
Abstract
The pig is becoming an increasingly used non-primate model in experimental studies of human retinal diseases and disorders. The anatomy, size, and vasculature of the porcine eye and retina closely resemble their human counterparts, which allows for application of standard instrumentation and diagnostics used in the clinic. Despite many reports that demonstrate immunohistochemistry as a useful method for exploring neuropathologi-cal changes in the mammalian central nervous system, including the pig, the porcine retina has been sparsely described. Hence, to facilitate further immunohistochemical analysis of the porcine retina, we report on the successful use of a battery of antibodies for staining of paraformaldehyde-fixed cryosectioned retina. The following antibodies were evaluated for neuronal cells and structures: recoverin (cones and rods), Rho4D2 (rods), transducin-γ (cones), ROM-1 (photoreceptor outer segments), calbindin (horizontal cells), PKC-α (bipolar cells), parvalbumin (amacrine and displaced amacrine cells), and NeuN (ganglion cells and displaced amacrines). For detecting synaptic connections in fiber layers, we used an antibody against synaptobrevin. For detecting retinal pigment epithelium, we studied antibodies against cytokeratin and RPE65, respectively. The glial cell markers used were bFGF (Müller cells and displaced amacrine cells), GFAP (Müller cells and astrocytes), and vimentin (Müller cells). Each staining effect was evaluated with regard to its specificity, sensitivity, and repro-ducibility in the identification of individual cells, specific cell structures, and fiber layers, respectively. The markers parvalbumin and ROM-1 were tested here for the first time for the porcine retina. All antibodies tested resulted in specific staining of high quality. In conclusion, all immunohistochemical protocols presented here will be applicable in fixed, cryosectioned pig retina.
Keywords
T
Today a range of in vitro and in vivo porcine models with pathophysiological hallmarks of human retinal disorders are available (Chader 2002). For example, glaucoma models characterized by a loss of ganglion cells have been established, using acute (Kyhn et al. 2009b) or chronic elevated intraocular pressure (Ruiz-Ederra et al. 2005). A transgenic model of the inherited disease retinitis pigmentosa, characterized by a loss of photoreceptors, has been established (Petters et al. 1997; Li et al. 1998). Also, a transgenic pig with expression of green fluorescent protein has been produced and used for isolation of retinal stem cells, which have subsequently been used in transplantation studies (Park et al. 2001; Klassen et al. 2008). Retinal conditions such as retinal detachment (Scholda et al. 1999) and proliferative vitreoretinopathy (García-Layana et al. 1997) can also be modeled. The possibility of making organotypic cultures of embryonic and adult pig retina offers an experimental tool for many approaches for retinal research (Gaudin et al. 1996; Luo et al. 2001; García et al. 2002; Winkler et al. 2002; Engelsberg et al. 2005; Kaempf et al. 2008).
Studies of retinal development as well as evaluation of neuropathological changes in the diseased retina are preferably based on histological and immuno-histochemical methods. For the pig retina, some cell type–specific antibodies have already been used in retinogenesis, descriptive, and neuropathological studies (Luo et al. 2001; García et al. 2002; Ghosh and Arnér 2002; Hendrickson and Hicks 2002; Yang et al. 2002; Ghosh et al. 2004,2007; Ruiz-Ederra et al. 2004; Warfvinge et al. 2005,2006; Lee et al. 2006b; Klassen et al. 2007,2008; Ahn et al. 2009; Guduric-Fuchs et al. 2009). In most of the previous reports, the focus is on one or a few retinal cell types and often one developmental stage. However, in the very recent report by Guduric-Fuchs et al. (2009), a range of markers for different developmental stages of pig retina was explored.
The aim of the present study was to contribute to present knowledge by extending the range of markers specific for neuronal and glial cells in the adult normal pig retina. Danish domestic mixed breed (Danish Landrace/Duroc/Hampshire/Yorkshire) pigs were utilized for evaluating the specificity of different mono-and polyclonal antibodies, using fixed and cryosectioned tissue. The following antibodies were evaluated for neuronal and glial cells, respectively: recoverin (cones and rods), Rho4D2 (rods), transducin-γ (cones), calbindin (horizontal cells), protein kinase C-α (PKC-α) (bipolar cells), parvalbumin (amacrine cells), NeuN (ganglion cells, displaced amacrines), basic fibroblast growth factor (bFGF) (Müller cells, amacrine, and displaced amacrine cells), glial fibrillary acidic factor (GFAP) (Müller cells and astrocytes), and vimentin (Müller cells). An antibody against ROM-1 for detection of photoreceptor outer segments was also studied. For visualizing synaptic layers, an antibody against synaptobrevin was used. For identification of retinal pigment epithelium (RPE), antibodies against cytokeratin and RPE65, respectively, were used. For the first time, parvalbumin and ROM-1 were tested in this study for porcine retina.
At least 3 but to up to 15 specimens were used per antibody examined. Each staining effect was evaluated with respect to specificity and sensitivity in identification of individual cell types and structures.
