
Abstract
The liver possesses the capacity to restore its function and mass after injury. Liver regeneration is controlled through complicated mechanisms, in which the phosphoinositide (PI) cycle is shown to be activated in hepatocytes. Using a rat partial hepatectomy (PH) model, the authors investigated the expression of the diacylglycerol kinase (DGK) family, a key enzyme in the PI cycle, which metabolizes a lipid second-messenger diacylglycerol (DG). RT-PCR analysis shows that DGKζ and DGKα are the major isozymes in the liver. Results showed that in the process of regeneration, the DGKζ protein, which is detected in the nucleus of a small population of hepatocytes in normal liver, is significantly increased in almost all hepatocytes. However, the mRNA levels remain largely unchanged. Double labeling with bromodeoxyuridine (BrdU), an S phase marker, reveals that DGKζ is expressed independently of DNA synthesis or cell proliferation. However, DGKα protein localizes to the cytoplasm in normal and regenerating livers, but immunoblot analysis reveals that the expected (80 kDa) and the lower (70 kDa) bands are detected in normal liver, whereas at day 10 after PH, the expected band is solely recognized, showing a different processing pattern of DGKα in liver regeneration. These results suggest that DGKζ and DGKα are involved, respectively, in the nucleus and the cytoplasm of hepatocytes in regenerating liver.
The liver is a unique organ in that it has a potential capacity to restore the function and mass after injury. Under normal conditions, hepatocytes rarely divide, although it has long been known that after resection of 70% of rat liver (left and middle lobes), the remaining lobe immediately starts to proliferate and grow back almost completely within a month (Alison 1986; Fausto 2000). Liver regeneration is controlled through complicated biological responses to injury, which involve growth factors, cytokines, and cell cycle regulators (Michalopoulos and DeFrances 1997; Fausto 2000; Crljen et al. 2004). Previous reports have described that, in the process of liver regeneration, the phosphoinositide (PI) cycle might be activated in the hepatocyte nucleus where the levels of phosphatidylinositol-4,5-bisphosphate (PIP2) and diacylglycerol (DG) are changed during the process (Bocckino et al. 1989; Banfic et al. 1993; Mazzotti et al. 1995; Bregoli et al. 2002). Indeed, it is demonstrated that the nucleus is equipped with most of the PI cycle–related enzymes, including phospholipase C (PLC) and diacylglycerol kinase (DGK) (Goto et al. 2006).
DGK is an enzyme responsible for the phosphorylation of DG, which is derived from PIP2 by the action of PLC, to phosphatidic acid (PA). Both of the lipids are important components of phospholipid biosynthesis in the PI cycle (Kanoh et al. 1990). A well-known functional role of DGK is the regulation of protein kinase C (PKC), for which DG acts as an allosteric activator and whose activity plays a central role in many different cell types (Sakane and Kanoh 1997). The DG-PKC pathway is implicated in widely various cellular processes such as cellular growth, differentiation, and secretion (Nishizuka 1984). In addition, PA, the product of DGK, reportedly serves as a second messenger (Tsai et al. 1990; Moritz et al. 1992; Jones GA and Carpenter 1993; Limatola et al. 1994). Consequently, DGK is regarded as a key enzyme in the PI signal transduction pathway by controlling the levels of both DG and PA. To date, 10 isozymes of mammalian DGKs have been identified. They have been shown to differ from each other with respect to molecular size, enzymological properties, and differential tissue distribution (Goto et al. 2007; Sakane et al. 2007; Topham and Epand 2009). These data strongly suggest that each isozyme has its own specific function in various biological processes. In this respect, we show that DGKζ, containing a nuclear localization signal, localizes to the nucleus in cells of various kinds (Goto and Kondo 1996; Hozumi et al. 2003; Katagiri et al. 2005; Goto et al. 2007). Collectively, these results strongly suggest that nuclear PI-related enzymes, including DGK, are intimately involved in the liver regeneration process and that they regulate the intranuclear events (Jones DR and Divecha 2004).
In this study, we sought to elucidate the functional significance of DGK isozymes in the process of liver regeneration using a rat partial hepatectomy model. We show that the expressions of DGKζ and DGKα are altered, respectively, in the nucleus and the cytoplasm of hepatocytes in a regenerating liver.
Materials and Methods
Partial Hepatectomy Model and BrdU Inoculation
All animal experiments were conducted in accordance with the Yamagata University Guide for the Care and Use of Laboratory Animals. A rat partial hepatectomy model was used according to the protocol described in a previous report (Crljen et al. 2004). The hepatectomized and sham-operated animals were killed on days 1, 5, 10, and 30 after partial hepatectomy (PH) (n=10, 3, 14, and 3 for days 1, 5, 10, and 30, respectively). One hour before they were killed, rats were given a single intraperitoneal injection of bromodeoxyuridine (BrdU; Roche Diagnostics Corp., Mannheim, Germany) at a dosage of 100 mg/kg body weight.
Tissue Preparation and Immunohistochemistry
The livers (hepatectomized and sham operated) were processed for immunohistochemistry as described previously (Nakano et al. 2006). To observe BrdU incorporation, sections were treated with 2N HCl for 30 min at 37C, with subsequent neutralization with 0.1 M boric acid. The primary antibodies used were anti-DGKζ (0.1 µg/ml), DGKα (1:3000), PKCα (1.0 µg/ml), and BrdU (1:1000; Roche) antibodies in PBS containing 5% NGS and 0.1% Tween-20 (Goto et al. 1992; Hozumi et al. 2003; Nakano et al. 2006). Immunolabeling was accomplished with diaminobenzidine tetrahydrochloride (DAB) containing nickel chloride or immunofluorescence. Images were taken using a confocal laser-scanning microscope (PASCAL; Carl Zeiss, Inc., Oberkochen, Germany).
