
Abstract
Recent studies have revealed that phosphoinositide (PI) signaling molecules are expressed in mammalian retinas, suggesting their importance in its signal transduction. We previously showed that diacylglycerol kinase (DGK) isozymes are expressed in distinct patterns in rat retina at the mRNA level. However, little is known about the nature and morphological aspects of DGKs in the retina. For this study, we performed immunohistochemical analyses to investigate in the retina the expression and localization of DGK isozymes at the protein level. Here, we show that both DGKβ and DGKι localize in the outer plexiform layer, within which photoreceptor cells make contact with bipolar and horizontal cells. These isozymes exhibit distinct subcellular localization patterns: DGKι localizes to the synaptic area of bipolar cells in a punctate manner, whereas DGKβ distributes diffusely in the subsynaptic and dendritic regions of bipolar and horizontal cells. However, punctate labeling for DGKϵ is evident in the outer limiting membrane. DGKζ and DGKα localize predominantly to the nucleus of ganglion cells. These findings show distinct expression and localization of DGK isozymes in the retina, suggesting a different role of each isozyme.
Keywords
Following activation of Gq protein–coupled receptors in response to external stimuli, phospholipase C (PLC) yields a pair of second messengers, diacylglycerol (DG) and inositol 1,4,5-trisphosphate, in phosphoinositide (PI) turnover. In this system, diacylglycerol kinase (DGK) phosphorylates DG to produce another second messenger, phosphatidic acid (PA). A well-known functional role of DGK is regulation of protein kinase C (PKC), for which DG acts as an allosteric activator, and the activity of which plays a central role in many different cell types. In addition, recent studies have revealed that PA acts as a messenger to regulate many signaling molecules. Therefore, DGK is thought to mediate signal transduction by modulating levels of DG and PA (i.e., the attenuation of DG and the production of PA). To date, 10 mammalian DGK isozymes have been identified (Topham 2006; Goto et al. 2007; Sakane et al. 2007; Martelli et al. 2011). A salient feature of the DGK family is that most isozymes are abundantly expressed in the brain, suggesting the physiological importance of this enzyme family for proper brain function (Goto and Kondo 2004; Goto et al. 2006).
Photoreceptors contain highly specialized structures for phototransduction, which is mediated by rhodopsins and heterotrimeric G-proteins. Signal transduction in the retina is initiated by the absorption of light by photopigments in the outer segment of the photoreceptor cells. Transduction of the image by photoreceptor cells is transmitted to bipolar cells by excitatory chemical synapses (Sterling and Demb 2004). The photosignal is conveyed via bipolar cells to ganglion cells, the axons of which extend to the lateral geniculate nucleus through the optic nerve and tract. Each synaptic stage has specialized, laterally connecting neurons called horizontal and amacrine cells. These neurons modify (largely by inhibitory chemical synapses) forward transmission across the synaptic layers (Sterling and Demb 2004). Phototransduction in mammalian retinal photoreceptor cells involves a cyclic guanosine monophosphate (cGMP) signaling cascade that controls cGMP-gated cation channels, through which photoreceptors release glutamate continuously in the dark or attenuate its release when stimulated by light (Arshavsky et al. 2002).
In contrast, phototransduction in flies is operated by PI signaling (Zuker 1996; Hardie 2003). Although the signal cascades in photoreceptor cells of vertebrates and flies are initiated by both activation of related rhodopsins and engagement of heterotrimeric G-proteins, notable differences exist: Light stimulation of rhodopsin and cone opsins in mammalian photoreceptors increases the activity of a cGMP phosphodiesterase and closure of the cGMP-gated channels, whereas the cascade in fly photoreceptors is operated through activation of PLCβ and opening of light-sensitive channels (transient receptor potential and transient receptor potential-like) (Zuker 1996; Hardie 2003; Wang and Montell 2007). In addition, the importance of PI signaling in fly retina is suggested by findings related to various Drosophila mutants of the PI cascade components (Hardie 2003; Wang and Montell 2007). For example, a defect in the Drosophila-PLC enzyme results in a mutant called norpA, the photoreceptors of which fail to show any electrical response to light (Hotta and Benzer 1970; Bloomquist et al. 1988). Furthermore, Drosophila retinal degeneration A (rdgA) mutant, which lacks eye-specific DGK activity, has photoreceptor cells that differentiate normally but degenerate rapidly after eclosion (Hotta and Benzer 1970; Masai et al. 1993).
Recent studies have revealed that PI signaling molecules, which include the group I metabotropic glutamate receptors (mGluRs), Gq subclass of the G-protein, PLCβ subfamily, DGKs, and conventional PKCs, are expressed at the mRNA or protein level in mammalian retina (Philp et al. 1987; Fujisawa et al. 1992; Jiang et al. 1996; Kohyama-Koganeya et al. 1997; Koulen et al. 1997; Peng et al. 1997; Lee et al. 2007; Whitaker and Cooper 2009; Sato et al. 2011). Based on these data, it can be readily inferred that PI signaling is involved in signal transduction in mammalian retinas, although the detailed functional aspects related to this question remain poorly understood. In this regard, we showed that some DGK isozymes are expressed in distinct patterns in rat retina at the mRNA level (Sato et al. 2011). To date, we have raised specific antibodies against DGKs and examined unique expression patterns of respective isozymes in rat brain under pathophysiological conditions (Hozumi et al. 2003; Goto et al. 2007; Nakano, Iseki, et al. 2009; Saino-Saito et al. 2011; Hozumi and Goto 2012; Suzuki et al. 2012). However, little is known about the nature and morphological aspects of DGKs in the retina, which is also of neuroectodermal origin. In this study, we investigated in the retina the expression and localization of DGK isozymes at the protein level using immunohistochemistry. Here, we show that DGK isozymes display distinct cellular expression and subcellular localization patterns. These findings suggest a unique role assigned to each isozyme in retinal signal transduction.
Materials and Methods
Animals
Adult male Wistar rats (Japan SLC) were used in the present study. The rats were housed under freely ranging circumstances in our animal facilities under normal lighting conditions (lights on 08:00-20:00) and were allowed ad libitum access to normal rodent chow and water. The eyes were dissected under light conditions (12:00-15:00). All animals were treated according to the guidelines for the care and use of laboratory animals of the Yamagata University School of Medicine.
Tissue and Section Preparation
For immunohistochemistry, rats anesthetized with ether were fixed transcardially with 4% paraformaldehyde in 0.1 M sodium phosphate buffer (PB, pH 7.2) or periodate lysine paraformaldehyde (PLP) for immunofluorescence microscopy (Hozumi et al. 2003; Hozumi et al. 2008; Hozumi et al. 2010). After fixation, eyeballs were cryoprotected in a 30% sucrose/0.1 M PB and cut into 16-µm-thick sections using a cryostat (CM1900; Leica, Nussloch, Germany).
Antibody Production
The cDNA fragment encoding the N-terminal region of DGKι (amino acid residues 99–132) was obtained by polymerase chain reaction using rat DGKι cDNA (Ito et al. 2004). The cDNA fragment, flanked by BamHI and EcoRI sites, was subcloned into the pGEX-4T-1 vector (GE Healthcare; Piscataway, NJ) for expression of glutathione S-transferase (GST) fusion protein and was sequenced to confirm the reading frame. GST fusion proteins were expressed and purified using glutathione–Sepharose 4B (GE Healthcare), according to the manufacturer’s instructions. GST fusion proteins were emulsified with Freund’s complete or incomplete adjuvant (DIFCO; Detroit, MI) and immunized subcutaneously to guinea pigs at intervals of 2 weeks. After the fifth injection, polyclonal antibodies specific to DGKι were affinity-purified using GST-free peptide coupled to CNBr-activated Sepharose 4B (GE Healthcare). GST-free peptides were prepared by in-column thrombin digestion of GST fusion proteins bound to glutathione–Sepharose 4B media. The specificity of the guinea pig anti–DGKι antibody is shown in Suppl. Fig. S1.
