
Abstract
A 1-year-old pregnant Yorkshire gilt was found dead with no previous clinical signs. Gross findings included metritis, splenomegaly, and valvular endocarditis. Bacterial endocarditis (in the mitral and tricuspid valves) and metritis with dissemination to multiple organs was diagnosed by using histologic examination. Gram-negative coccobacillary organisms present in the valvular lesions were characterized as Actinobacillus equuli by using polymerase chain reaction examination on formalin-fixed, paraffin-embedded tissues (FFPE). A. equuli is rarely reported as a cause of septicemia in pigs in Europe. A. equuli in pigs in the United States has been reported only twice and not, to our knowledge, in the last 30 years. This is the first time that molecular techniques have been used to characterize this organism in FFPE porcine tissues.
Actinobacillus spp., other than Actinobacillus pleuropneumoniae, are considered opportunistic pathogens in pigs. Actinobacillus spp., including A. suis, A. equuli, and A. ureae, can produce abortion, septicemia, and polyarthritis in pigs. 20 Reports of A. equuli infection in pigs are uncommon and come mostly from European countries. 2, 4, 7, 10, 12, 14– 16, 24, 26– 28 To our knowledge, there have been no documented porcine A. equuli infections in the United States in the last 30 years and only 2 reports before that. 7, 27 We describe gross and histologic findings in a pig with septicemic actinobacillosis, confirmed by using molecular methods. A 1-year-old Yorkshire gilt, found dead without previous clinical signs, was submitted for a full necropsy. Grossly, there was splenomegaly and generalized congestion. The kidneys had multiple, dark-red foci in the cortex that were 1 mm in diameter. There was scant serosanguineous fluid in the pericardial sac. Both auricles had subepicardial hemorrhages. The mitral (Fig. 1) and tricuspid (Fig. 2) valve leaflets were irregularly thickened and coated with friable, red-to-tan exudate. A tan-to-red thrombus was attached to the right atrial wall, adjacent to the tricuspid valve (Fig. 2). The lungs were congested and edematous. The right uterine horn contained 3 macerated fetuses. Brown serous fluid was present in the amniotic sac. Tissues examined using histologic analysis included brain, lung, heart, trachea, esophagus, tongue, liver, kidney, multiple lymph nodes, small and large intestines, pancreas, urinary bladder, uterus, adrenal gland, bone marrow, and spleen. In addition to HE, representative sections were stained with Gram, Grocott methenamine silver, and Warthin-Starry stains. Using histologic analysis, affected cardiac valves were shown to have adherent thrombi containing fibrin, neutrophils, red blood cells, and myriads of Gram-negative bacterial coccobacilli (Figs. 3, 4). Mixed inflammatory infiltrates (neutrophils, lymphocytes, and macrophages) and coccobacilli were multifocally present in the myocardial interstitium. Cardiomyocytes in inflamed areas had variable degeneration or liquefactive necrosis. The endometrium of the right uterine horn was covered by a thick layer of necrotic debris, fibrin, and coccobacilli (Fig. 5). Renal glomerular capillaries were frequently occluded by fibrin or contained numerous leukocytes, including neutrophils and mononuclear cells interpreted as lymphocytes and monocytes (Fig. 6). Fewer glomeruli had extensive lysis of the glomerular tuft, which was replaced by neutrophils, red blood cells, and fibrin. Bacterial colonies, like those in the heart, were present in some glomerular capillaries, hepatic sinusoids, and cerebral vessels. Some cerebral vessels were cuffed by lymphocytes with spongiosis of adjacent neuropil (Fig. 7). Tissues submitted to the bacteriology laboratory were uterus, heart, liver, spleen, and kidney. Each tissue was homogenized, at 1 : 10 ratio, in BHI broth and streaked for isolation with 5% sheep blood on MacConkey agar plates. The plates were incubated at 35°C in the presence of 5% CO2 for 24 to 48 hours. Attempts to isolate relevant microorganisms from these tissues were unsuccessful.