Immunoblot Analysis and RT-PCR
Normal and regenerating livers (1 and 10 days after PH) were freshly removed and homogenized in buffer containing 50 mM Tris-HCl (pH 7.5), 0.25 M sucrose, 20 mM KCl, 4 mM EGTA, and a complete protease inhibitor mixture (Roche). After removal of debris using low-speed centrifugation (550 × g) for 5 min, the supernatant was used as total lysate. The nuclear fraction was isolated as described by Sindic et al. (2001). The total lysate was used for immunoblot analysis of DGKα and PKCα, whereas the nuclear fraction was used for DGKζ. Samples were subjected to SDS/10% PAGE. Immunoreactive bands for anti-DGKζ, DGKα, PKCα, and PKCγ antibodies were visualized using the chemiluminescent ECL + Plus Western blot detection system (GE Healthcare, Buckinghamshire, UK) as described (Hozumi et al. 2003; Yoshida et al. 2006). Total RNA was extracted from normal and regenerating livers and was used as a template for reverse transcription with subsequent PCR as described previously (Katagiri et al. 2005). Band intensities were quantified by densitometry using Image J (National Institutes of Health, Bethesda, MD) as described previously (Evangelisti et al. 2010).
Cell Culture
Isolation and purification of rat primary hepatocytes were performed and cultured as described previously (Okumoto et al. 2003). The HepG2 cells were cultured in DMEM containing 25 mM glucose and 10% FBS at 37C and 5% CO2. Cultured cells were prepared for RT-PCR and for immunocytochemistry as described above.
Results
Six DGK isozymes were cloned from rat tissues. We first performed RT-PCR analysis to examine the expression profile of DGKs in the liver under normal conditions. Figure 1 shows that expression signals were detected intensely for DGKζ and DGKα and weakly for DGKϵ in normal liver. The signals for the other DGKs were faint or below detection level. The result shows that DGKζ and DGKα are the major isozymes in normal liver. Therefore, we specifically examined these two isozymes in the following experiments.
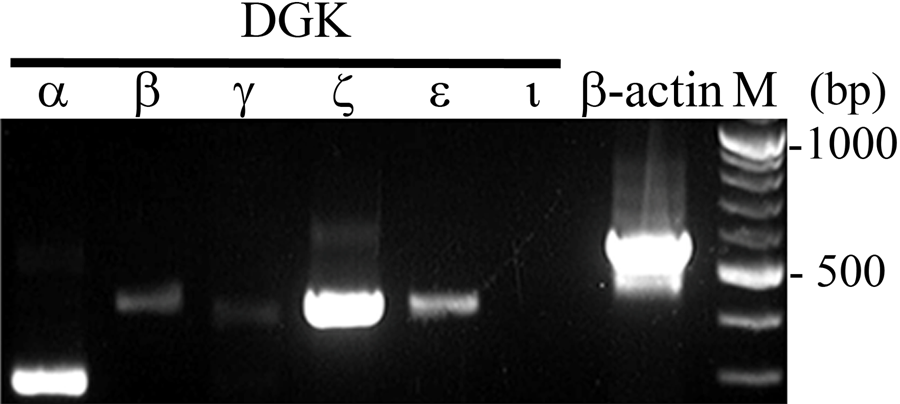
Expression of diacylglycerol kinase (DGK) isozyme mRNA in normal rat liver. RT-PCR analysis using specific primer for each DGK isozyme. β-Actin primer was used as a control. M, size marker.
In normal liver, DGKζ was expressed most abundantly at the mRNA level. We next examined the subcellular localization of this isozyme using immunohistochemistry (Fig. 2A). In normal liver, DGKζ-immunoreactivity was sparsely and unevenly distributed in the hepatic lobule and recognized intensely in the nucleus of hepatocytes. The immunoreactive cells were estimated roughly as comprising around 10% of the total cell population under normal conditions. At day 1 after PH, DGKζ-immunoreactive cells were increased significantly in number and comprised almost all the cells. The immunoreactivity was recognized in the nucleus of cells at various intensities—that is, some cells were stained intensely, as in a normal liver, although numerous cells were stained weakly (Fig. 2A, arrows). At day 5 after PH, DGKζ-immunoreactivity was further increased in intensity in the nucleus of hepatocytes. At day 10, almost all the cells remained immunopositive in the nucleus, and the immunoreactivity was greatly increased in intensity in each cell compared with that at days 1 and 5 after PH (Fig. 2A, arrowheads). At day 30, the immunoreactivity returned to the level seen in the normal liver. In immunoblot analysis of the nuclear fraction, DGKζ protein was estimated as increasing about sixfold (as determined by semi-quantitative densitometric analysis) at day 10 after PH compared with that in normal liver (Fig. 2B).
