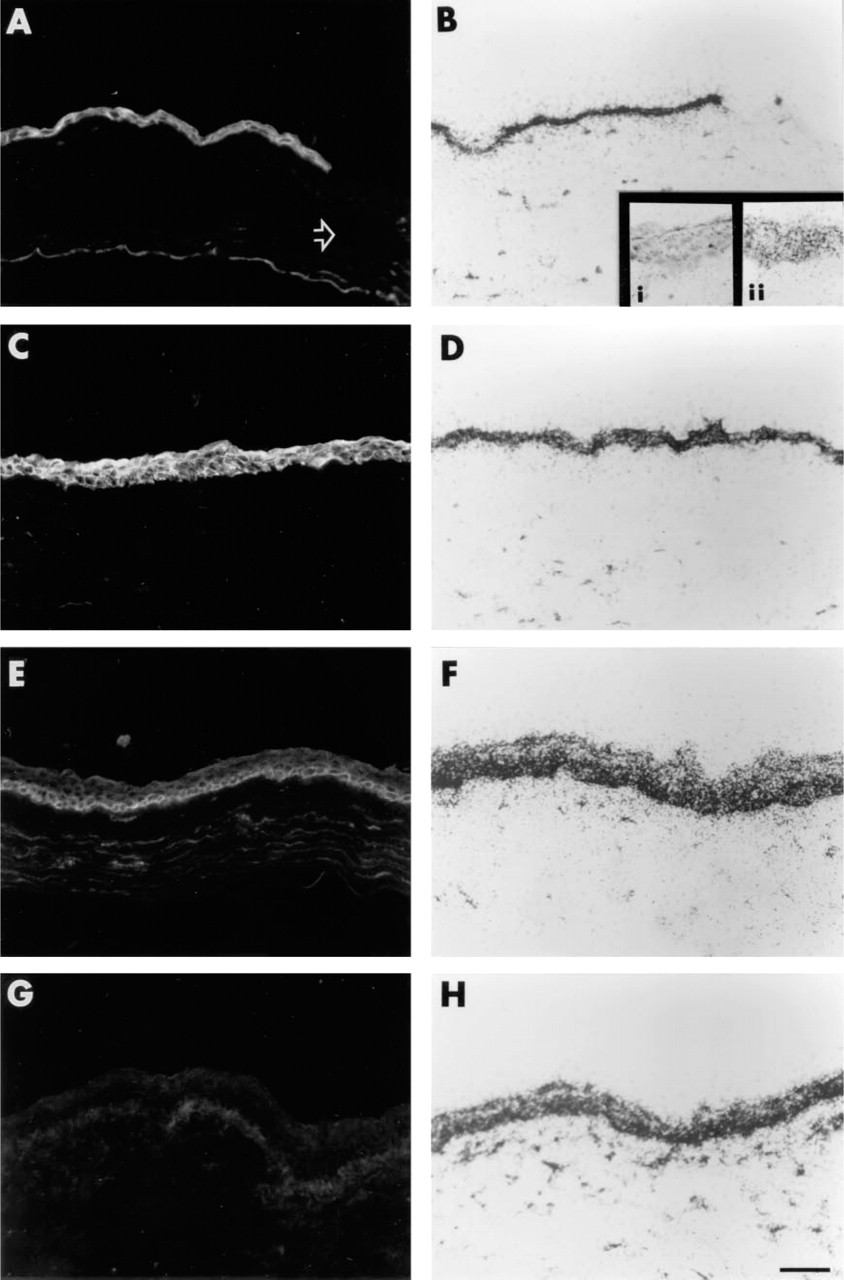
Localization of α9 integrin protein and mRNA in corneal epithelium after 3-mm debridement.