Material and Methods
Animals
Fifteen female pigs of randomly mixed breed, Danish Landrace/Duroc/Hampshire/Yorkshire, were used in this study. Pigs were obtained from a specific-pathogen-free local Danish farm and were housed at the Eye Pathology Section, Institute for Neuroscience and Pharmacology, University of Copenhagen, Denmark. After eyes were enucleated under anesthesia (see details below), further tissue processing was conducted at the Department of Ophthalmology, Lund University, Lund, Sweden. The age and weight of the pigs used were ×4–5 months and 30 kg, respectively. In the stables, pigs were kept under a 14-hr light/10-hr dark cycle and at a room temperature of 20C. The pigs were group housed in a secluded underground stable in concrete pens (1.50 m × 3 m) and provided with straw. The animals were given toys for behavioral enrichment. A commercial diet of food pellets for slaughter pigs (Swine-Lux; Altromin GmbH and Co., Lage, Germany) and additional vegetables were given. The research protocol was approved by the Danish Animals Experiment Inspectorate and was conducted in accordance with the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research.
Tissue Processing
Normal eyes were enucleated with the animal under anesthesia consisting of an intramuscular injection of 15 mg of midazolam (Dormicom A; Roche, Hvodovre, Denmark) and a combination of zolazepam hydro-chloride (11.9 mg/ml; Zoletil 50 Vet; Virbac SA, Carros, France) mixed with xylazine hydrochloride (12.38 mg/ml; Intervet, Skovlunde, Denmark), ketamine hydrochloride (14.29 mg/ml; Intervet), and methadone (2.38 mg/ml; Nycomed, Roskilde, Denmark). After enucleation, pigs were killed by an intravenous injection of 2 to 4 g of pentobarbital sodium (200 mg/ml; KVL, Copenhagen, Denmark). Globes were placed in 4% paraformalde-hyde (PFA) for a few minutes. The anterior segment, the lens, and most of the vitreous were then removed, and the posterior segment was postfixed for 2 hr in 4% PFA, with subsequent rinsing in increasing concentrations of sucrose diluted in 0.1 M phosphate buffer. A horizontal cut was made that extended from the temporal retinal margin to 2 to 3 mm nasal to the optic disc, resulting in a sample composed of the temporal ciliary margin, the area centralis, and the optic disc. The sections selected for this study included the area centralis in the pig, which is enriched in cones and therefore comparable to the human fovea (Hendrickson and Hicks 2002). The tissues were then embedded in a gelatin medium (30% egg albumin and 3% gelatin in distilled water), sectioned into 12-μm slices with a cryostat, and stored at −20C until further processing.
Immunohistochemistry
Table 1 summarizes the antibodies used, the final optimized concentrations recommended, and the sources for antibodies. Hematoxylin–eosin (H&E) was used for routine staining. In brief, for immunohistochemical staining, retinal sections were dried in room temperature for at least 30 minutes, rinsed in 0.1 M PBS, and thereafter exposed to primary antisera diluted in 0.1 M PBS containing 0.25% Triton X-100 (PBST) and 1% BSA in a moist chamber for 16 to 18 hr at 4C. Thereafter, sections were rinsed in PBST and incubated in PBST and 1% BSA with appropriate FITC- and Texas Red–conjugated secondary antibodies (1:200 dilution; Jackson Immunoresearch, West Grove, PA). Negative controls with the primary antisera omitted were used as controls.
Microscopy Analysis
Specimens were examined using light and epifluorescence microscopy (Nikon; Europe BV, Amsterdam, The Netherlands). Pictures were taken using a Nikon camera.
Results
Histological Results
A detailed description of the antibodies investigated, including cell type or structure recognized, species in which the respective antibodies were produced, dilutions, and sources is shown in Table 1.
We chose to use immersion fixation of the dissected whole porcine retina with incubation in PFA for 2 hr, since this method, used in our previous experiments, successfully rendered satisfactory and consistent immunohistochemical staining results with all parts of the retina.
Histological evaluations of gross morphology using H&E-stained material revealed that in all 15 untreated porcine retinas, the layering of both the nuclear and the synaptic layers was normal. This indicated that all retinas were healthy and could be further used (Figures 1A and 1A'). Three to 15 specimens were included in the studies of the respective antibodies. All antibodies were applied in at least three independent staining sessions to validate the reproducibility of the staining results. Furthermore, each staining effect was evaluated with respect to specificity and sensitivity in the identification and visualization of the respective cell type or structure.
Overall, all antibodies studied specifically visualized the respective cell type or structure they have been reported to identify, in agreement with previous reports that used porcine or non-porcine tissue from the retina. In addition, no notable variability was observed for the localization and distribution of the respective antibody staining pattern among the specimens. If any unspecific staining related to the secondary antibody was found, it was associated mainly with the blood vessels of the choroid. No quantitative measurements were attempted in this study. Therefore, all of the descriptions are of a qualitative nature but can indeed serve as a valuable source of information for more in-depth studies of detailed quantifications using, e.g., stereological cell counting. The photomicrographs presented for all the immunostained samples are from the mid periphery of the retina.
Primary antibodies used for immunohistochemistry
Gift from Dr. A. Dizhoor, Detroit, MI.
Gift from Dr. R.S. Molday, Vancouver, BC, Canada.
Gift from Dr. A. Thompson, Ann Arbor, MI. mo, mouse; ra, rabbit.