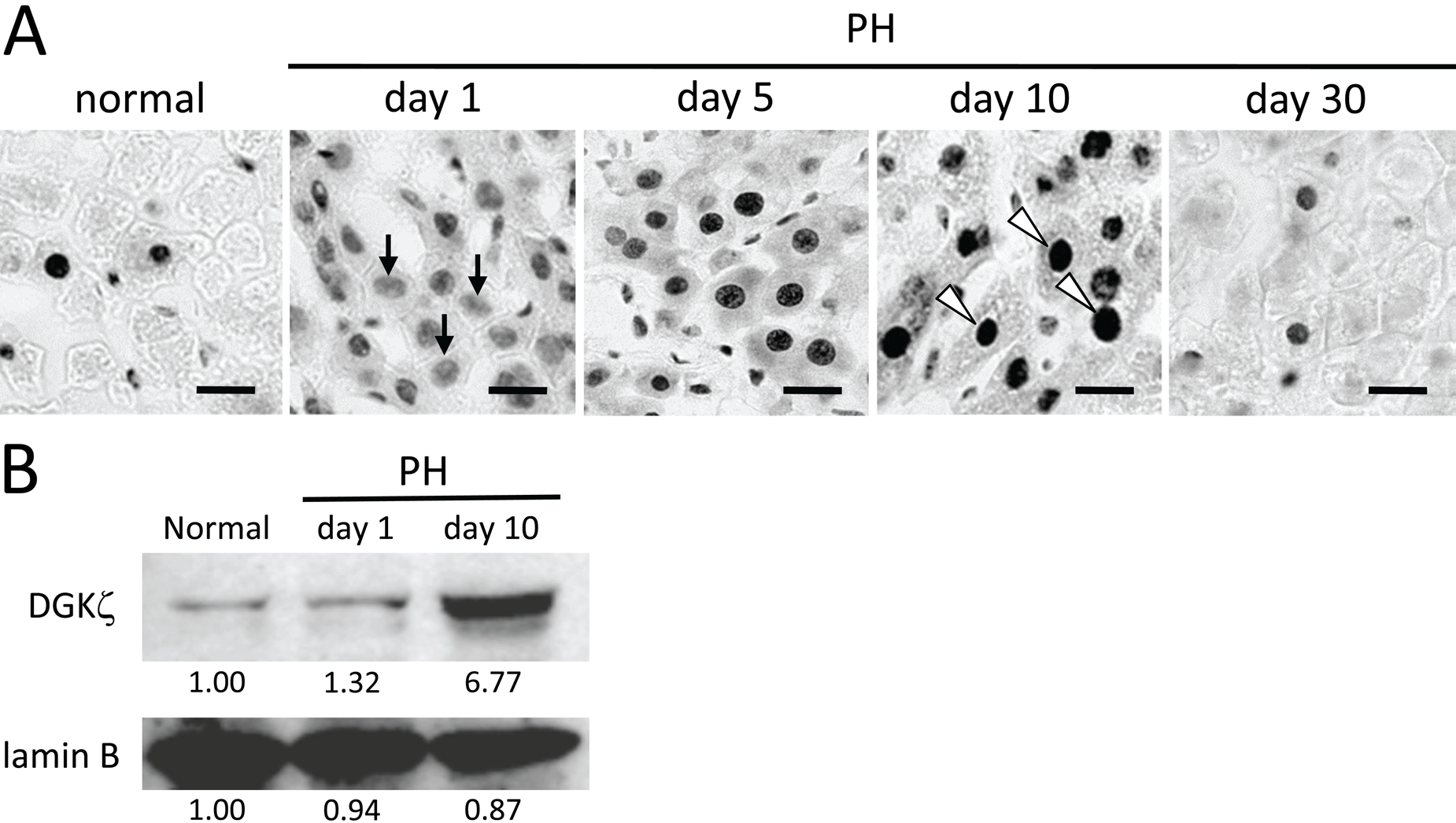
Expression of DGKζ in normal and regenerating livers. (A) Immunohistochemically, DGKζ-immunoreactivity is sparsely distributed and recognized intensely in the nucleus of hepatocytes in normal liver. At day 1 after partial hepatectomy (PH), DGKζ-immunoreactive cells are significantly increased in number, although the intensity of immunoreactivity in each cell is low (arrows). At day 5, DGKζ-immunoreactivity is further increased in intensity in the nucleus of hepatocytes. At day 10, almost all of the cells show intense immunoreactivity in the nucleus (arrowheads). At day 30, the immunoreactivity returns to the level seen in the normal liver. (B) Nuclear fractions (20 µg) were analyzed by immunoblot analysis using anti-DGKζ antibody. DGKζ protein is prominently increased at day 10 after PH compared with that in normal liver. Anti–lamin B antibody was used as a control for nuclear fraction. Blots were scanned, and band intensities were quantified by Image J (National Institutes of Health, Bethesda, MD) densitometry analysis as described in Materials and Methods. Bands of the control were normalized to 1.00. Bars: 20 µm.
We next investigated whether the increased expression of DGKζ after PH is regulated at the transcriptional level (Fig. 3). Intriguingly, RT-PCR analysis revealed that the DGKζ mRNA expression level remains largely unchanged in samples obtained from normal liver and livers at day 1 and 10 after PH. This was in sharp contrast to a marked (about sixfold) increase in protein amount at day 10 after PH, suggesting that the increased DGKζ protein is not ascribed to upregulated transcriptional activity.
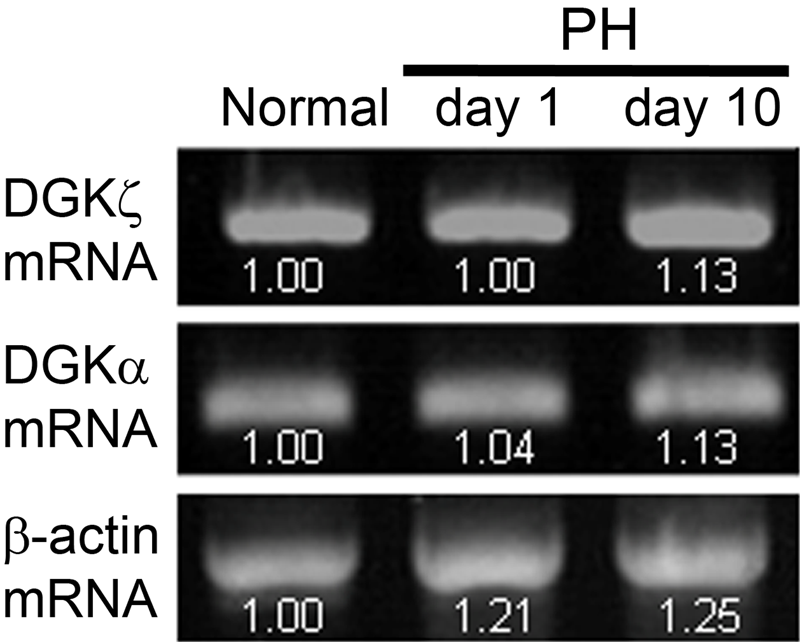
RT-PCR analysis of DGKα and DGKζ in regenerating liver. Expression levels for DGKα and DGKζ mRNAs remain largely unchanged in samples obtained from normal liver and livers at days 1 and 10 after partial hepatectomy (PH). β-Actin primer was used as a control. PCR was performed for 35 cycles. Band intensities were quantified by Image J (National Institutes of Health, Bethesda, MD) densitometry analysis, and bands of the control were normalized to 1.00. Similar results were obtained in PCR for 30 and 32 cycles.