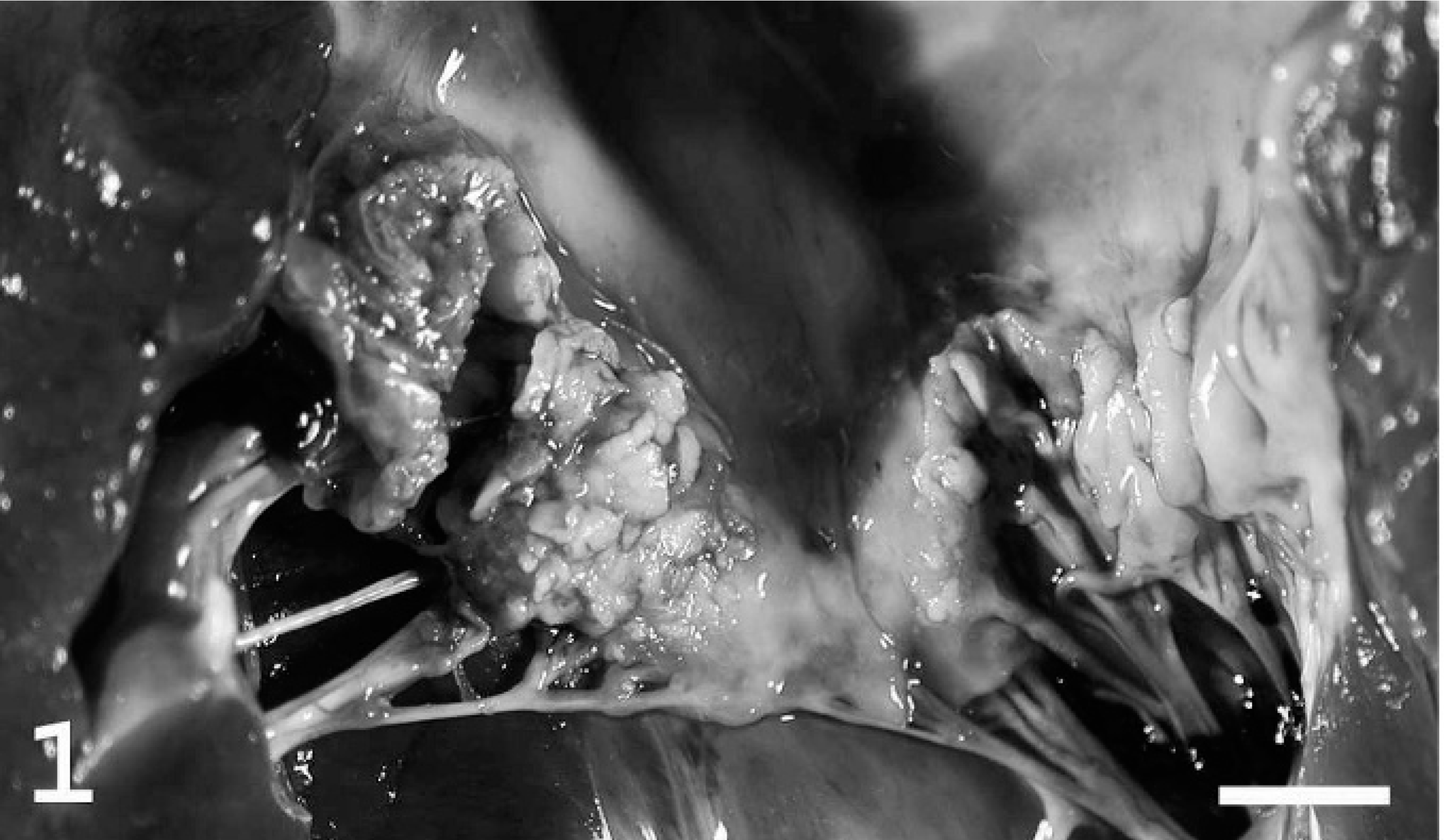
Mitral valve; heart; pig. Vegetative endocarditis. Irregular thickening of valve leaflets with fibrin, bacteria, and leukocytes. This lesion extends to the adjacent atrial myocardium. Bar = 1.5 cm.