Discussion
The data presented above show that α9 integrin protein and mRNA expression are enhanced when cell proliferation is induced by debridement. The in situ hybridization studies indicate that α9 integrin mRNA levels increase along with protein levels during cell proliferation in response to injury. This suggests that the changes observed in α9 integrin production are regulated at the level of transcription. Studies are under way to quantitate mRNA levels, using RNAse protection analyses, to enable us to quantify the steady state expression of integrin mRNAs in the cornea in response to injury. It is also interesting that α9 mRNA can be observed in cells that do not express detectable levels of the α9 protein. This is easily observed in the central region of the normal cornea and in the failure of the mRNA to be restricted to the basal cells of the limbus (Figures 2A and 2B). We lack sufficient data to enable us to determine whether there is α9 protein expressed and rapidly degraded in these cells or whether the mRNA is being stored and not used for translation of protein. In the normal cornea, therefore, α9 protein accumulation is regulated at both transcriptional and non-transcriptional levels; in the injured cornea during cell proliferation, however, there appears to be increased expression of α9 mRNA suggesting transcriptional regulation.
By comparing the integrin profile after smaller wounds, which close without the requirement for cell mitosis, vs larger wounds whose closure requires both mitosis and migration, we have shown that α9 protein and mRNA expression are induced coordinately with cell proliferation. In addition, tenascin accumulated under the corneal epithelial cells expressing α9 integrin during restratification, and was especially apparent 6 days after the larger wound. This intense localization of tenascin observed after the larger debridement appears to correlate with active remodeling of the basement membrane and re-insertion of hemidesmosomes, as suggested by studies showing co-distribution of tenascin with p4 integrin at this time point (data not shown). α9 integrin expression, cell proliferation, and tenascin accumulation at the epithelial basement membrane are correlated.
α9 integrin is one of the most recently characterized of the integrin molecules (Palmer et al. 1993). Studies have shown that expression of α9 is regulated developmentally in the mouse (Wang et al. 1995). In the differentiating mouse corneal epithelium, α9 is expressed transiently (Stepp et al. 1995). It is initially absent early in corneal development, when the corneal epithelium is only two cell layers thick. α9 protein expression is upregulated in the developing cornea at the same time that the corneal epithelium undergoes a period of rapid cell proliferation and becomes fully stratified. Therefore, in the corneal epithelium, expression of α9 integrin is regulated both developmentally and during proliferation in response to injury.
α9 integrin forms heterodimers with the β1 integrin subunit and in adults localizes to the basal cells of the epidermis, skeletal muscle, hepatocytes, to the airway epithelium lining the lungs, and to the basal cells of the corneal limbus. The only well-characterized ligand for α9β1 integrin is tenascin (Yokosaki et al. 1994). The expression of α9β1 in tissues that lack tenascin, as well as the failure of α9 to co-localize with tenascin in some cells that express both proteins (Wang et al. 1995), suggests that additional ligands will probably be revealed as research on α9 integrin advances. Because α9β1 is present on lateral, apical, and basal membranes of epidermal cells, it cannot be ruled out that α9β1 functions in cell-cell adhesion. α2β1 and α3β1 are two of the most abundant integrins expressed in epithelial tissues and have been suggested to play a role in cell-cell adhesion, possibly via homophilic integrin-integrin interaction (Jensen and Wheelock 1995; Symington, et al. 1993; Tenchini et al. 1993; Carter et al. 1990; Larjava et al. 1990).
Tenascin is a large extracellular matrix protein believed to play a role in epithelial-mesenchymal interactions both during development and in adults (Steindler et al. 1995; Lightner 1994; Erickson 1993). In the developing avian eye, tenascin expression is correlated with important cell migration events in the corneal endothelium and stroma (Tucker 1991; Weller et al. 1991). The tenascin molecule is a hexamer, and its structure is arranged in a series of globular repeats. Its role in mediating cell-matrix interactions depends on cellular context. In some cell types, tenascin can block or interfere with cell adhesion to fibronectin via steric blocking and by interfering with focal contact formation (Lightner 1994). In other cells, tenascin can support cell adhesion via integrin-dependent mechanisms. Both α9β1 and another β1 family integrin heterodimer, α8β1 (Schnapp et al. 1995), have recently been shown to interact with the third fibronectin Type III repeat of tenascin. This region of tenascin is not subject to alternative splicing and is present in all of the alternatively spliced forms of tenascin. In addition to α8β1 and α9β1, αvβ6 is believed to interact with tenascin (Prieto et al. 1993).