Immunohistochemistry
A brief section pretreatment with methanol greatly enhanced immunoreactivities for PI signaling molecules (Nakamura et al. 2004; Nomura et al. 2007; Fukaya et al. 2008; Hozumi et al. 2008). Therefore, sections for immunofluorescence were treated, prior to normal serum blocking, with dipping in 100% methanol for 15 min.
All immunohistochemical incubations were performed at room temperature. In double immunofluorescence, guinea pig anti–DGKβ antibody (1 µg/ml) (Hozumi et al. 2008), rabbit anti–DGKζ antibody (1 µg/ml) (Hozumi et al. 2003), guinea pig anti–DGKζ antibody (1 µg/ml) (Okada et al. 2011), rabbit anti–DGKι antibody (1 µg/ml) (Ito et al. 2004), or guinea pig anti–DGKι antibody (1 µg/ml) diluted with PBS containing 0.1% Triton X-100 was incubated overnight in a mixture with one of the following antibodies: rabbit anti–DGKι (1 µg/ml), rabbit anti–protein kinase Cα (PKCα; 1 µg/ml; Frontier Institute Co. Ltd, Hokkaido, Japan), guinea pig anti–protein kinase Cα (PKCα; 1 µg/ml; Frontier Institute Co. Ltd), guinea pig anti–vesicular glutamate transporter 1 (VGluT1; 1 µg/ml) (Miyazaki et al. 2003), goat anti–Brn-3a (1 µg/ml; sc-31984; Santa Cruz Biotechnology, Santa Cruz, CA), mouse anti-calretinin (1 µg/ml; MAB1568; Chemicon, Southampton, UK), or rabbit anti–calbindin D-28K (1:500; AB1778; Chemicon). They were visualized by 2-hr incubation with species-specific secondary antibodies at a dilution of 1:200 (Molecular Probes; Eugene, OR). In double immunofluorescence using anti–DGKα antibody (Goto et al. 1992), immunofluorescence was performed using the TSA Biotin System (NEL700A; NEN, Boston, MA) according to the manufacturer’s instructions. First, sections were sequentially incubated with TNB blocking buffer (included in the kit) for 30 min, rabbit DGKα antibody (1 µg/ml) overnight, biotinylated secondary antibody for 30 min, streptavidin-peroxidase complex for 30 min, and biotin tyramide solution (included in the kit) for 10 min, followed by visualization using streptavidin-conjugated Alexa 488 (Molecular Probes) in PBS for 30 min. After blocking with 5% normal goat serum, conventional immunofluorescence was performed using mouse anti–glutamine synthetase (1 µg/ml; 610517; BD Transduction Laboratories, San Jose, CA) or goat anti–Brn-3a (1 µg/ml; sc-31984; Santa Cruz Biotechnology) antibody as above.
For single immunofluorescence, antibodies used are as follows: guinea pig anti-DGKβ (1 µg/ml), rabbit anti-DGKζ (1 µg/ml), rabbit anti-DGKι (1 µg/ml), rabbit anti-DGKϵ (1 µg/ml) (Nakano, Hozumi, et al. 2009), rabbit anti-Gαq/11 (1 µg/ml; sc-392; Santa Cruz Biotechnology) (Tanaka et al. 2000), rabbit anti–type 1α metabotropic glutamate receptor (mGluR1α; 1 µg/ml) (Tanaka et al. 2000), rabbit anti–type 5 metabotropic glutamate receptor (mGluR5; 1 µg/ml) (Uchigashima et al. 2007), mouse anti–phospholipase Cβ1 (PLCβ1; 1 µg/ml; sc-5291; Santa Cruz Biotechnology), goat anti–phospholipase Cβ2 (PLCβ2; 1 µg/ml; sc-31757; Santa Cruz Biotechnology), rabbit anti–phospholipase Cβ3 (PLCβ3; 1 µg/ml) (Nomura et al. 2007), guinea pig anti–phospholipase Cβ4 (PLCβ4; 1 µg/ml) (Nakamura et al. 2004), rabbit anti–protein kinase CβII (PKCβII; 1 µg/ml) (Hasegawa et al. 2008), rabbit anti–protein kinase Cγ (PKCγ; 1 µg/ml) (Yoshida et al. 2006), or rabbit anti–diacylglycerol lipase-α (DGLα; 1 µg/ml) (Yoshida et al. 2006) antibody. After thorough washing in PBS, they were visualized by 2-hr incubation with species-specific secondary antibodies at a dilution of 1:200 (Molecular Probes). The specificities of the antibodies used in the current study are shown in Suppl. Figs. S1 and S2 and Suppl. Table S1.
Sections were stained for the nuclei of cells using TO-PRO-3 (1:200; Molecular Probes) and scanned using a confocal laser scanning microscope (LSM510META; Carl Zeiss, Göttingen, Germany) in multitrack mode. In addition, the light microscopic differential interference contrast (DIC) images were taken together with the immunofluorescence images.
Results
Expression and Localization of DGK Isozymes
In our previous study, Northern blot analysis showed that in adult rat retina, hybridization signals were detected intensely for DGKι and DGKϵ, moderately for DGKβ and DGKζ, and weakly for DGKα. DGKγ mRNA was below detection levels (Sato et al. 2011). In this study, we examined cellular expression and localization of DGK isozymes in the retina at the protein level by immunofluorescence and counterstained simultaneously with TO-PRO-3 for the nuclei.
DGKβ
DGKβ immunoreactivity was found intensely in the outer plexiform layer (OPL) and moderately in the inner nuclear layer (INL) and inner plexiform layer (IPL) (Fig. 1A). To examine which cell type(s) are responsible for the expression, we next performed double immunolabeling experiments using antibodies against calbindin D-28K, a marker for horizontal cells (Zheng et al. 2012), and PKCα, a marker for rod bipolar cells (Whitaker and Cooper 2009). DGKβ was coexpressed in cells that were immunoreactive for calbindin D-28K (arrowheads, Fig. 1B). In addition, most DGKβ immunoreactivity was detected in PKCα-positive cells (thick arrows), but occasionally it was recognized in PKCα-negative perikarya (thin arrows) in the INL (Fig. 1C). These findings suggest that DGKβ localizes in rod and cone bipolar cells together with horizontal cells.