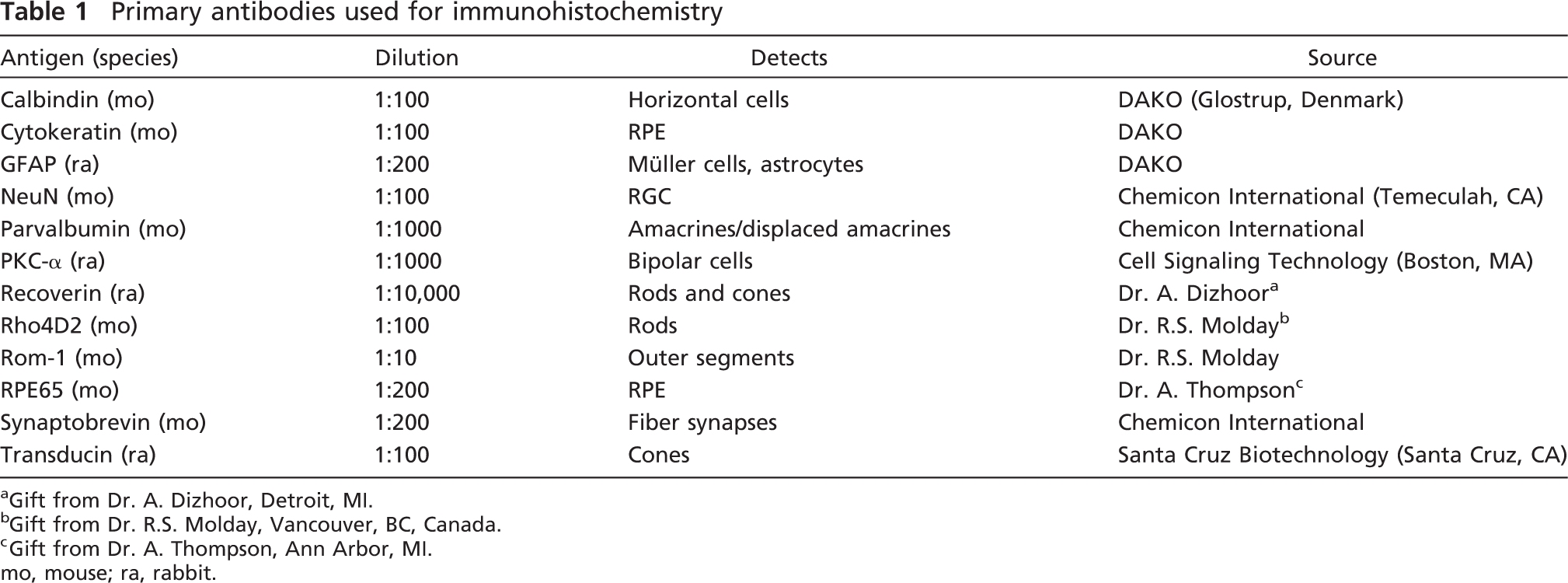
Markers for Photoreceptors
The photoreceptors together with photosensitive retinal ganglion cells (RGCs) are responsible for converting light into electric signals, a process called phototransduction. The photoreceptors, divided into rods and cones as defined by their morphology and functional photopigments used, are localized in the outer part of the retina (Figure 2). Their nuclei are found in the outer nuclear layer (ONL), and their inner and outer segments are oriented toward the RPE. The lining between the nuclei and the segment layers are called the outer limiting membrane (OLM). The synapses between photoreceptors and bipolar cells and horizontal cells, respectively, occur in the outer plexiform layer (OPL).
Here, three different antibodies, recoverin (rods and cones), Rho4D2 (rods), and transducin (cones), were used to detect the photoreceptors (Figures 2A–2C'). Use of all three antibodies resulted in staining specifically associated with photoreceptors.
The antibody against recoverin, a Ca2+-binding protein (Dizhoor et al. 1996), detected the cellular parts of the rods and cones placed in the ONL (Figures 2A and 2A'). However, the staining of recoverin-labeled cones, identified by the location of their nuclei close to the scleral side of the ONL, was more intense than the fraction of labeled rods (Figures 2A and 2A’ to 2B and 2B', cf. with Rho4D2 staining). Also, in a small fraction of the cells, the inner segments of the photoreceptor were visualized (Figures 2A and 2A', inset).
Rods were specifically detected using an antibody against Rho4D2, which recognizes the rod-specific pigment rhodopsin and, more specifically, its N-terminal region (Hicks and Molday 1986). The labeling visualized primarily the lining of the cell bodies at the location of their nuclei in the ONL (Figures 2B and 2B'). In addition, dense Rho4D2-positive labeling was found in the rod outer segments (Figures 2B and 2B', inset).
Cones and rods were identified using fluorescence immunohistochemistry with antibodies against recoverin, Rho4D2, transducin, and ROM-1.
The population of cones was identified using an antibody against transducin, a G protein coupled to the photopsins, the cone-specific pigments (Figures 2C and 2C'). There are cone- and rod-specific forms of transducin, and it is activated by the conformational change that occurs when light is absorbed by the respective photopigment molecule (Conklin and Bourne 1993). Transducin immunolabeling was most intense in the inner segments of the cones. In addition, transducin expression was noted in some of the cone outer segments (Figures 2C and 2C', inset). Cone nuclei located close to the OLM displayed a weak transducin expression. The cones’ synaptic terminals in the OPL showed strong expression of transducin.
Outer segments of photoreceptors, cones and rods, were detected using the novel marker ROM-1 (Figure 2D, inset). ROM-1 encodes an integral membrane protein found in the photoreceptor disk rim of the eye and is essential for disk morphogenesis and may also function as an adhesion molecule involved in the stabilization and compaction of outer segment disks or in the maintenance of the curvature of the rim (Lee et al. 2006a).