Active DNA synthesis is known to occur for cell proliferation at day 1 after PH (Michalopoulos and DeFrances 1997). To examine whether the expression of DGKζ coincides with DNA synthesis in regenerating liver, we performed double immunostaining for DGKζ and incorporated S phase marker BrdU (Fig. 4). In the normal liver, a small fraction of cells, around 10% of the total cell population, were immunoreactive for DGKζ, although no BrdU-positive cells were detected. In the regenerating liver at day 1 after PH, almost all the cells were DGKζ-immunoreactive, about 30% of which showed BrdU-immunoreactivity. At day 10 after PH, DGKζ-immunoreactive cells remained mostly unchanged, although BrdU-positive cells were not detected any more. These results suggest that the expression of DGKζ is regulated independently of DNA synthesis in both normal and regenerating livers.
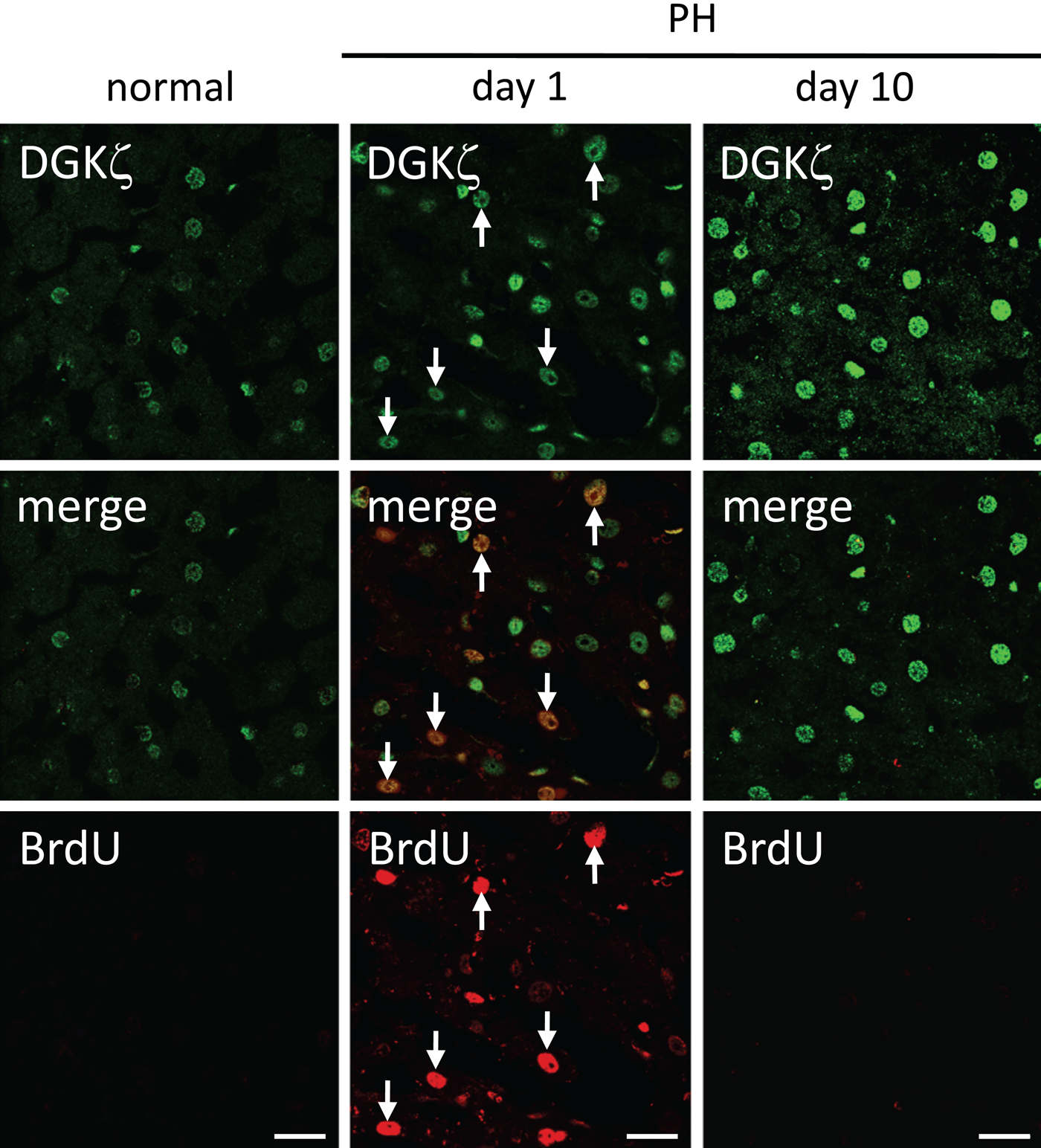
Expression of DGKζ in S phase cells in regenerating livers. BrdU was inoculated intraperitoneally 1 hr before sacrifice to identify cells in S phase. In normal liver, a small fraction of cells are immunoreactive for DGKζ, whereas no BrdU-positive cells are detected. At day 1 after partial hepatectomy (PH), almost all of the cells are DGKζ-immunoreactive, ~30% of which show BrdU-positive (arrows). At day 10 after PH, DGKζ-immunoreactive cells remain largely unchanged, although BrdU-positive cells are no longer detected. Bars: 20 µm.