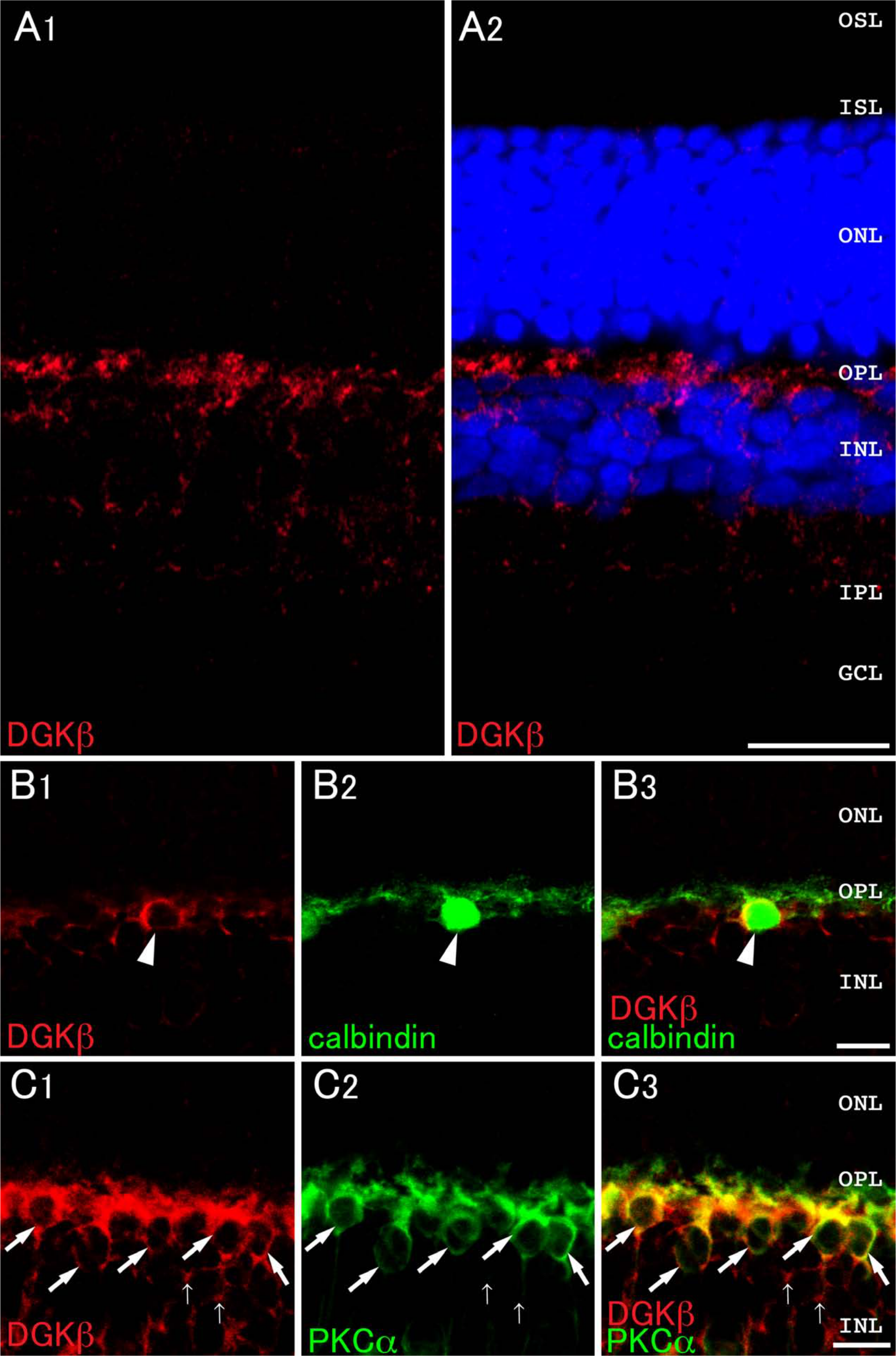
Expression and localization of DGKβ in the retina. Double immunofluorescence for characterization of DGKβ (red) and calbindin D-28K (calbindin, green, B) or PKCα (green, C) in the retina. Blue fluorescence represents nuclear staining with TO-PRO-3. (A) DGKβ is intensely detected in the OPL. At higher magnification, DGKβ immunoreactivity is detected in the perikarya and dendritic regions of calbindin-positive horizontal cells (arrowheads, B) and PKCα-positive rod bipolar cells (thick arrows, C) in the IPL. DGKβ immunoreactivity is also found in putative cone bipolar cells that are not stained by PKCα (thin arrows, C). GCL, ganglion cell layer; IPL, inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer; ISL, inner segment layer; OSL, outer segment layer. Scale bars = 20 µm (A); 10 µm (B and C).
DGKι
DGKι-immunoreactive puncta were distributed mainly in the OPL and sparsely in the INL (Fig. 2A). In double immunostaining, these DGKι-positive puncta were shown to be apposed side-by-side and never overlapped with immunoreactivity for vesicular glutamate transporter 1 (VGluT1), a marker for ribbon synapses of photoreceptor terminals in the OPL (Whitaker and Cooper 2009) (Fig. 2A,B). This suggesting that DGKι localizes to the postsynaptic region juxtaposed to the photoreceptor synapses. DGKι-positive puncta were detected intensely in dendrites of PKCα-positive cells (rod bipolar cells) (Fig. 2C), although the signals for DGKι were not overlapped with immunoreactivity for calbindin D-28K (horizontal cells) (Fig. 2D). These results suggest that DGKι mainly localizes at the postsynaptic region of rod bipolar dendrites but not of horizontal cells.
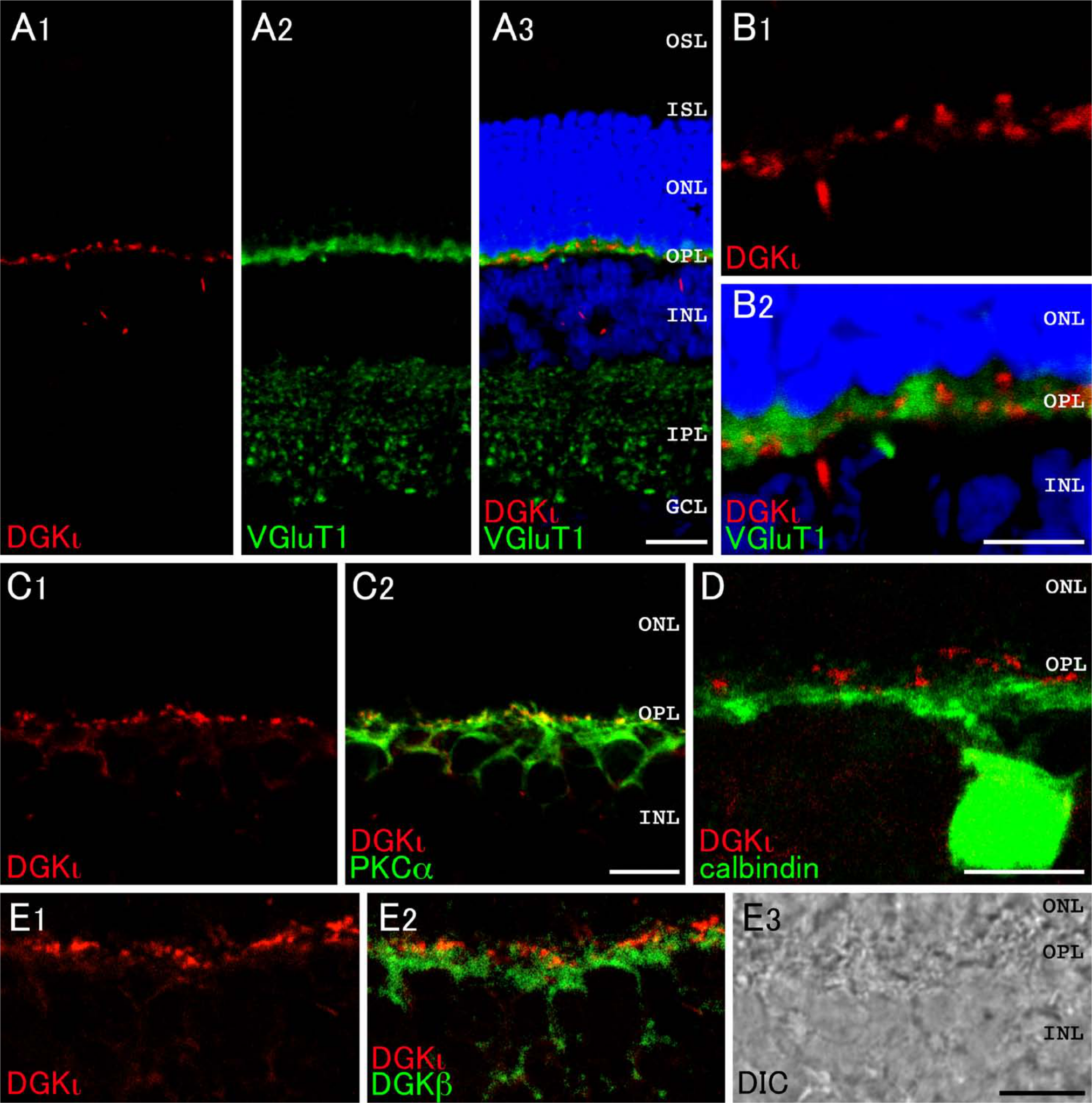
Expression and localization of DGKι in the retina. Double immunofluorescence for characterization of DGKι (red) in the retina. Green fluorescence represents vesicular glutamate transporter 1 (VGluT1, A and B), PKCα (C), calbindin D-28K (calbindin, D), or DGKβ (E). Blue fluorescence represents nuclear staining with TO-PRO-3. (A, B) Expression and localization for DGKι and VGluT1 at low (A) and high magnification (B). DGKι-immunoreactive puncta are distributed mainly in the OPL. Note that puncta labeled for DGKι and for VGluT1 are not overlapped but rather positioned side-by-side. (C, D) DGKι immunoreactivity is detected in PKCα-positive rod bipolar cells (C) but not in calbindin-positive horizontal cells (D) in the OPL. (E) Comparison of DGKι and DGKβ. DGKι-positive puncta are detected in the postsynaptic region, whereas DGKβ positions in the subsynaptic and dendritic areas in rod bipolar cells. DIC, differential interference contrast image; GCL, ganglion cell layer; IPL, inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer; ISL, inner segment layer; OSL, outer segment layer. Scale bars = 20 µm (A); 10 µm (B–E).