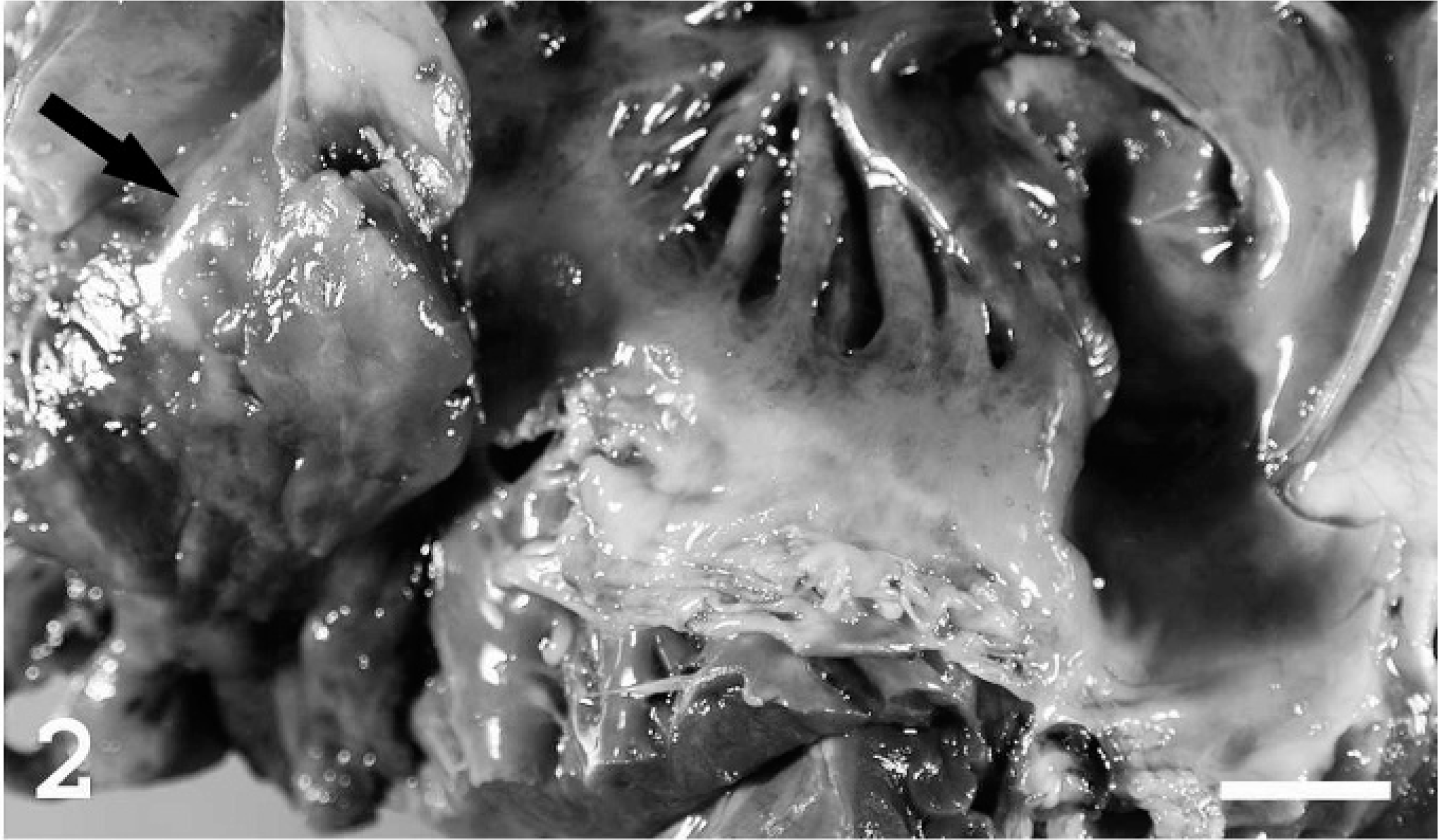
Tricuspid valve; heart; pig. In addition to valvular endocarditis there is a mural thrombus attached to the right atrial wall (arrow). Bar = 1.5 cm.
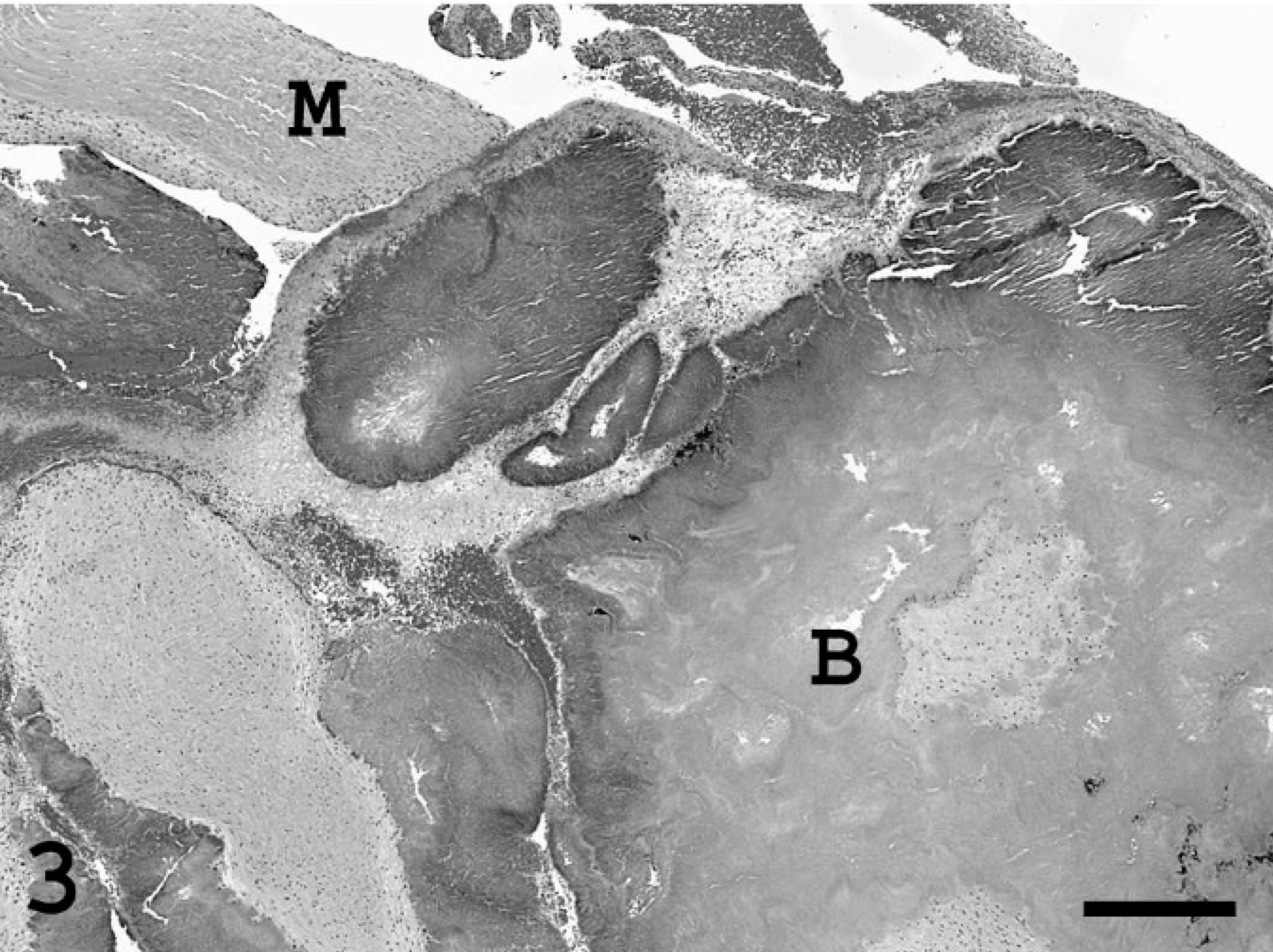
Heart; pig. Portion of the mural thrombus containing numerous bacterial