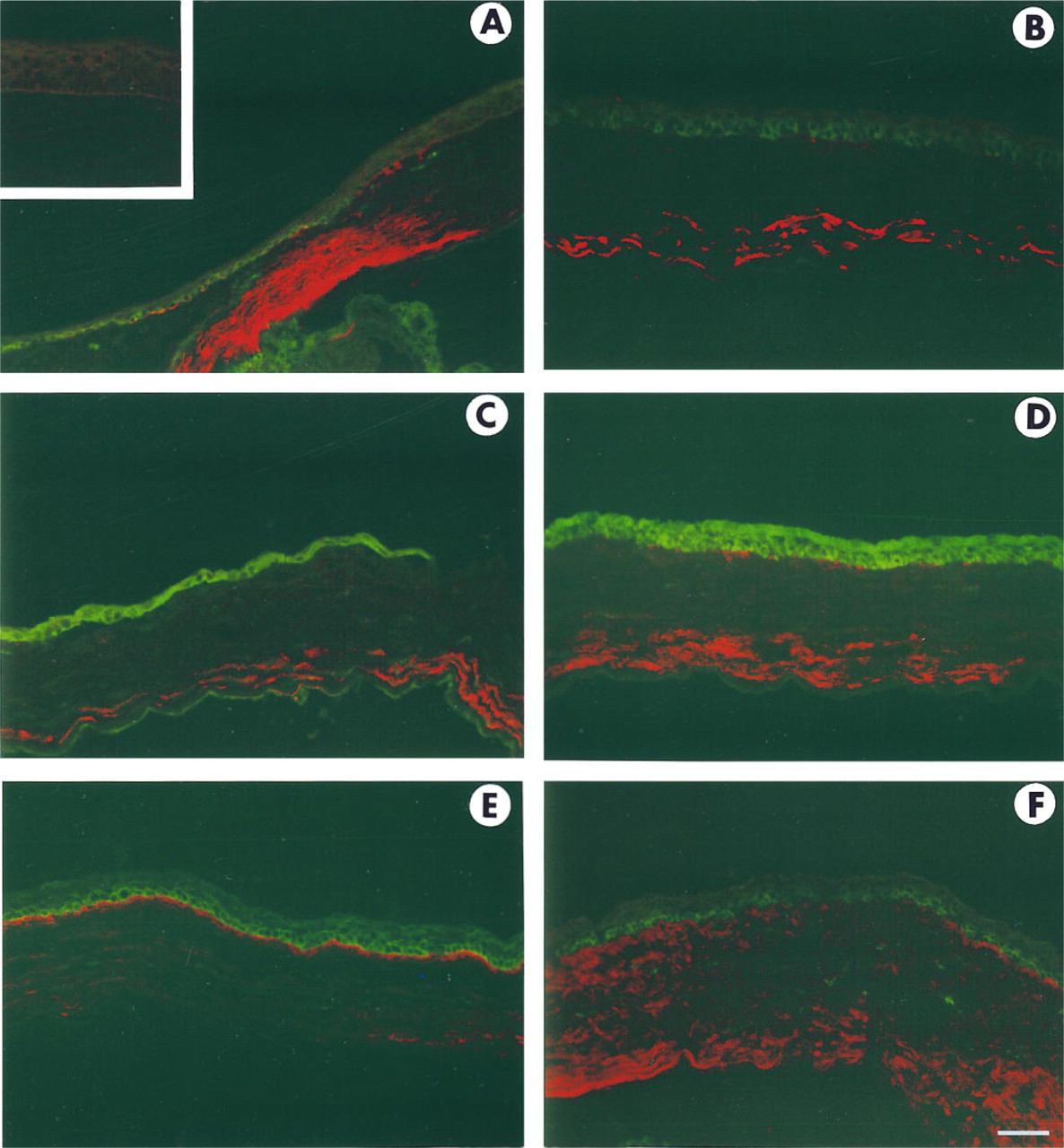
Co-localization of α9 integrin protein (green) and tenascin (red) after epithelial debridement. Confocal microscopy was used to simultaneously visualize α9 integrin and tenascin on tissue sections.