Intense expression and localization of DGKι in the OPL motivated us to investigate whether DGKι and DGKβ are colocalized at the same region of bipolar cells. Double immunofluorescence imaging revealed that, at higher magnification, immunoreactivity for DGKι shows a punctate pattern of distribution, whereas that for DGKβ exhibits a rather diffuse pattern in the dendritic region. These immunoreactivities were not overlapped but rather positioned side-by-side (Fig. 2E). These data suggest that DGKι localizes at the postsynaptic region, whereas DGKβ positions in the subsynaptic and dendritic areas in rod bipolar cells.
DGKϵ
In our previous study, in situ hybridization histochemistry suggested that DGKϵ is the only isozyme expressed in photoreceptors (Sato et al. 2011). Immunohistochemical analysis showed that immunoreactivity for DGKϵ was detected intensely as granular structures in the inner segment layer (ISL) and diffusely as punctate structures throughout the OPL, INL, and IPL (Fig. 3A). At higher magnification, DGKϵ-positive puncta were restricted to a narrow line corresponding to the outer limiting membrane (OLM) (arrowheads, Fig. 3B).
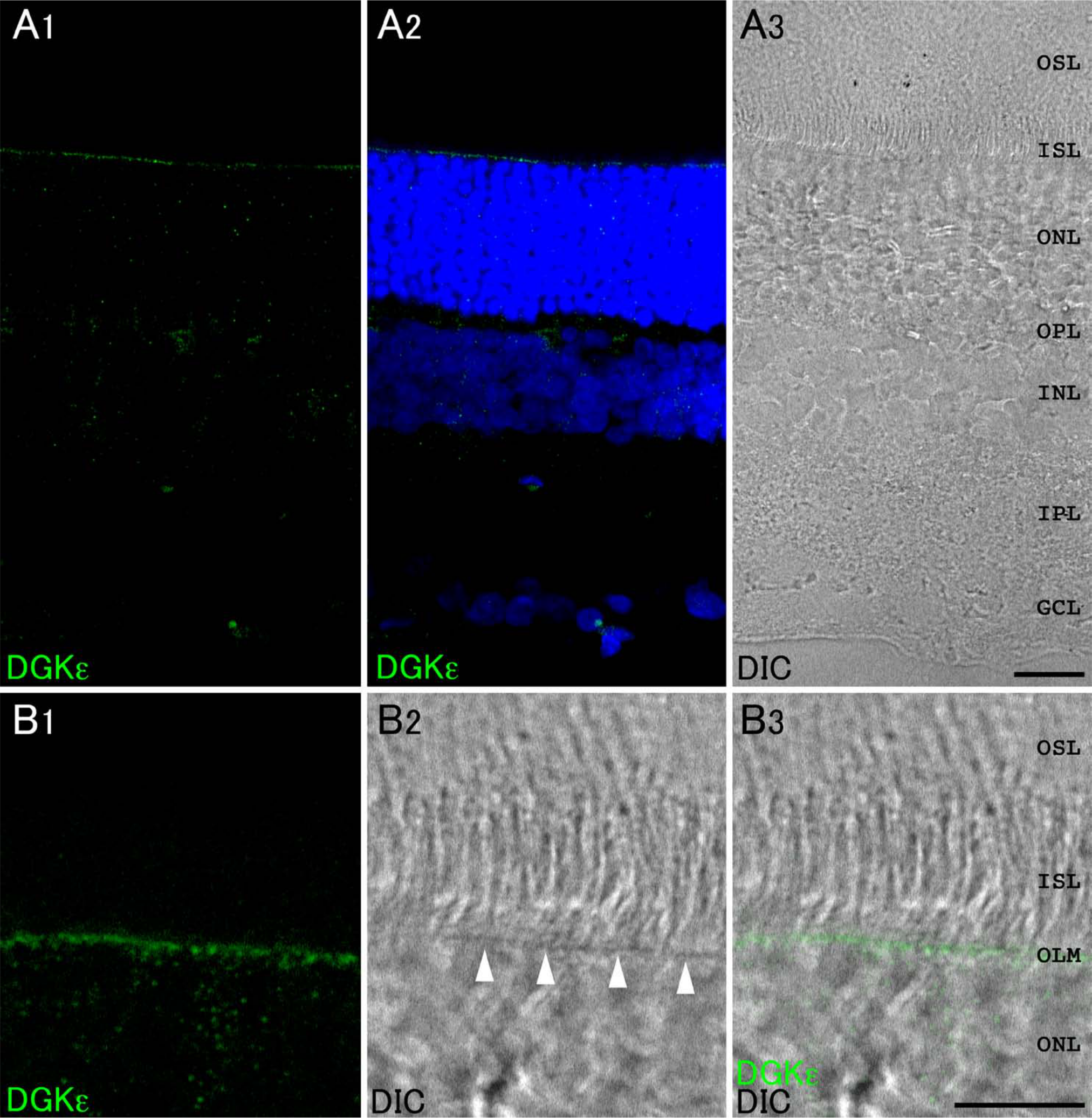
Expression and localization of DGKϵ in the retina. (A) Immunoreaction for DGKϵ is detected intensely as granular structures in the inner segment layer (ISL). Note that DGKϵ-positive puncta are restricted to a narrow line corresponding to the outer limiting membrane (OLM) (arrowheads, B). Blue fluorescence represents nuclear staining with TO-PRO-3. GCL, ganglion cell layer; IPL, inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer; OSL, outer segment layer. Scale bars = 20 µm (A); 10 µm (B).
DGKζ
DGKζ is shown to localize to the nuclei of most, if not all, neurons in the brain (Hozumi et al. 2003). Similar to this localization pattern in the brain, DGKζ immunoreactivity was detected clearly in the nuclei of ganglion cells in the ganglion cell layer (GCL) (Fig. 4A). In addition, the signals for DGKζ were found moderately in the OPL, INL, and IPL (Fig. 4A). To examine which cell type(s) were responsible for the expression, we next performed double immunolabeling experiments using antibodies against Brn-3a (a marker for ganglion cells) (Zhang et al. 2012), PKCα, calbindin D-28K, calretinin (a marker for amacrine cells) (Ciolofan et al. 2006), and glutamine synthetase (a marker for Müller cells) (Bui et al. 2009). As shown in the Fig. 4B, all the Brn-3a–positive ganglion cells were co-immunostained with DGKζ (arrowheads). Some of the DGKζ-positive cells in the GCL, which were not immunoreactive to Brn-3a (arrows), are likely to be displaced amacrine cells. DGKζ immunoreactivity was also detected in the nucleus of both PKCα-positive or PKCα-negative cells in the INL and in the axons of PKCα-positive rod bipolar cells in the IPL (Fig. 4C). DGKζ was coexpressed in cells that were immunoreactive for calbindin D-28K (Fig. 4D) and calretinin (Fig. 4E). Immunoreactivity for glutamine synthetase was not overlapped with that for DGKζ in the INL (Fig. 4F) and IPL (Fig. 4G). These findings suggest that DGKζ localizes in rod and cone bipolar cells, horizontal cells, amacrine cells, and ganglion cells. The signals for DGKζ were below detection levels in the photoreceptor cells.