Proliferation of hepatocytes in the liver is strictly regulated by several factors, such as growth factors and cytokines, produced by surrounding and circulating cells (Alison 1986; Michalopoulos and DeFrances 1997). To investigate the expression patterns of DGKζ in cells in culture, which provides a controlled environment, we used primary cultured hepatocytes from normal rat liver and HepG2 cells derived from human hepatoma (Fig. 5). In immunocytochemical analysis, all the hepatocytes in culture at in vitro days 3 and 5 showed intense DGKζ-immunoreactivity in the nucleus (Fig. 5A). In contrast, HepG2 cells exhibited continuous proliferation. RT-PCR revealed in this cell line that DGKζ mRNA was solely detected; the other isozymes were below detection level, suggesting that DGKζ is the most abundant isozyme in HepG2 cells (data not shown). In immunocytochemical analysis, all the cells were intensely immunoreactive for DGKζ. In double staining, around 40% of the DGKζ-immunoreactive cells were positive for BrdU incorporated (Fig. 5B). The pattern of DGKζ-immunoreactivity in primary hepatocytes and HepG2 cells clearly differs from that in normal liver, but it is similar to that in regenerating liver in terms of the DGKζ-immunoreactive ratio in the cell population. Taken together, it is likely that DGKζ is expressed independently of DNA synthesis or cell proliferation in culture conditions and in the liver in situ.
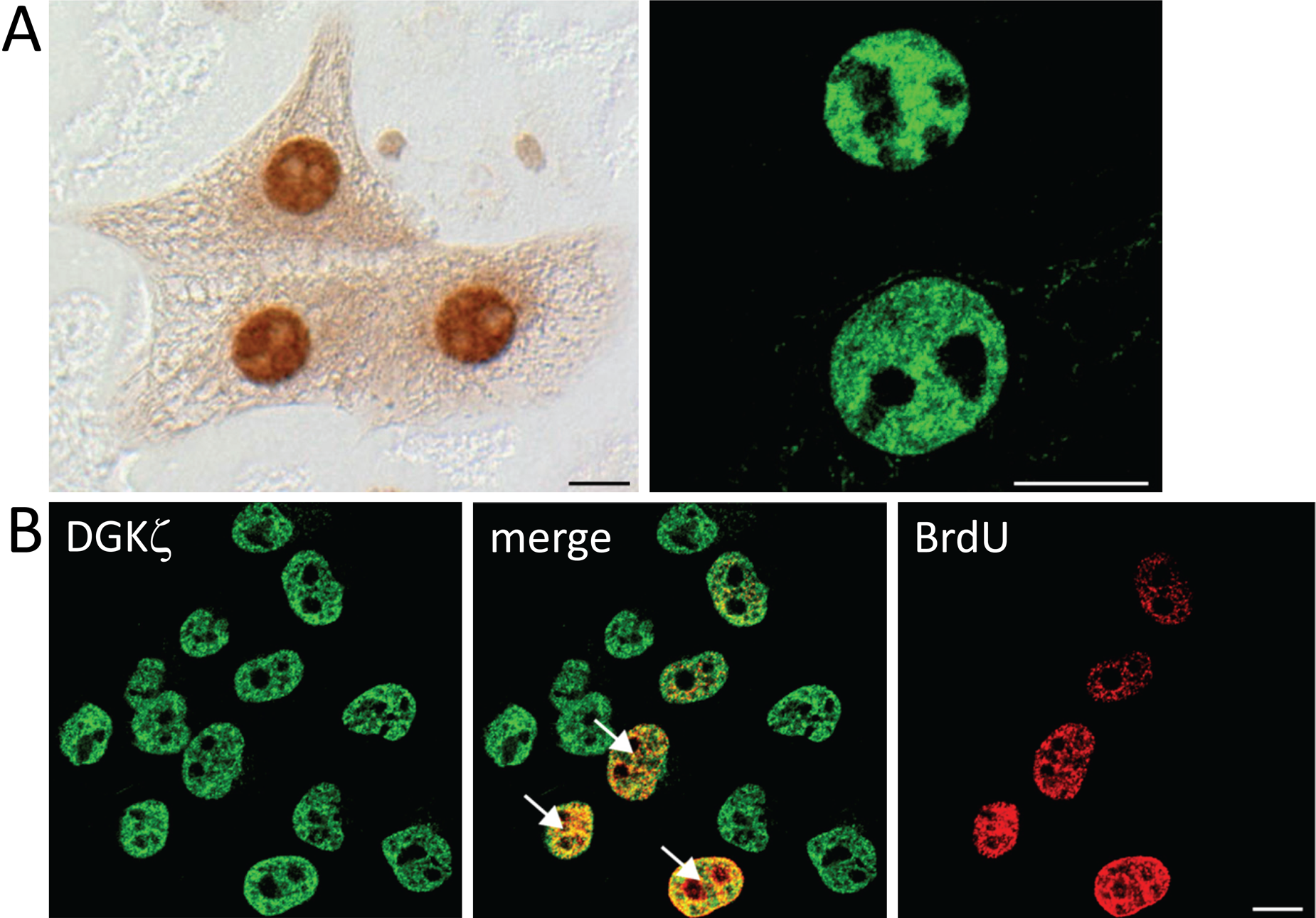
Expression of DGKζ in primary cultured rat hepatocytes (A) and HepG2 cells (B). (A) Immunocytochemically, all of the hepatocytes in primary culture at in vitro days 3 and 5 show intense DGKζ-immunoreactivity in the nucleus, visualized by diaminobenzidine (left) and the confocal image (right). (B) HepG2 cells were incubated with 10 µM BrdU for 10 min to label cells in S phase. DGKζ-immunoreactivity is intensely detected in all of the HepG2 cells, ~40% of which are positive for BrdU incorporated (arrows). Bars: 10 µm.