colonies (B). Myocardium (M). HE stain. Bar = 1,400 µm.
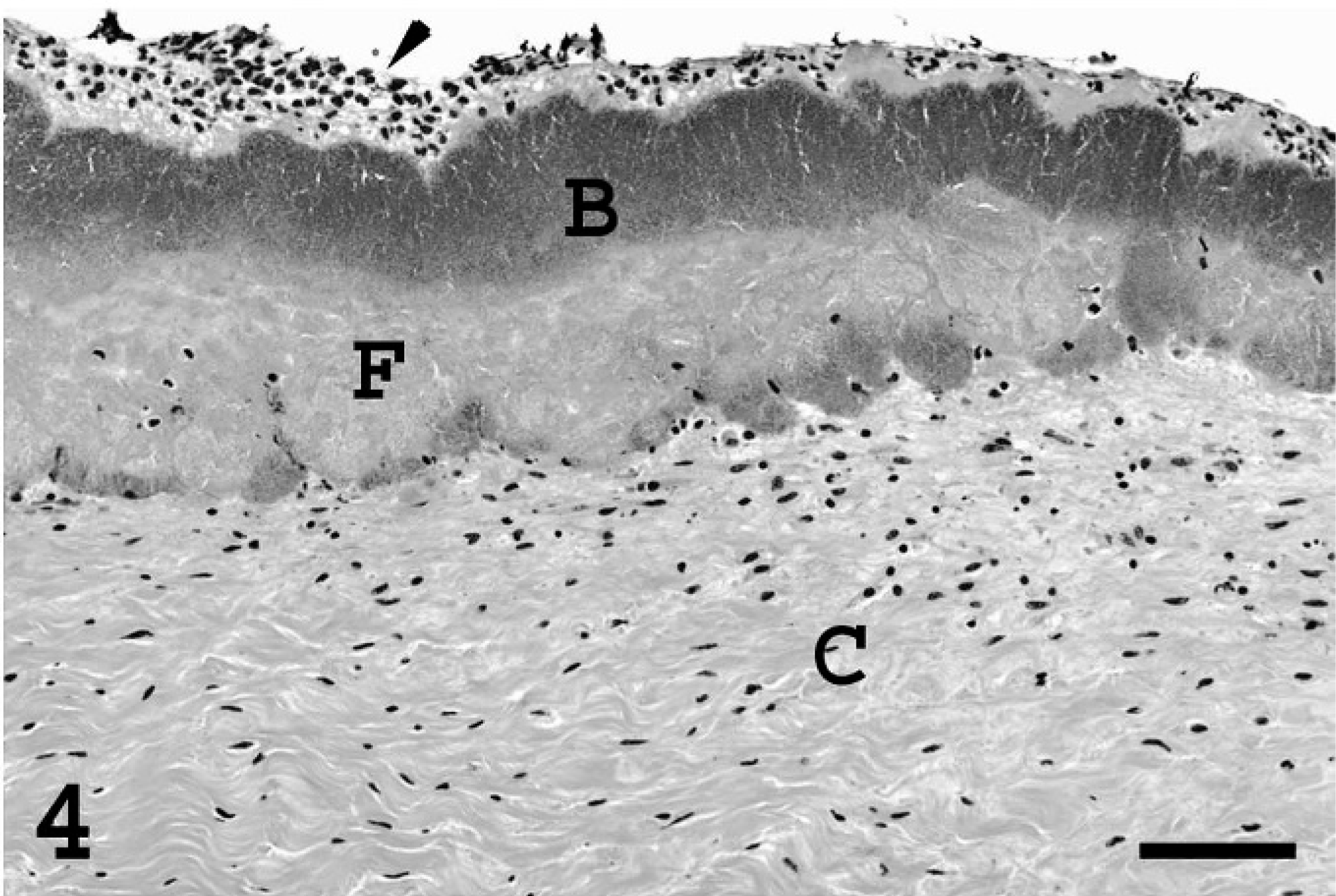
Heart; pig. Section of a valvular thrombus at the level of the corda tendinae (C). Fibrin (F) is deposited on the surface of the corda tendinae and is covered by a layer of bacteria (B) and mixed leukocytes (arrowhead). Bar = 300 µm.