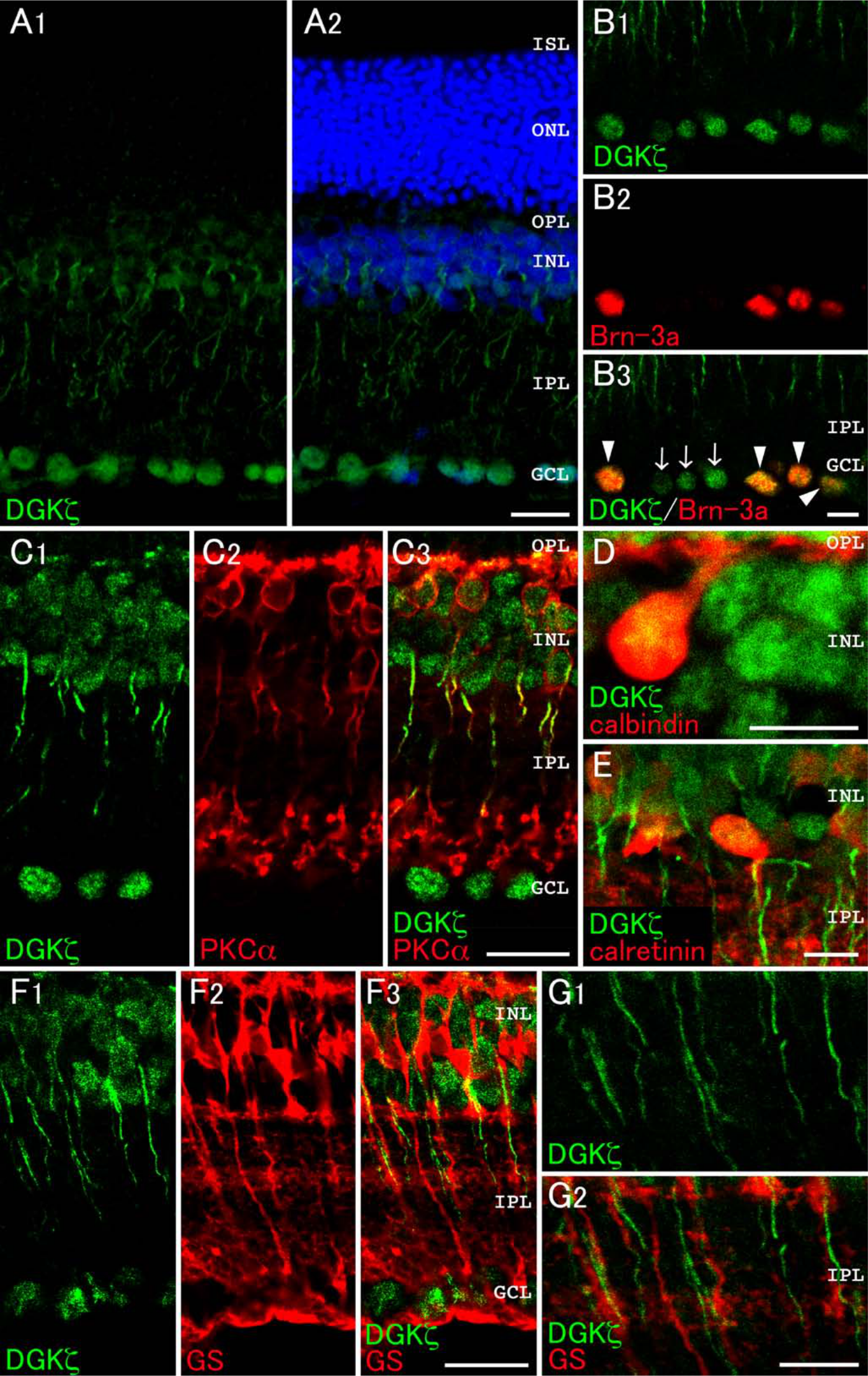
Expression and localization of DGKζ in the retina. Double immunofluorescence for characterization of DGKζ (green) in the retina (A–G). Red fluorescence represents Brn-3a (B), PKCα (C), calbindin D-28K (calbindin; D), calretinin (E), or glutamine synthetase (GS; F and G). (A) DGKζ immunoreactivity is detected in the GCL, IPL, INL, and OPL. (B) Immunoreactivity for DGKζ is observed in all the Brn-3a–positive ganglion cells (arrowheads). Note that some DGKζ-positive cells in the GCL are not immunoreactive for Brn-3a (arrows). (C) DGKζ immunoreactivity is detected in the nuclei and axons of PKCα-positive rod bipolar cells in the INL and IPL. (D, E) DGKζ is expressed in calbindin D-28K–positive horizontal cells (D) and calretinin-positive amacrine cells (E). (F, G) Immunoreactivity for glutamine synthetase is not colocalized with that for DGKζ in the INL (F) and IPL (G). GCL, ganglion cell layer; IPL, inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer; ISL, inner segment layer; OSL, outer segment layer. Scale bars = 20 µm (A, C, F); 10 µm (B, D, E, G).
DGKα
As predicted from its low mRNA expression level (Sato et al. 2011), DGKα immunoreactivity was very weak when using the conventional method. Therefore, we used the TSA Biotin System to intensify DGKα immunoreactivity. Immunoreactivity for DGKα was detected intensely in the GCL and weakly in the IPL (Fig. 5A). Because DGKα is shown to be expressed in oligodendrocytes, a glial cell type responsible for myelin formation in the central nervous system (Goto et al. 1992), we performed double immunofluorescence with glutamine synthetase, a marker for Müller cells. Results showed that no DGKα-immunoreactive puncta are found in cells that are immunoreactive for glutamine synthetase in the IPL and INL (Fig. 5A). The signals for DGKα were recognized in the nucleus of ganglion cells but were not detected in the inner limiting membrane (ILM) (Fig. 5B). Next, to determine if ganglion cells indeed express DGKα, we used the specific ganglion cell marker Brn-3a (Fig. 5C). All Brn-3a–positive ganglion cells were co-immunostained with DGKα (arrowheads). Some of the DGKα-positive cells in the GCL, which were not immunoreactive to Brn-3a (arrows), are presumed to be displaced amacrine cells. These data suggest that DGKα localizes primarily to the nucleus of ganglion cells but not to those of Müller cells.