A role for tenascin in wound healing in the skin and in cornea has been suggested by the dramatic increase in tenascin in the healing dermis (Betz et al. 1993; Mackie et al. 1988) and in the stroma in the healing cornea (Figures 5B-5F) (van Setten et al. 1992; Tervo et al. 1991). In normal epidermis, low levels of tenascin are localized beneath the basal surface of the basal cells within the basement membrane. In situ hybridization studies of the epidermis of newts (Onda et al. 1991) and of skin wound healing in rats (Aukhil et al. 1996) indicate that the epithelial cells themselves produce the tenascin that is found associated with the basement membrane. Tenascin is not present in the epithelial basement membrane of the normal unwounded cornea (Latvala et al. 1995; van Setten et al. 1992; Tervo et al. 1991). In situ hybridization analyses using tenascin RNA probes are under way to resolve whether the accumulation of tenascin beneath epithelial cells we observe in Figure 5 during restratification is due to production of the molecule by epithelial cells or by stromal fibroblasts. Figure 5 also demonstrates expression of tenascin in the stroma itself in response to both the small and large wounds. Expression is first observed in the posterior stroma and then spreads to involve the entire stroma. These data are consistent with those from similar studies conducted in the corneas of rabbits (Latvala et al. 1995). The work of Chiquet-Ehrismann et al. (1994), studying tenascin promoter function in fibroblasts in culture that were subjected to stress or tension, suggests a possible mechanism for the increase in tenascin expression observed in response to injury i.e., the swelling of the corneal stroma that occurs after the loss of the epithelial barrier could induce tenascin expression by placing the fibroblasts under increased tension, which would then activate the tenascin promoter. The poor preservation of the corneal stroma that occurs in unfixed frozen sections does not enable us to make a more detailed analysis of integrin and tenascin expression in the corneal stroma.
Several studies have shown that production of specific integrin α and β chains appears to be increased in the skin and cornea after injury (Stepp et al. 1996; Gailit et al. 1994; Cavani et al. 1993; Juhasz et al. 1993; Larjava et al. 1993; Hertle et al. 1992; Guo et al. 1991; Clark 1990). Studies in nude mice of the healing of wounded human skin grafts have shown that β1 and αv are increased at the leading edge of migrating epithelial sheets (Juhasz et al. 1993). A recent report shows that αvβ6 is upregulated in human epidermal wound healing (Haapasalmi et al. 1996).
In the cornea, we recently showed that the hemidesmosomal integrin α6β4 was upregulated in the epithelial cell early (3–8 hr after injury) during migration in response to debridement wounding (Stepp et al. 1996). These β4 experiments were conducted in both rats and mice, using the smaller wounds in which re-epithelialization occurs without a requirement for cell proliferation. Because the production of α9 is not altered early in response to small injuries but later during the re-stratification stages of wound healing, the corneal epithelial cells clearly regulate β4 and α9 integrins via distinct mechanisms. In our previous studies we have been unable to document any significant changes in the expression of β1 or αv integrin in the cornea during wound healing (Stepp et al. 1994, 1996). Although α9 forms heterodimers with β1, much more β1 is present in the epithelial cells forming heterodimers with α2 and α3. Therefore, it is not surprising that the modest increase in expression of α9 we observed in these studies in the smaller wounds at 24 and 48 hr (Figures 1E-1H) occurred in the absence of changes in the abundant β1 integrins. The increased expression of αvβ6 in skin during healing was demonstrated using a complex specific monoclonal antibody against human αvβ6 (Haapasalmi et al. 1996) which does not react with rodent tissues. We have evaluated the αv subunit biochemically in the corneal epithelium but did not look at expression of β5 and β6.