DGKα was also abundantly expressed at the mRNA level in normal liver (Fig. 1). In immunohistochemical analysis, DGKα-immunoreactivity was detected diffusely in the cytoplasm of all hepatocytes under normal conditions (Fig. 6A). This pattern of the expression and localization remained unchanged in regenerating livers. RT-PCR analysis showed no marked alteration of mRNA expression in normal and regenerating livers (Fig. 3). However, immunoblot analysis of DGKα should be noted particularly (Fig. 6B). In normal liver, anti-DGKα antibody recognized a lower band (70 kDa) in addition to the expected band (80 kDa), which had been reported in a previous study (Goto et al. 1992). In this regard, the intriguing pattern of the immunoreactive bands was observed in samples from regenerating liver: At day 1 after PH, two bands (80 kDa and 70 kDa) were recognized as observed in normal liver, whereas at day 10 after PH, a single band of the expected length (80 kDa) was recognized and the lower band (70 kDa) almost disappeared.
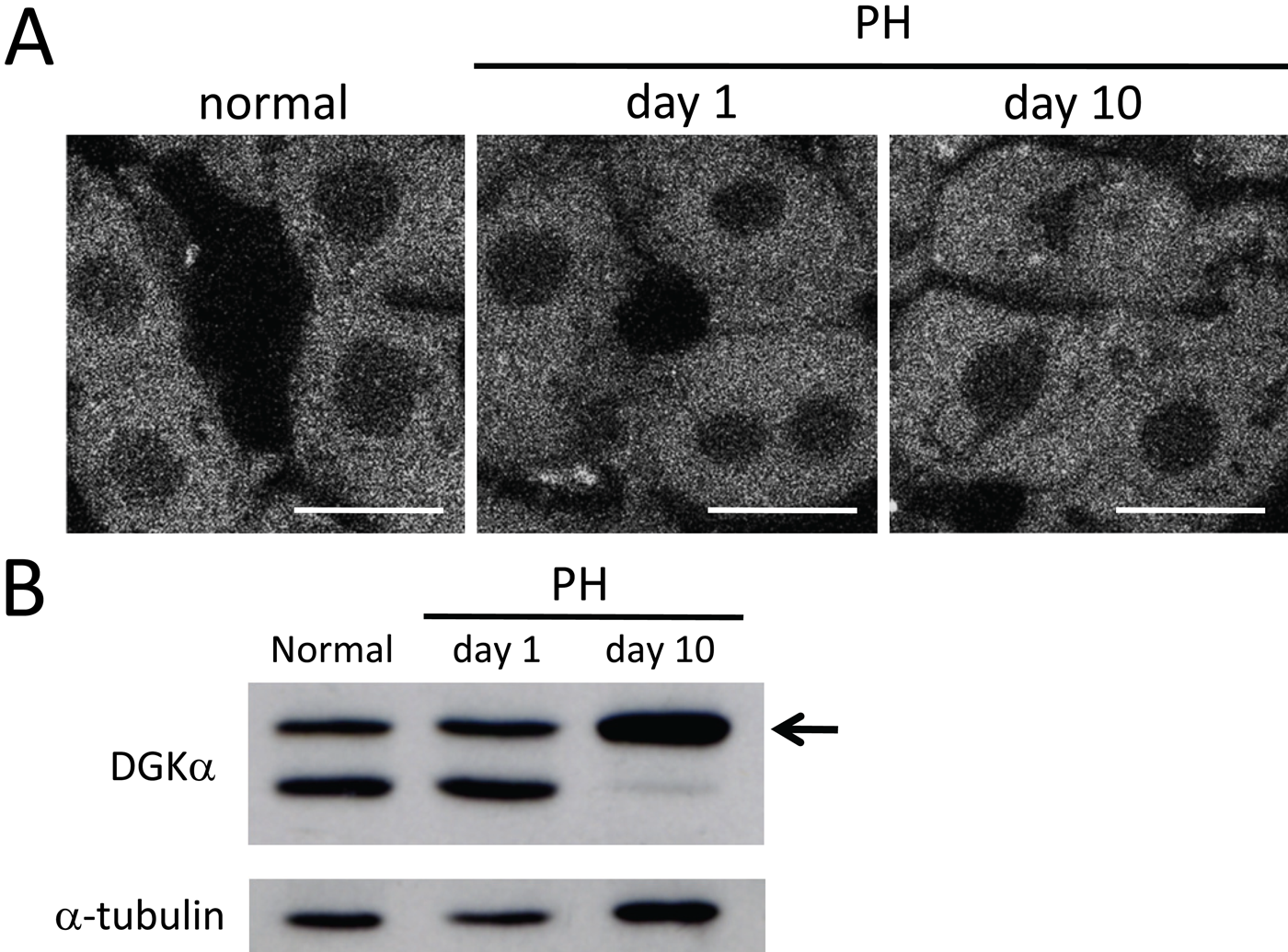
Expression of DGKα in normal and regenerating livers. (A) In immunofluorescence, DGKα-immunoreactivity is detected diffusely in the cytoplasm of all hepatocytes in normal liver. This pattern of expression remains unchanged in regenerating livers at days 1 and 10 after partial hepatectomy (PH). (B) Total proteins (30 µg) were analyzed by immunoblot analysis using anti-DGKα antibody. The antibody recognizes a lower band (70 kDa) in addition to the expected band (80 kDa, arrow) in normal liver and at day 1 after PH. On the other hand, a single band of the expected length is solely recognized at day 10 after PH. Anti–α-tubulin antibody was used as a control. Bars: 20 µm.
Classical PKC (cPKC) is a major downstream molecule under the control of DG. Of cPKCs, PKCα was expressed abundantly in the liver, whereas PKCγ was weak or below detection levels (data not shown). Therefore, we examined whether the expression and localization of PKCα are altered in regenerating liver. In immunohistochemical analysis, PKCα was detected near the plasma membrane of all hepatocytes in normal liver. In the regenerating liver, the immunoreactivity remained unchanged at day 1 after PH, although it was observed more or less clearly on the plasma membrane at day 10 after PH (Fig. 7A). Immunoblot analysis showed that the protein levels for PKCα were largely unchanged in normal and regenerating livers (Fig. 7B).