Markers for Bipolar, Horizontal, Amacrine, and Ganglion Cells
The direct pathway of information flow from the photoreceptors to the optic nerve is a three-neuron chain: photoreceptor to bipolar cell to ganglion cell. The processes of the horizontal cells enable lateral interaction between photoreceptors and bipolar cells, whereas amacrine cells enable lateral interaction between bipolar cells and RGCs. The axons of the RGCs form the optic nerve that leaves the retina and carry visual information to the visual cortex.
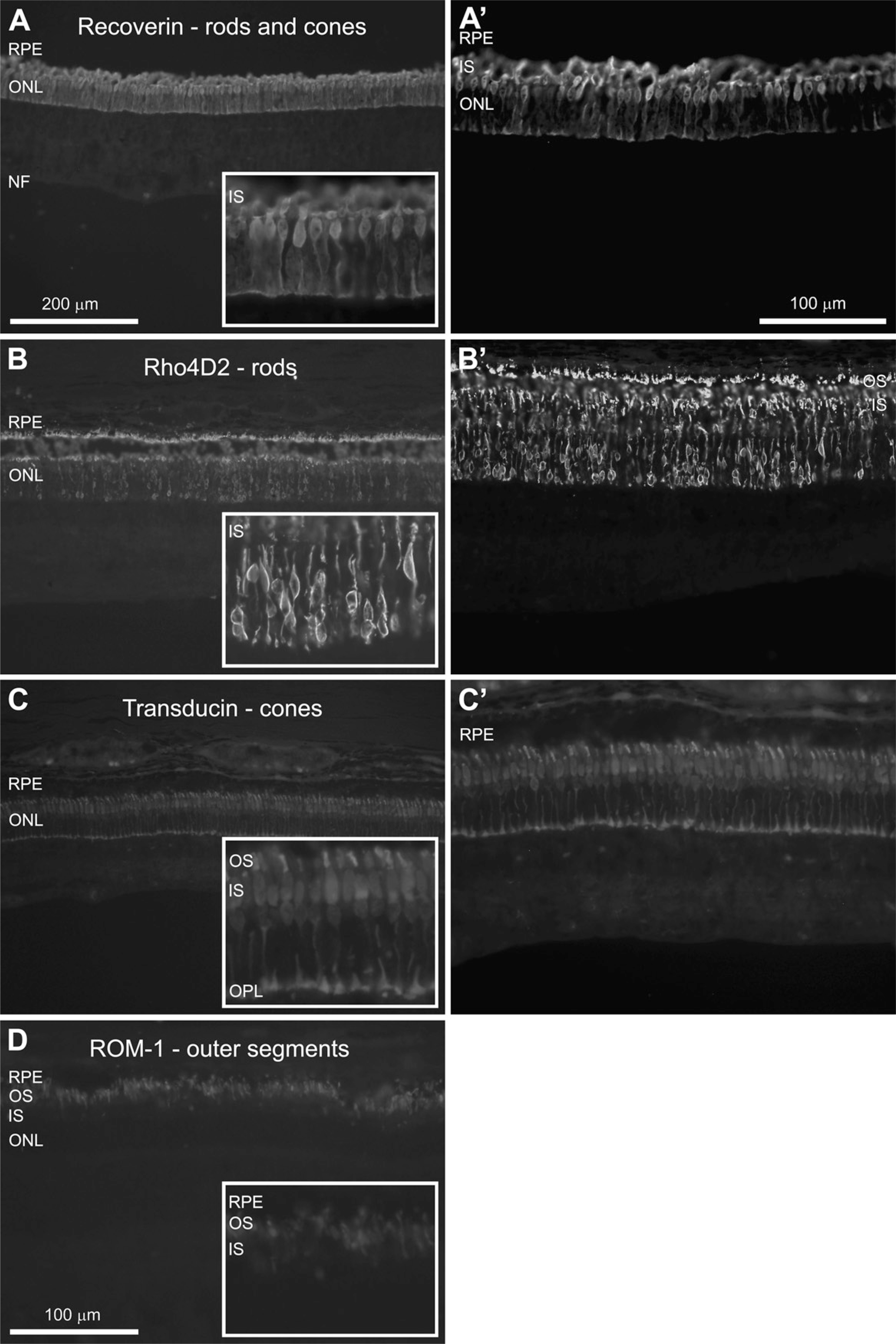
In this study, bipolar, horizontal, amacrine, and RGCs were detected using antibodies against PKC-α, calbindin, parvalbumin, and NeuN, respectively (Figure 3). Evaluation of all four antibodies revealed labeling specifically associated with the appropriate neuronal cell type.
Horizontal cells, with their cell bodies in the inner nuclear layer (INL) and processes localized in the OPL, were detected using an antibody against calbindin (Figures 3A and 3A'). Calbindin is a Ca2+-binding protein and was specifically expressed by the horizontal cells, identified by their cellular morphology and location (Hamano et al. 1990). The staining of the whole-cell cytoplasm, including the nucleus and the extending processes, allowed for visualization of the orientation of the processes in the OPL (Figures 3A and 3A').