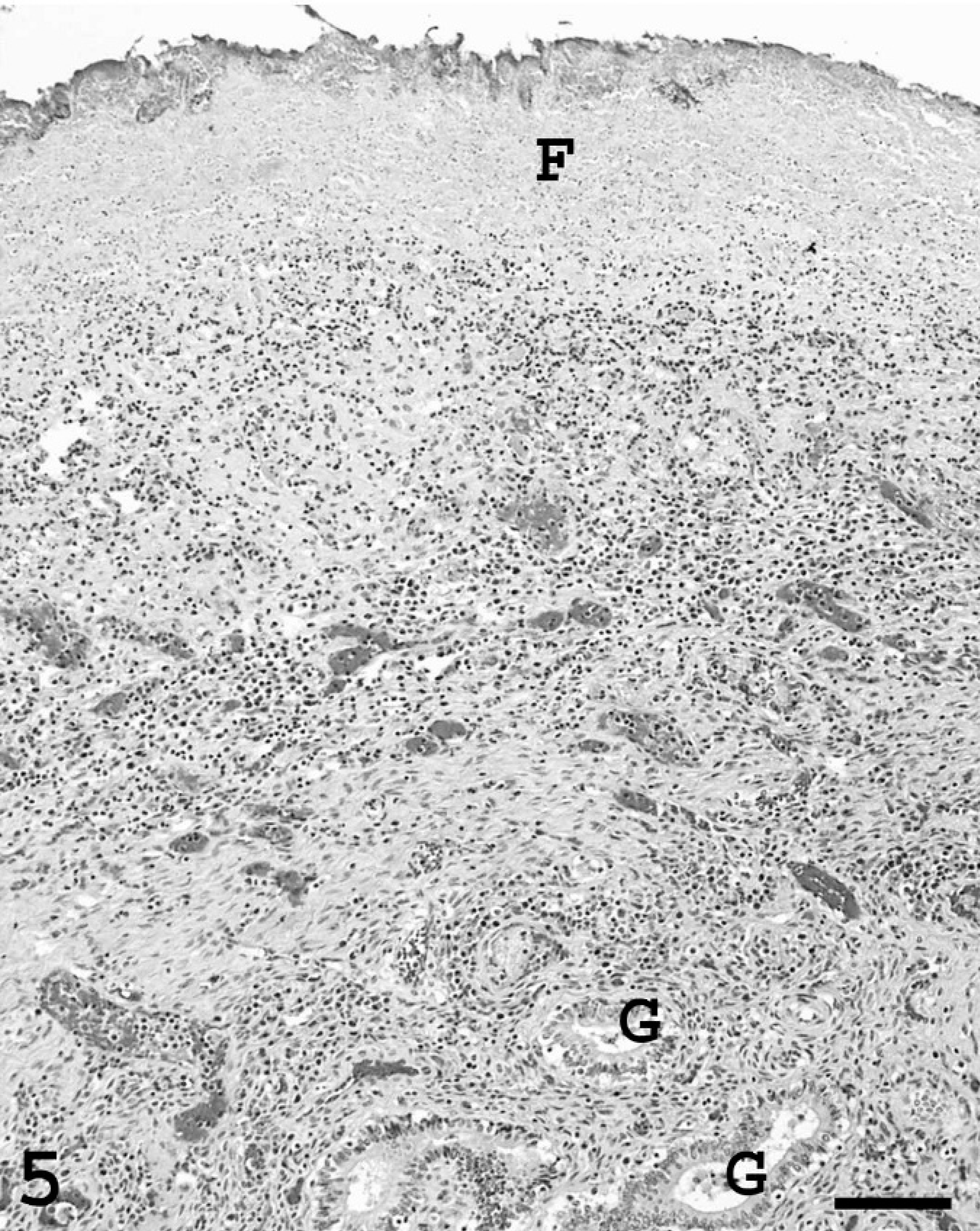
Endometrium; pig. Fibrinosuppurative exudate (F) containing bacterial colonies covers the endometrial surface. There is extensive inflammation and neovascularization in the underlying tissue. Uterine glands (G). Bar = 600 µm.