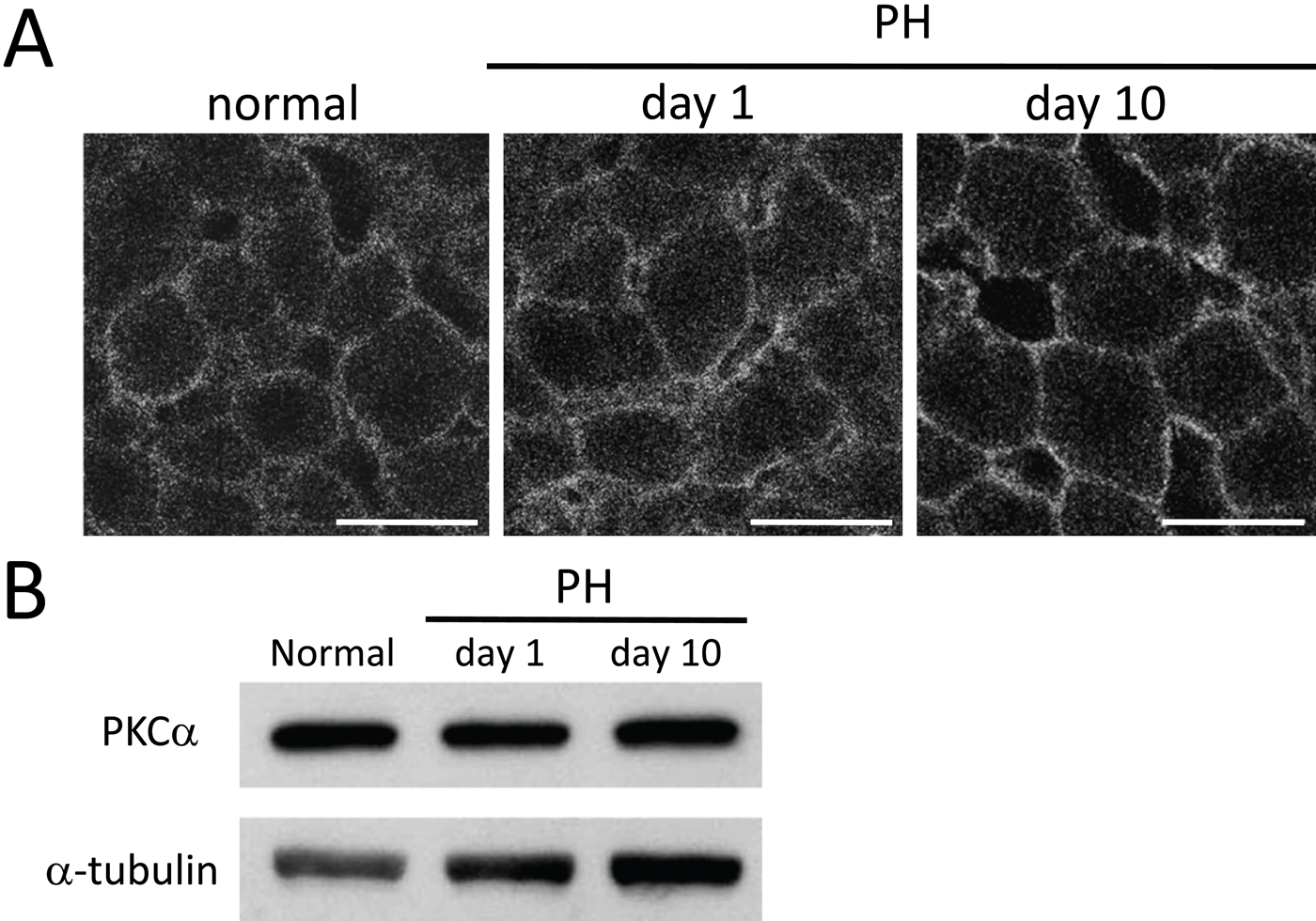
Expression of PKCα in normal and regenerating livers. (A) Immunohistochemically, PKCα is detected near the plasma membrane of all hepatocytes in normal liver. At day 1 after partial hepatectomy (PH), the immunoreactivity remains largely unchanged, whereas it is observed more or less clearly on the plasma membrane at day 10 after PH. (B) Total proteins (30 µg) were analyzed by immunoblot analysis using anti-PKCα antibody. PKCα is largely unchanged at the protein level in normal and regenerating livers. Anti–α-tubulin antibody was used as a control. Bars: 20 µm.
Discussion
In this study, we show that the expression of DGKζ is greatly enhanced in the nucleus of hepatocytes in the process of liver regeneration and returns to the normal level after 30 days of PH. What is the functional significance of increased DGKζ in the nucleus? First, previous reports have described that hepatocytes enter S phase at around day 1 after PH, and the proliferative response after PH is accompanied by upregulation of cell cycle control proteins as well as induction of global protein synthesis (Gottlieb et al. 1964; Michalopoulos and DeFrances 1997). Reportedly, the chromatin contains phospholipids comprising 10% of those present in the nucleus and are closely involved in cell cycle control (Rozengurt et al. 1984; Albi et al. 2004). The initiation of S phase coincides with a transient increase in DG attributable to the hydrolysis of PI, which is accompanied by increased activity of the enzyme PI-PLCβ1 localized to the chromatin (Banfic et al. 1993; Albi et al. 2003). These results of earlier studies suggest the necessity of strict control of the DG level to keep cellular signal transduction within the physiological range. In this regard, it has been shown that DGKζ is associated with chromatin in NIH3T3 cells (Hasegawa et al. 2008) and that it might bind to the retinoblastoma protein (pRB), a tumor suppressor and key regulator of the cell cycle, whose activity depends on the phosphorylation status of pRB (Los et al. 2006). Furthermore, the enhanced activity of DGKζ in the nucleus downregulates phosphorylation at Ser807/811 of pRB in C2C12 cells (Evangelisti et al. 2007). Actually, pRB plays a major role in cell cycle control. Therefore, it is plausible that DGKζ is involved in the control of cell cycle progression through the phosphorylation status of pRB on critical serine residues.