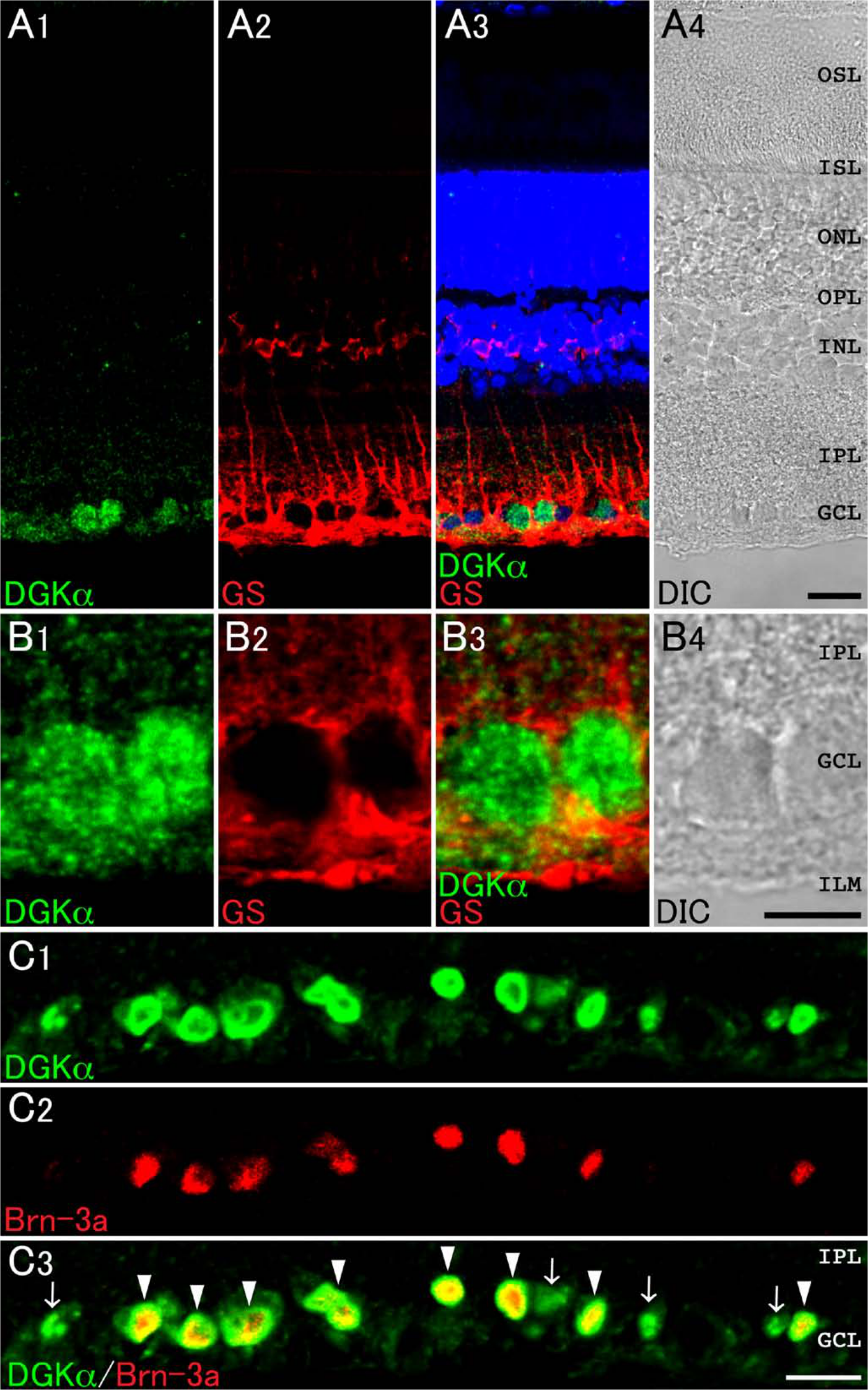
Expression and localization of DGKα in the retina. Double immunofluorescence for characterization of DGKα (green) in the retina (A–C). Red fluorescence represents glutamine synthetase (GS; A and B) and Brn-3a (C). (A) DGKα (green) is expressed intensely in the ganglion cell layer (GCL) and is not colocalized with GS-positive Müller cells (red). (B) A high-magnification image of the GCL. DGKα immunoreactivity is not detected in GS-positive inner limiting membrane (ILM). (C) Immunoreactivity for DGKα is observed in all the Brn-3a–positive ganglion cells (arrowheads). Note that some DGKα-positive cells in the GCL are not immunoreactive for Brn-3a (arrows). Blue fluorescence represents nuclear staining with TO-PRO-3. DIC, differential interference contrast image; IPL, inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer; ISL, inner segment layer; OSL, outer segment layer. Scale bars = 20 µm (A, C); 10 µm (B).
Expression and Localization of Other PI Signaling Molecules
Group I mGluRs, including mGluR1 and mGluR5, are known to operate PI signaling via G-proteins (Gilman 1987; Masu et al. 1991; Abe et al. 1992). A previous study revealed that both DGKβ and PLCβ4 are coexpressed in cells of the pituitary intermediate lobe, and they localize near the plasma membrane (Hozumi et al. 2010). Furthermore, DGKβ and PLCβ1 are shown to localize side-by-side together with mGluR5 or DGLα in the striatum (Hozumi et al. 2008). Therefore, we investigated whether PI signaling molecules, such as group I mGluRs, Gαq/11, PLCβ subfamily, conventional PKCs, and DGLα, are expressed in the retina and, if so, where they localize. Results showed that the immunohistochemical signals for Gαq/11, mGluR1α, mGluR5, PLCβ1, PLCβ3, PLCβ4, PKCα, PKCβII, and DGLα were detected in rat retina, showing that PI signaling molecules are expressed broadly in this organ (Fig. 6).
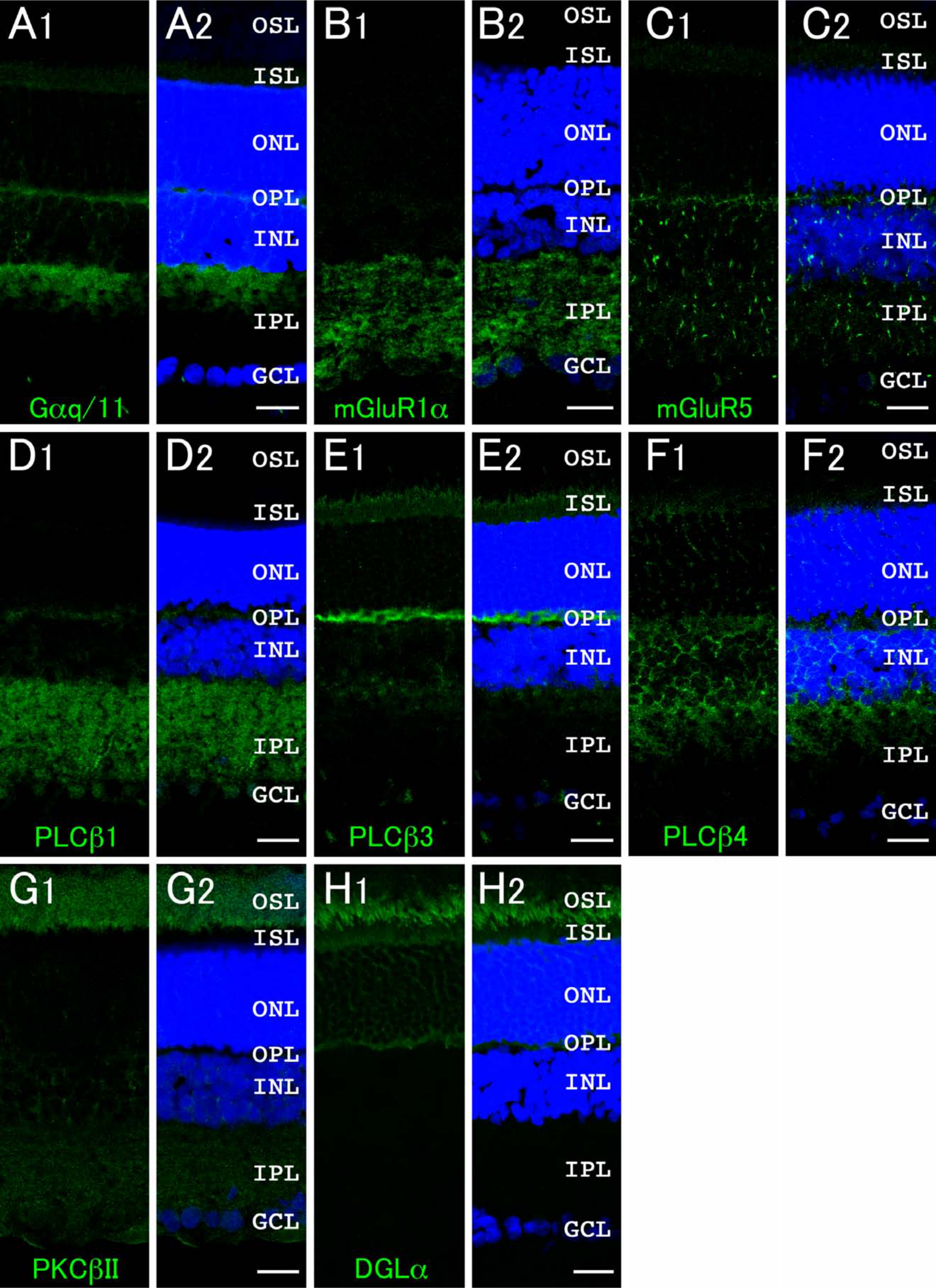
Expression and localization of phosphoinositide signaling molecules in the retina. Green fluorescence represents Gαq/11 (A), mGluR1α (B), mGluR5 (C), PLCβ1 (D), PLCβ3 (E), PLCβ4 (F), PKCβII (G), and DGLα (H). Blue fluorescence represents nuclear staining with TO-PRO-3 (A–H). GCL, ganglion cell layer; IPL, inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer; ISL, inner segment layer; OSL, outer segment layer. Scale bars = 20 µm (A–H).