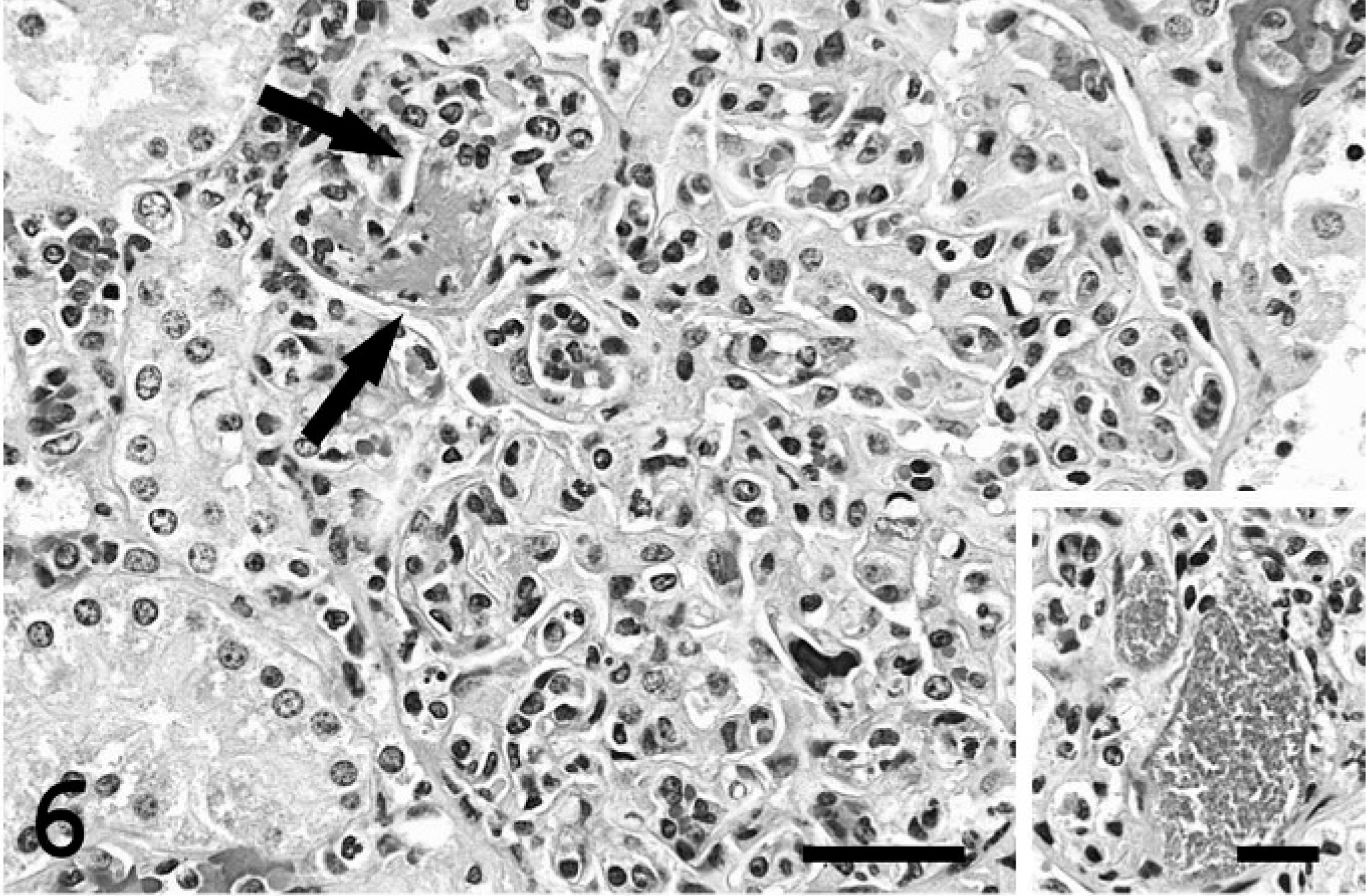
Kidney; pig. Glomerulonephritis. This glomerulus is hypercellular due to marked leukocytosis. Fibrin thrombus (arrows). Bar = 150 µm. Inset. Dilated glomerular capillaries filled with bacterial colonies. Bar = 75 µm.