Bipolar cells, with their cell bodies located in the INL and synapses in the OPL and IPL, were detected using an antibody against PKC-α. PKC-α is involved in cell signaling cascades, and its expression was most intense at the synapses of the bipolar cells, i.e., in the OPL and IPL (Figures 3B and 3B') (Wood et al. 1997). The nuclei of the cells as well as the long extending processes could also be detected (Figure 3B').
Amacrine cells, whose bodies are also confined to the INL while their processes are in the IPL, were detected here for the first time in the pig retina by using an antibody against parvalbumin (Figures 3C and 3C'). Parvalbumin is another Ca2+-binding protein (Hamano et al. 1990), and its expression was restricted to amacrine cells in the INL and some displaced amacrine cells found in the ganglion cell layer (GCL). In addition, two fiber layers in the central part of the IPL stained positive for parvalbumin. The location of these layers suggests that they are composed of amacrine cell processes.
In the GCL, an antibody against NeuN was used to visualize mainly RGCs (Figures 3D and 3D'). NeuN has been previously observed to label neuronal nucleus protein in a variety of neuronal subtypes (Mullen et al. 1992; Sarnat et al. 1998). However, this marker also detected weakly labeled displaced amacrine cells. NeuN expression was found in the nuclei and cytoplasm of stained cells, displaying nuclei that were more strongly labeled than cytoplasm. RGCs with different sizes of cell bodies were observed with this stain. In addition, the nucleoli of the cells could be visualized.
Fluorescence immunostaining of the major interneurons of the retina, i.e., horizontal, bipolar, and amacrine cells. In addition to ganglion cell labeling, the output neurons of the retina are shown.
Marker for Synapses in OPL and IPL: Synaptobrevin
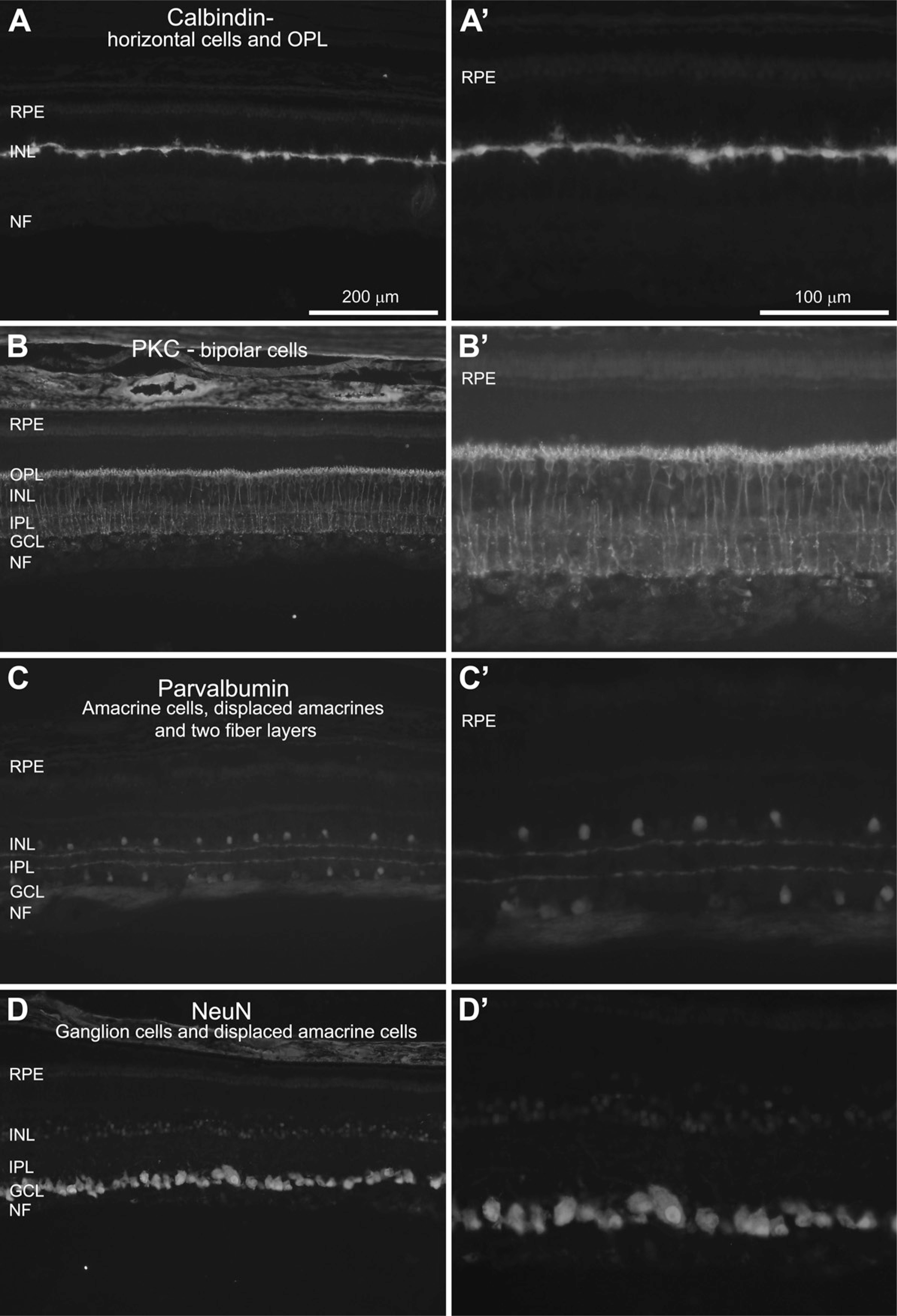
The fiber layers of the retina that consist of synaptic connections, i.e., the OPL and the IPL, were identified using the marker synaptobrevin (Figure 4A). Synaptobrevin, also called vesicle-associated membrane protein (VAMP), is a synaptic vesicle membrane protein with a central role in neurotransmission (Linberg et al. 2006). The OPL consists of a dense network of synapses between dendrites of horizontal and bipolar cells from the INL and processes from photoreceptors from the ONL. The IPL consists of bipolar and amacrine cell synapses with RGCs and could be clearly visualized here. Here, the synaptobrevin staining revealed two fiber layers in the IPL and one fiber layer in the OPL.