Immunoreactivity for Gαq/11, as well as the α subunit of the Gq subclass of GTP-binding proteins that mediate signals from receptors to the PLC, was detected intensely in the outer half of the IPL (OFF sublamina) and OPL and weakly in the INL and ISL (Fig. 6A), suggesting that the PI signaling cascade may exist in photoreceptor cells, bipolar cells, and/or horizontal cells. The signals for mGluR1α were below detection levels in the photoreceptor cell area, including the OPL, outer nuclear layer (ONL), ISL, and outer segment layer (OSL), but were detected intensely in the IPL (Fig. 6B). However, the signals for mGluR5 were recognized moderately throughout the OPL, INL, and IPL and weakly in the ISL (Fig. 6C). PLCβ isoforms exhibited distinct expression patterns: PLCβ1 signals were abundant throughout the IPL and moderately in the OPL (Fig. 6D), whereas PLCβ3 signals were detected intensely in the OPL and moderately in the ISL (Fig. 6E). PLCβ4 signals were visible moderately in the OPL, INL, and the outer half of the IPL (OFF sublamina) and weakly in the ISL (Fig. 6F). No significant signal was detected for PLCβ2 in the retina (data not shown).
Regarding the downstream molecules of PI turnover, PKCβII immunoreactivity was predominantly detected in the OSL (Fig. 6G), whereas PKCγ signals were below detection levels (data not shown). As shown in Figs. 1C and 2C, PKCα is well known as a marker for rod bipolar cells (Whitaker and Cooper 2009). The signals for DGLα, which hydrolyzes the sn-1 acyl chain of DG to produce 2-arachidonoyl glycerol (2-AG), were recognized solely in the photoreceptor cell area with the most intense signals in the OSL (Fig. 6H).
Discussion
This study revealed for the first time the expression and localization of DGKs in mammalian retina at the protein level using specific antibodies. We show clearly that DGK isozymes exhibit distinct localization patterns in the rat retina. These findings suggest that DGKs are involved in photosignal transduction and that they play different roles in distinct retinal cells. We also confirmed the retinal expression of other components of a possible pathway involving the glutamate-induced PI signaling cascade, such as group I mGluRs, Gαq/11, PLCβs, PKCs, and DGL. Table 1 summarizes the distribution of PI signaling molecules including DGK isozymes in rat retina. What are the functional roles of DGKs in retinal cells?
Distribution of PI Signaling Molecules in Rat Retina.
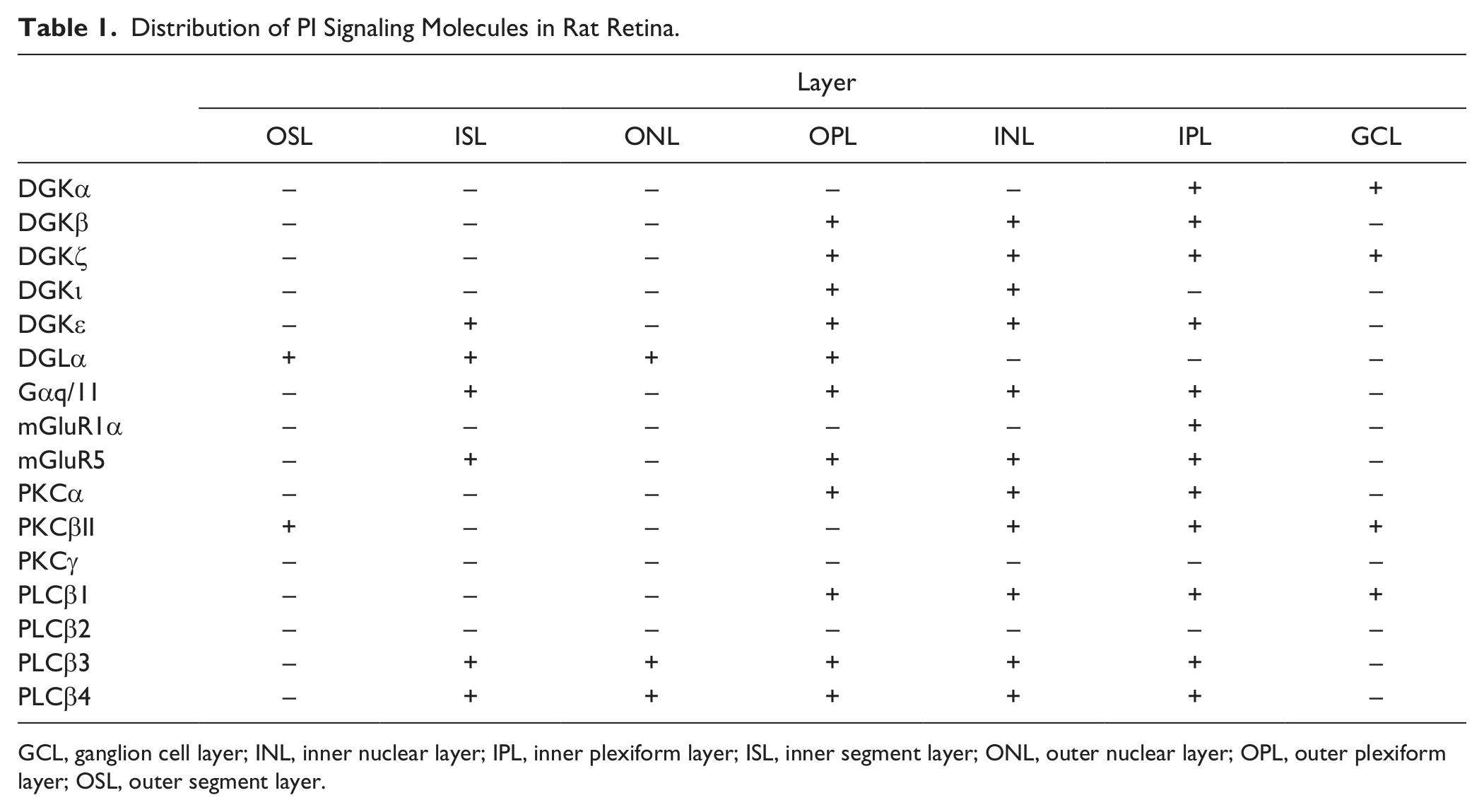
GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; ISL, inner segment layer; ONL, outer nuclear layer; OPL, outer plexiform layer; OSL, outer segment layer.