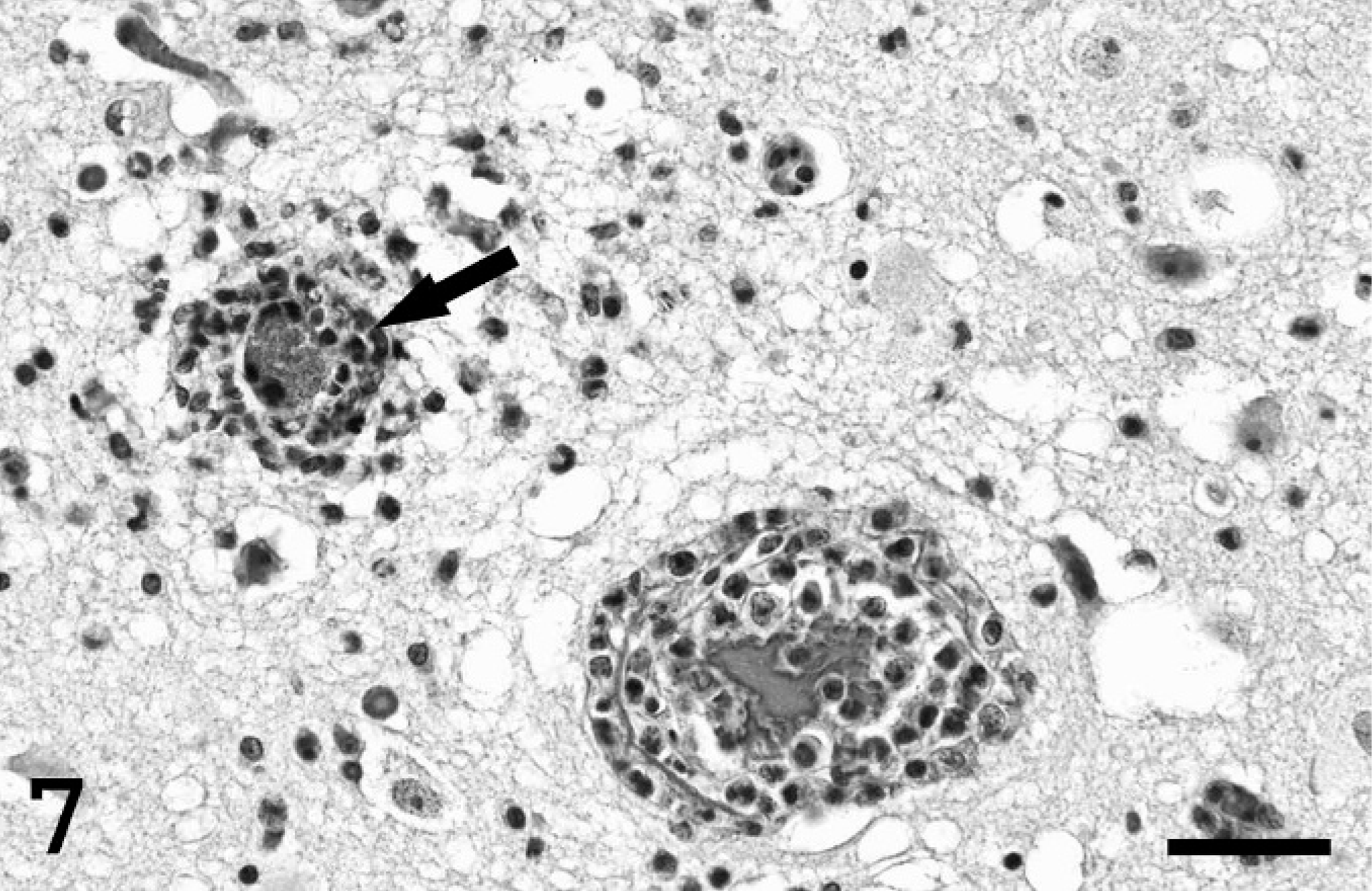
Cerebrum; pig. Two capillaries are cuffed with lymphocytes and rare neutrophils. The lumen of 1 vessel (arrow) is occluded by bacterial colonies. Spongiosis is present in the adjacent neuropil. HE stain. Bar = 150 µm.