The RPE: RPE65 and Cytokeratin
The RPE consists of one layer of epithelial cells responsible for removal of photoreceptor discs. Here, the RPE was identified by using antibodies against RPE65 (Iseli et al. 2002) and cytokeratin (Hiscott et al. 1984; Schraermeyer et al. 1997). RPE65 has been well described as playing a critical role in the visual cycle by being implicated in the metabolism of vitamin A, a precursor for rhodopsin. Cytokeratin is an intermediate filament present in epithelial cells (Page 1989). Both markers clearly detected the chain-like pattern of cells in the RPE. (Figures 4B and 4C).
Glial-specific Markers GFAP, Vimentin, and bFGF
The major glial cells of the porcine retina are Müller cells, astrocytes, and microglia. The Müller cell is the principal glial cell of the retina and has its cell body situated in the INL and processes stretching from the outer to the inner limiting membrane. Müller cells serve as architectural supports for the retina, stretching their processes across the entire retina and insulating the neuronal cells from electrical and chemical signals from nearby neurons by ensheathing them. These cells also play a critical role in maintaining overall homeostasis in the retina and thus maintaining survival and viability of the neurons. Astrocytes, the other group of glial cells, have their cell bodies and processes in the nerve fiber layer (NFL). Astrocytes have functions similar to those of the Müller cells in the retina.
Müller cell nuclei were visualized using an antibody against bFGF (Li et al. 1998), which intensely stained nuclei in the INL (Figure 5A). bFGF is also known to be expressed by displaced amacrine cells in the GCL, which was also found here.
Application of antibodies against the intermediate filaments, i.e., GFAP and vimentin, resulted in detection of Müller cell processes (Lewis and Fisher 2003) (Figures 5B and 5C). Both of the markers visualized Müller cell fibers reaching from the NFL into the outer retinal layers. However, stronger labeling of the fibers was found with the vimentin staining, whereas GFAP expression was stronger in the Müller cell end-feet situated in the NFL (cf. Figures 5B and 5C). GFAP also labeled astrocytes situated in the NFL (Figure 5B). GFAP-labeled astrocytes were often found associated with blood vessels and displayed small cell bodies and multiple short processes.
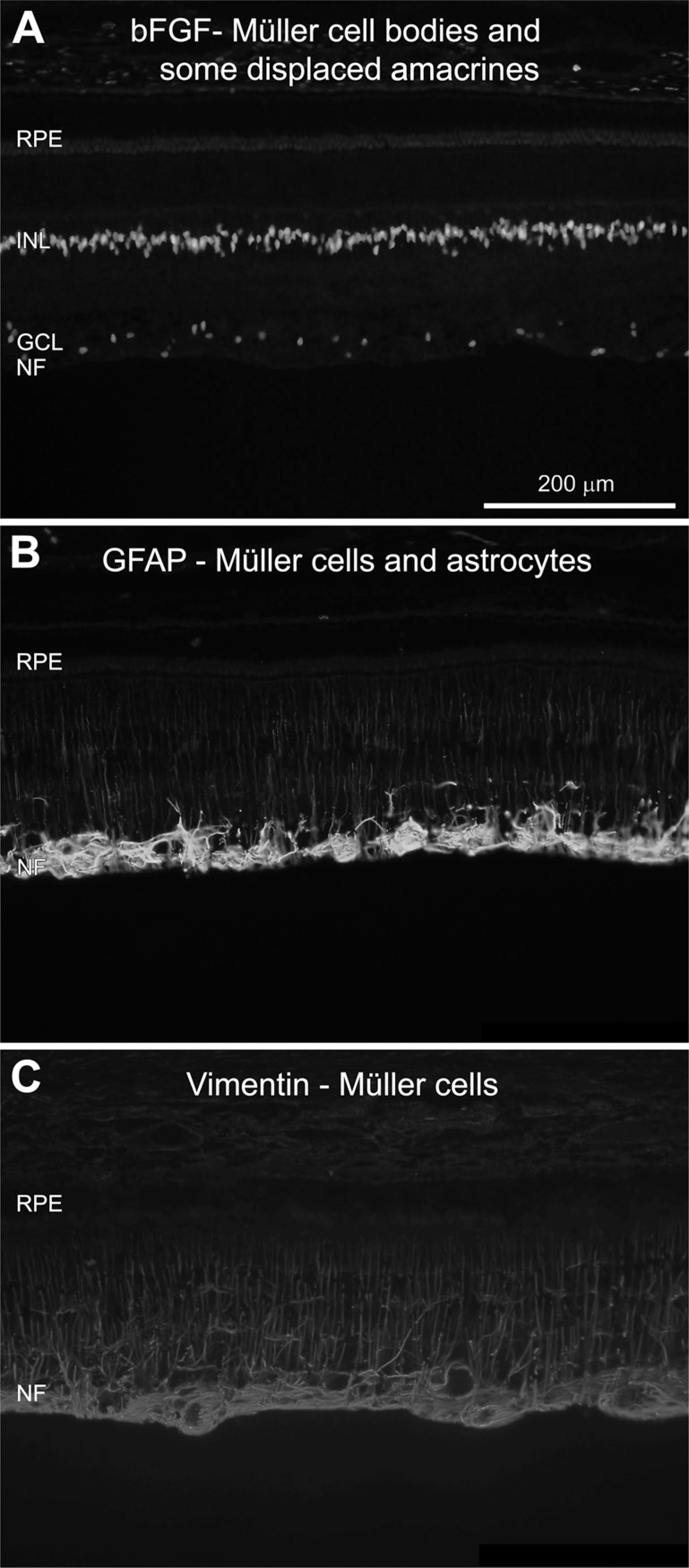
Fluorescent immunolabeling for retinal glial cells is shown.