Evidence suggests that integrin expression and function in the epidermis are involved in mediating epithelial cell proliferation and differentiation (Hotchkin et al. 1995; Jones and Watt 1993; Watt et al. 1993). Jones et al. (1995) have succeeded in isolating stem cell-rich populations of cells from human epidermis by selecting cells on the basis of their enhanced expression of the β1 family integrins, α2β1 and α3β1. Cells expressing high levels of these two integrin heterodimers have the proliferative properties expected of epidermal stem cells, with cells expressing low levels having low proliferative potential. The role of α9 in epidermal proliferation has yet to be evaluated.
Although progress has been achieved that implicates integrins in cell proliferation and migration, less progress has been made in the analysis of the extracellular ligands for integrins. The relative lack of progress in this area is due primarily to the large number of distinct extracellular matrix proteins present in the epithelial basement membrane. Several different laminin isoforms are present whose molecular identity has only recently been solved (Burgeson et al. 1994; Marinkovich et al. 1992a,b). In addition, a variety of Type IV collagen genes and proteins are found in the corneal stroma and basement membrane (Ljubimov et al. 1995). Fibronectin and tenascin are present, but their expression is restricted and transient (Latvala et al. 1995). The complexity of the matrix is mirrored in the additional complexity of the integrins in terms of their ligand binding properties. Most integrin αβ heterodimers have been found to bind to more than a single extracellular matrix molecule. For example, α2β1 can mediate attachment both to various collagen proteins and to many laminin isoforms. The variety of matrix molecules present, coupled with the ability of many integrins to interact with multiple distinct matrix proteins, has resulted in slower progress in the identification of the extracellular matrix molecules involved in integrin-mediated cell attachment during wound healing.
Understanding of epithelial cell-cell and cell-substrate adhesion at the molecular level is just beginning to be achieved. Data point to integrins as mediators of epithelial adhesion, migration, proliferation, and differentiation. Research on epithelial cell proliferation and migration in response to injury has clinical relevance for improving our understanding of the basis of treatment for patients with defects in healing. In ophthalmology, for example, recurrent epithelial erosions and related corneal dystrophies are significant corneal diseases. Minor trauma causes the debridement of the epithelium in these patients. A popular treatment for this condition involves debriding the epithelial tissue around the site of the erosion, followed by bandaging the eye (Wood and Griffith 1988; Buxton and Constad 1987). Debridement of the corneal epithelium is routinely performed before eximer laser treatment of patients with myopic corneas and also during retinal surgery, because the epithelium becomes cloudy and interferes with the surgeon's view of the retina. An improved understanding of epithelial cell's proliferative and migratory responses to this type of injury will both aid clinicians and help us to better understand the basic mechanisms that control these events.
Taken together, our results and those of others studying skin and corneal wound healing point to roles for specific integrin αβ heterodimers in the proliferation, migration, and differentiation of epithelial cells. α9 integrin is abundant in the corneal limbus in cells that are believed to be the corneal epithelial stem cells. In response to corneal debridement, we have shown here that enhanced α9 integrin expression is correlated with induction of cell proliferation. We have also shown that accumulation of the α9β1 ligand tenascin is altered during epithelial cell proliferation. Further studies must be performed to determine whether these changes are essential for appropriate healing of epithelia.
Footnotes
Acknowledgements
Supported by NIH grant EYO8512–08 (MAS).
We thank James Kendrick and the photography staff in Biomedical Communications at the George Washington University Medical Center for help with photography, Fred Lightfoot in the Center for Microscopy and Image Analysis for help with the confocal microscopy, and Dr Abdo Romano Jurjus, Visiting Associate Professor in the Department of Anatomy and Cell Biology at the George Washington University Medical Center and Associate Professor, Department of Human Morphology, Faculty of Medicine, American University of Beirut, Lebanon, for comments on the manuscript.