First, DGKβ and DGKι are revealed to localize in the OPL. Reportedly, the flow of visual information in the vertical direction from the photoreceptors to bipolar cells and horizontal cells in the OPL and from bipolar cells to ganglion cells and amacrine cells in the IPL is mediated via glutamate (Massey 1990; Watanabe et al. 1994; Bui et al. 2009). Neurotransmitter glutamate triggers signaling through two classes of receptors: ionotropic and metabotropic glutamate receptors. Ionotropic glutamate receptors mediate fast excitatory synaptic transmission, whereas mGluRs are G-protein–coupled receptors that bind the ligand and modulate neuronal activity through intracellular second messengers (Seeburg 1993; Hollmann and Heinemann 1994; Nakanishi 1994). Of the mGluR family, mGluR1 and mGluR5, both belonging to the group I mGluRs, are functionally coupled to Gq-mediated PLCβ stimulation, which triggers the DG-dependent signaling cascade (Gilman 1987; Masu et al. 1991; Abe et al. 1992; Nakanishi 1994).
In agreement with results of earlier studies (Koulen et al. 1997), the group I mGluRs were detected in both OPL and IPL layers (Fig. 6B,C). Simultaneously, Gαq/11, the major isoform of the Gq protein α subunits in the central nervous system (Tanaka et al. 2000), and its intracellular effector PLCβ were found to be expressed in both synaptic layers (Fig. 6D–F). These data suggest that synaptic signals between photoreceptors and bipolar/horizontal cells and between bipolar cells and ganglion/amacrine cells are transmitted through the glutamate-triggered PI signaling cascade.
In this regard, DGKβ and DGKι, both of which are expressed predominantly in the OPL but not in the IPL, might be involved in DG-mediated signaling in postsynaptic sites of PKCα-positive rod bipolar cells. What is the functional difference of these DGK isozymes in this case? Notably, DGKβ and DGKι localize to closely related but distinct subcellular sites: DGKι-positive puncta localize predominantly to the postsynaptic region apposed to the photoreceptor synapses labeled with VGluT1, whereas DGKβ distributes in subsynaptic and dendritic areas. Recently, we reported that DGKβ is highly concentrated at the perisynapses of dendritic spines and that it might play an important role in spine maturation/remodeling through actin filament assembly (Hozumi and Goto 2012). However, DGKι is shown to localize diffusely to the cytoplasm in hippocampal CA1 neurons (Ito et al. 2004). From a functional perspective, DGKι negatively regulates RasGRP3 (Regier et al. 2005), a DG effector with guanine nucleotide exchange factor activity for Ras and Rap small GTPases (Yamashita et al. 2000). These small G-proteins are shown to regulate synaptic/spine structure and synaptic plasticity (Zhu et al. 2002). It is therefore conceivable that DG signals triggered by glutamate may be diversified into distinct cascades through different isoforms of PLCβ in bipolar cells to regulate synaptic transmission and/or plasticity.
Second, DGKϵ displays a prominent, restricted expression pattern in the retina: DGKϵ is distributed predominantly to the OLM, which is located at the neck of the inner segment. The OLM consists of desmosome-like adherens junctions and their “terminal web” of cytoplasmic filaments in photoreceptors and in the Müller cell “collars” that surround each photoreceptor (Ciolofan et al. 2006). Because our in situ hybridization histochemistry shows that DGKϵ mRNA signal is detected in photoreceptors (Sato et al. 2011), the DGKϵ-immunoreactive structure should be a component of photoreceptor cells. Previous reports have described that the OLM is immunoreactive for adherens junction–related molecules, including zonula occludens-1 (Puller et al. 2009), N-cadherin (Koike et al. 2005), and membrane-associated palmitoylated proteins (Kantardzhieva et al. 2006). These molecules, which are well known as intracellular components of tight and adherens junctions, are involved in the maintenance of photoreceptor-Müller glial cell adhesion (Kantardzhieva et al. 2006). In addition, the tight barrier may help to establish and maintain the unequal distribution of membrane lipids and proteins in polarized cells, such as epithelia, photoreceptors, and other neurons (Altschuler et al. 2003). How DGKϵ contributes to photoreceptor function at this specific site remains undetermined. One possibility is suggested by the structural feature of the outer segment and the enzymatic property of DGKϵ: The outer segment is a site of the initial step of photoreception in photoreceptor cells. It comprises hundreds of stacked rhodopsin-laden membranous disks enveloped by the plasma membrane. Phospholipids for the disks are assumed to be provided by the inner segment. Furthermore, it is noteworthy that the outer segment membrane is characteristically enriched in polyunsaturated fatty acids, such as arachidonic acid and docosahexaenoic acid (Simopoulos and Bazán 2009). The reason for this enrichment remains uncertain, although it is speculated that phospholipids with polyunsaturated fatty acids stabilize the outer segment membrane structure. DGKϵ acts preferentially on arachidonoyl-DG, which is a major component in PI turnover, suggesting that DGKϵ is probably engaged in the PI cycle. Therefore, it might be hypothesized that DGKϵ plays a role in enrichment of arachidonoyl-containing phospholipids in the disk membrane at this site. However, it remains unclear how DGKϵ is specifically targeted to the OLM. Previous reports have described that DGKϵ is localized to the endoplasmic reticulum in transfected cells (Kobayashi et al. 2007), which is not true in retinal cells. It would be interesting to investigate whether DGKϵ is associated with adherens junction proteins, which can serve as scaffolding proteins (Kantardzhieva et al. 2006; Puller et al. 2009).
Regarding other DGK isozymes, DGKζ and DGKα are shown to localize predominantly to the nucleus of ganglion cells. Nuclear localization of DGKζ is compatible with our previous reports, which shows that DGKζ localizes to the nucleus in neurons of various types in the brain (Goto and Kondo 1996; Hozumi et al. 2003). However, the nuclear localization of DGKα differs greatly from its feature in the brain. In our previous study, DGKα was shown to be expressed in oligodendrocytes, a glial cell responsible for myelin formation, and localized to myelin structures in the central nervous system (Goto et al. 1992). This discrepancy remains unclear, but DGKα can localize to the nucleus in transfected cells, suggesting that DGKα is nuclear in some instances (Kobayashi et al. 2007). Although the functional roles of these isozymes in the nucleus remain obscure, previous studies implicate their contradictory effects on cell cycle regulation: DGKα as a positive regulator (Flores et al. 1996; Cipres et al. 2003) and DGKζ as a negative regulator (Evangelisti et al. 2007). However, because retinal ganglion cells are postmitotic neurons that do not proliferate anymore at adult ages, DGKα and DGKζ might be engaged in other mechanisms different from cell cycle regulation. Further study is necessary to elucidate this point.
Finally, it should be described that we detected no significant immunohistochemical signals of DGK isozymes in the outer segment, which is the primary site of photoreception in photoreceptor cells under normal lighting conditions. However, previous studies have shown high levels of expression of DGKγ in the outer segments of bovine and human retina (Kai et al. 1994; Huang et al. 2000), suggesting that DGK isozymes exhibit a distinct expression pattern in the retina among animal species. We find in the outer segment intense signals for PKCβII, which is activated by DG, and DGLα, which hydrolyzes DG into 2-AG, a retrograde transmitter endocannabinoid, under normal lighting conditions (Fig. 6). Taken together, these results raise the possibility that DG signaling in the outer segment is terminated by DGLα or other DGKs that have not been examined yet.
In conclusion, DGK isozymes, together with other PI signaling molecules, are expressed broadly in the retina. Their distinct cellular expression and subcellular localization patterns suggest that each isozyme plays a unique role in photosignal transduction. The findings presented herein provide clues that elucidate functional aspects of DGKs in the pathophysiology of the retina. Recently, there is increasing evidence of a role for DG-induced PKC activation contributing to the vascular complications that arise in diabetic retinopathy (Hammes et al. 2003; Das Evcimen and King 2007). In addition, it is reported that total DG levels and PKCβII in rat retina are increased in diabetes (Shiba et al. 1993). These data suggest the importance of DG-dependent signaling in diabetic retinopathy. We are currently developing and conducting more detailed analyses by electron microscopy. Furthermore, it is warranted to investigate whether the expression and localization of DGKs might be altered in light and dark conditions or in diabetic retinopathy.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants-in-aid from The Ministry of Education, Culture, Sports, Science and Technology (MEXT) of Japan (YH, KG).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.