Discussion
Here we present a range of robust retinal cell- and structure-specific antibodies, including the novel markers parvalbumin and ROM-1, which were used for immunohistochemical analysis of the normal adult pig eye. Overall, our staining results confirm those of previous reports, when present, using the current antibodies. The current data concern the morphology and location of the immunostained cells and structures in the mid periphery of the pig retina. Specific and stable immunolabeling was found for all 14 antibodies explored by using PFA fixation, retinal sections processed by a cryostat, and the presently described staining protocols. No variations in staining intensity and pattern were noted among the specimens included, i.e., 3 to 15 specimens per antibody evaluated.
Fixation Method and Tissue Preparation
An ideal fixation method and tissue preparation are most important when assessing specimens based on histological and immunohistochemical analysis. For the current antibodies, fixation with PFA was preferred. PFA diffuses through the tissue and inhibits degradation of the tissue by inducing covalent alterations and cross-binding of the tissue molecules, preserving to some extent the globular form of the proteins (Ikeda et al. 1998; Montero 2003). Here, eyes were placed in PFA to stabilize the tissues before the anterior segment, the lens, and the vitreous were removed. Then, the posterior segment was fixed by immersion for 2 hr. This method results in a thorough and even fixation and was chosen based on our recent successful his-tological results (Warfvinge et al. 2005,2006; Klassen et al. 2007,2008). The specimen preparation method also plays a role in the histochemical properties of the retinal tissue. The only method used here was sectioning into 12-μm-thick sections, using a cryostat. It is known that different processing techniques can alter the antigen availability and final morphology due to pretreatment (Lyck et al. 2006). Here, PFA immersion fixation followed by tissue processing and cryostat sectioning worked very well for the 14 antibodies investigated. The H&E-stained material revealed a well-preserved retinal layering as well as cellular profiles, further proving the adequacy of the chosen methods.
Markers for Photoreceptors Recoverin, Rho4D2, Transducin, and ROM-1
The staining results for the photoreceptor markers recoverin (rods and cones), Rho4D2 (rods), and transducin (cones) were in accordance with our recent reports and those of others. Here, recoverin visualized rods and cones. In the pig, recoverin has been widely used as a marker in studies of retinogenesis and photo-receptor degeneration (Petters et al. 1997; Li et al. 1998; Chandler et al. 1999; Traverso et al. 2003; Engelsberg et al. 2005; Engelsberg and Ghosh 2007; Gu et al. 2007; Klassen et al. 2007,2008). Rods were specifically detected by using an antibody against rhodopsin, i.e., Rho4D2, and the staining result was in parallel with that of the original description of the staining pattern (Hicks and Molday 1986) and with experimental in vitro and in vivo porcine studies performed by us and others (Gaudin et al. 1996; Warfvinge et al. 2005; Ghosh et al. 2007; Klassen et al. 2007,2008). The transducin-staining pattern was in agreement with those of recent studies where the marker was used as a parameter in phenotypic differentiation in retinal development, as well as transplantation studies (Mahmoud et al. 2003; Engelsberg et al. 2005; Engelsberg and Ghosh 2007; Ghosh et al. 2007; Klassen et al. 2007,2008). Photoreceptor outer segments were identified using the novel marker ROM-1. This marker was useful in grafting studies for exploring the level of differentiation of transplanted cells (Warfvinge K, unpublished data).
NeuN, a Marker for Neurons in the GCL
Previously, RGCs have been detected primarily by using retrograde labeling with fluorogold (FG) in vivo, which is injected into the optic nerve (Ruiz-Ederra et al. 2005; Lalonde et al. 2006). This method provides specific labeling of the RGCs in the GCL, but it puts an extra burden on the animal as it has to be exposed to surgery. Here the neurons in the GCL were labeled using an antibody against NeuN. The NeuN expression pattern was in accordance with that of the original literature describing NeuN (Mullen et al. 1992; Sarnat et al. 1998). As a nuclear marker, NeuN was useful for quantifying numbers of neurons remaining in the GCL in a pig model of acute retinal ischemia (Kyhn et al. 2009a). NeuN may also label displaced amacrine cells in the GCL. To further characterize NeuN as a marker exclusively for ganglion cells, a double-labeling study with FG or double labeling with an antibody that specifically reacts against displaced amacrine cells, such as AP2α, could be conducted (Guduric-Fuchs et al. 2009). Brn3a, a POU domain transcription factor, a marker successfully used to label RGCs in the adult rat retina (Nadal-Nicolás et al. 2009), was also recently reported to be expressed in adult porcine RGCs (Guduric-Fuchs et al. 2009). Brn3a may thus serve as a specific marker for RGCs in experimental studies.