In this regard, this study shows that enhanced expression of DGKζ is observed in the nucleus of hepatocytes in regenerating liver, irrespective of being in S phase or not (Figs. 4 and 5). In addition, DGKζ is expressed stably in the nuclei of primary cultured hepatocytes and hepatoma-derived HepG2 cells, with the former only slightly proliferating and the latter replicating continuously. These data at the tissue and cellular levels suggest that the expression of DGKζ is regulated independently of DNA synthesis in hepatocytes, although the functional relation remains unexplored in this study.
Is DGKζ involved in the transcriptional regulation of hepatocyte growth? Strong evidence exists to suggest that hepatocyte growth factor (HGF) mediates hepatocyte regeneration after liver damage. The level of HGF mRNA in rat liver increases dramatically during 3 to 6 hr after PH, reaches a peak at 12 hr, and returns gradually to undetectable levels by 72 to 96 hr after PH (Zarnegar et al. 1991). In this regard, the time course of DGKζ expression differs greatly from that of HGF mRNA expression, suggesting that DGKζ might not be involved directly in the HGF transcriptional activation.
Then what is the significance of DGKζ in the nucleus? One clue might be gained from our data of an imbalance of the DGKζ protein and mRNA levels. In immunoblot analysis, DGKζ protein is difficult to detect in all cell lysates of the liver and is barely detectable in the nuclear fraction, whereas DGKα protein is easily detected in the total lysate. This observation is unexpected because DGKζ mRNA is expressed more abundantly than DGKα mRNA. This paradoxical pattern of the mRNA and protein expression suggests regulatory mechanisms exerted on the DGKζ protein. It might be that DGKζ is repressed at the translational level or that it is degraded after translation under normal conditions. In this respect, regulation of hypoxia-inducible factor (HIF) 1α expression is well known to exemplify the discrepant expression of mRNA and protein. The HIF1α gene is transcribed and translated continuously under normoxic conditions, although HIF1α protein itself is detected at a very low level because of its rapid breakdown through the ubiquitin-proteasome pathway (Tanimoto et al. 2000). The benefit of a regulatory system of this kind, despite its apparent wastefulness, is its rapid response to emergent stressful conditions (i.e., oxygen deprivation in the case of HIF1α). Therefore, it can be inferred that the same regulatory system is also applicable to DGKζ in the liver, although no data are available. Hepatectomy is regarded as being life-threatening. Therefore, a rapid increase in DGKζ in the hepatocyte nucleus might be regarded as a sort of response to emergent stressful conditions. In a hepatectomized liver, protein synthesis is greatly enhanced to restore the original size and to meet the metabolic demand (Goggin et al. 2004). Considering our data showing that DGKζ protein expression remains upregulated at day 10, when regeneration is all done and returns to the normal level at day 30, it is suggested that DGKζ-protein levels may reversibly change under various conditions, such as the metabolic demand. In this regard, the mammalian target of rapamycin (mTOR), a highly conserved serine/threonine kinase, plays a central role in regulating cell growth through the control of protein synthesis. In growth-promoting conditions, mTOR sustains translation by phosphorylating the eukaryotic initiation factor (eIF) 4E-binding proteins and ribosomal protein S6 kinases (S6Ks) and activating the phosphorylation of eIF4G (Gingras et al. 2001). Intriguingly, mTOR-dependent phosphorylation of p70S6 kinase, one isoform of S6Ks, reportedly requires mTOR binding to PA that is produced by DGKζ, suggesting that DGKζ controls mTOR activity directly (Avila-Flores et al. 2005). Therefore, further study is warranted to address whether increased DGKζ contributes to upregulated protein synthesis through mTOR signaling in regenerating liver.
With regard to DGKα, results show that the DGKα-immunoreactive band pattern in immunoblot analysis is changed between normal and regenerating livers. As reported previously, two bands, the expected (80 kDa) and the lower (70 kDa) bands, are recognized with anti-DGKα antibody in normal liver (Goto et al. 1992). In addition, AKI human melanoma cells show two DGKα-immunoreactive bands, each of which is markedly decreased by siRNA specifically targeted against DGKα mRNA (Yanagisawa et al. 2007). It is therefore suggested that the lower band represents another form of DGKα protein and that it is probably generated by proteolysis or alternative splicing. The antibody used in the present study is produced against the N-terminus of DGKα (aa. 2–357). Therefore, the lower species might lack a C-terminus portion that covers a part of the catalytic domain (Goto et al. 1992). If this is the case, the lower species might exhibit reduced or no enzymatic activity, suggesting a sort of dominant-negative effect of DGKα activity in normal liver. In addition, higher DGKα activity might be gained in the cytoplasm of hepatocytes in a regenerating liver, which might reflect an elevated DG level following partial hepatectomy (Bocckino et al. 1989).
In summary, this study revealed an increased expression level of DGKζ protein in the nucleus and altered expression of DGKα protein in the cytoplasm of hepatocytes in regenerating liver. These findings suggest that DGKζ and DGKα are involved, respectively, in the nucleus and the cytoplasm of hepatocytes of regenerating liver. Further studies are necessary to clarify the roles played by these DGKs in the cellular processes of liver regeneration.
Footnotes
The authors declared no potential conflicts of interest with respect to the authorship and/or publication of this article.
The authors disclosed receipt of the following financial support for the research and/or authorship of this article: This work was supported by Grants-in-Aid from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) of Japan (TN, KG).