Cellular Markers for INL Horizontal Cells (Calbindin), Bipolar Cells (PKC-α), and Amacrine Cells (Parvalbumin)
Horizontal cells, identified by their normal cell morphology and location in the INL, were detected using an antibody against calbindin. The calbindin expression pattern was in parallel with that found using other species (Hamano et al. 1990; Pasteels et al. 1990). However, Guduric-Fuchs et al. (2009), using a calbindin antibody different from the oneweused, found immunostaining in cone, horizontal, amacrine, and ganglion cells in the adult pig retina. This may be explained by a different specificity for the respective antibodies. Calbindin expression has also been used in in vitro porcine studies for studying neuronal differentiation (Gu et al. 2007). The expression pattern of PKC-α revealed that normal bipolar cell profiles in the INL corresponded well to those of other studies using the pig retina (Ghosh et al. 2007). Expression of PKC-α clearly displays the entire cell morphology, making it appropriate for studying changes in retinal development and cytoarchitecture (Ghosh et al. 2007; Guduric-Fuchs et al. 2009). PKC-α expression has been used as a marker for phenotypic differentiation in porcine retinal cell cultures (Klassen et al. 2007,2008). For the first time here, we evaluated a parvalbumin antibody for detection of amacrine cells and found, indeed, the staining showed expression restricted to amacrine cells in the INL, some displaced amacrines in the GCL, and two fiber layers in the central part of the IPL. The location of these layers suggests that they are composed of amacrine cell processes. In agreement, parvalbumin expression was found in amacrine and displaced amacrine cells in chicken, rat, and dog (Hamano et al. 1990).
Synaptobrevin, a Marker for Synapses in Fiber Layers OPL and IPL
The synaptobrevin staining revealed two fiber layers in the IPL and one fiber layer in the OPL. These and related synaptic proteins have been characterized in depth in lower species, such as Brazilian opossum and rodents (Greenlee et al. 2001; Sherry et al. 2003). The current staining results are in parallel with those of our recent report using synaptobrevin to phenotypically characterize allogenically grafted porcine cells (Klassen et al. 2008).
Markers for the RPE: RPE65 and Cytokeratin
Similar staining patterns of the closely attached RPE cells were produced by using antibodies against RPE65 and cytokeratin, respectively, which are in parallel with those of our recent staining results (Warfvinge et al. 2005). Cytokeratin expression has been previously used to detect the RPE in pig transplantation studies (Del Priore et al. 2004).
Specificity of Glial Antibodies bFGF, GFAP, and Vimentin
The staining pattern of bFGF is in line with the description by Li et al. (1998), with expression found in Müller and displaced amacrine cell nuclei. The level of GFAP expression is commonly used as a hallmark for gliosis induced by disease or injury in models of retinal degeneration (Bignami and Dahl 1979). Müller cells can upregulate GFAP and vimentin in porcine models of retinal detachment, acute retinal ischemia, and retinitis pigmentosa, respectively (Li et al. 1998; Iandiev et al. 2006; Ghosh et al. 2007; and unpublished data). The overall staining pattern of GFAP is similar to that reported in recent studies using the adult pig (Kiilgaard et al. 2002; Warfvinge et al. 2005; Iandiev et al. 2006; Ghosh et al. 2007). Glial differentiation determined by vimentin and GFAP expression levels was studied in in vitro studies of porcine retinal progenitors and after grafting of these tissues in vivo (García et al. 2002; Gu et al. 2007; Klassen et al. 2007,2008).
Adding Markers to the Histology Tool Box for Future Retinal Research
Here we demonstrate robust immunostaining results for 14 retinal markers, to be added to the histology tool box for future phenotypic investigations of the porcine retina. We also present two novel markers, parvalbumin and ROM-1, for detection of amacrine cells and photoreceptor outer segments, respectively. The majority of the 14 markers are directly related to the main pathways of the visual transmission system. This makes them most relevant in histological investigations in a broad range of retinal research where the pig is used as a model, from mapping of retinal development to studies of retinal dysfunction caused by, e.g., neurodegeneration (age-related macular degeneration, glaucoma, and retinitis pigmentosa) or vascular disorders such as revascularization and thrombosis. In addition, drug toxicity and the efficacy of refinement of retinal surgery, such as laser treatment, can be assessed using a histological outcome. Since the pig eye and retina closely resemble their human counterparts, the pig will most likely continue to serve as an invaluable source for retinal research in many aspects.
Footnotes
Acknowledgements
This work was supported by Lund University and the Medical Faculty at Lund University (UEJ), the Crown Princess Margareta's Foundation for the vision disabled (UEJ and KW), the Gail and Richard Siegal Foundation (KW), and the Swedish Research Council (medicine) (KW).
For housing of the pigs, we thank the staff at Eye Pathology Section, Institute for Neuroscience and Pharmacology, University of Copenhagen, Denmark. Thanks to Dr. Per Ekström for valuable comments on the manuscript.