A diagnosis of bacterial metritis and valvular endocarditis with secondary septicemia was made.
To characterize the bacterial colonies present in the heart valves by molecular methods, formalin-fixed, paraffin-embedded tissues were used. Briefly, 5 paraffin sections, each 10 µm thick, were cut from a paraffin block containing the heart valve with endocarditis. The paraffin was removed from these sections using xylene. DNA in the deparaffinized tissue sections was extracted using a DNAeasy kit for animal cells or bacteria (Qiagen, Valencia, CA) following the procedures recommended by the manufacturer. Five microliters of the extracted DNA was used as the template in a 50 µl polymerase chain reaction (PCR) mixture (PCR buffer, 1.5 mM MgCl2, 0.2 mM each deoxynucleoside triphosphate, 0.4 µM of each primer, and 1.5 U of Taq polymerase) to amplify a 320 base-pair 16S ribosomal RNA (rRNA) gene sequence. 9 The sequences of the primers used were 16SFa: 5′-GCTCAGATTGAACGCTGG-3′; 16SFb: 5′-GCTCAGGAYGAACGCTGG-3′; and 16SR: 5′-TACTGCTGCCTCCCGTA-3′. 8, 9 16SFa and 16SFb are sense primers and 16SR is antisense primer. The PCR conditions were 5 minutes of denaturation at 94°C, followed by 35 amplification cycles consisting of 1 minute for denaturation at 94°C, 1 minute of annealing at 58°C, and 1 minute of extension at 72°C, with a final extension cycle of 7 minutes at 72°C. The PCR amplicon (320 base pairs) was visualized on a 1.5% agarose gel containing ethidium bromide (data not shown), excised using a sterile scalpel blade under UV illumination, and submitted for direct sequencing of both strands of DNA (Purdue Genomics Center, West Lafayette, IN). Forward and reverse sequences were subjected to a GenBank search for similarities 9, 19 using the on-line Basic Local Alignment Search Tool (BLAST) program provided by the National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/BLAST). Sequencing of amplified 320 base-pair16S rRNA gene (encompassing Actinobacillus species-specific sequences) from nucleic acid extracted from paraffin sections had 100% homology with the published sequence for 2 Actinobacillus equuli isolates: GenBank AY634640 and AY465359 (Table 1). In addition, PCR typing of the repeat in toxin (RTX) genes was negative for apxICA, apxIICA, and aqxA genes. These results are consistent with A. equuli and different from A. suis, in which apxICA and apxIICA genes are present. 13, 18
16S ribosomal RNA partial sequence for Actinobacillus equuli ∗

16S rRNA partial nucleotide sequence in the variable region of bacterial 16S rRNA (320 base pairs) amplified from DNA extracted from paraffin-embedded valvular lesions in the present case had 100% homology with two A. equuli isolates: GenBank AY634640 and AY465359.
Septicemic actinobacillosis in pigs is usually attributed to A. suis. 17, 23 Porcine A. equuli infection has been reported in Europe. 2, 4, 7, 10, 12, 14– 16, 24, 26– 28 To our knowledge, porcine A. equuli infections in North America have not been reported in the last 30 years; Cutlip et al. 5 isolated Actinobacillus spp. in septicemic pigs that resembled A. equuli but produced hemolysis in blood agar, which is rarely reported in A. equuli 2, 3, 25, 27, 28 but typically present in A. suis. 25, 26 A. equuli infections are common in young foals and usually produce septicemia with multiple organs involved, including the kidney and liver as well as joints. 21 Similarly, A. equuli can produce septicemia in piglets and sows, with lesions in joints and kidneys and, sometimes, endocardial localization. 1, 4, 7, 14– 16 A. equuli has also been isolated in metritis in sows, 7 porcine abortion, 27 and as a secondary infection in swine fever. 10 The gilt in our case had metritis, endocarditis, and evidence of bacterial embolism. The sequence of events in this case was initial infection of the gravid uterus with secondary bacteremia, bacterial colonization of cardiac valves, and thromboembolism. 22 Bacterial endocarditis in pigs is usually the result of Streptococcus and Erysipelothrix rhusiopathiae infections, 22 but valvular endocarditis has been reported in natural and experimental A. equuli infections of piglets 1, 12 and in natural infections in foals. 11, 22 The rarity of isolating A. equuli in North American pigs might reflect the separation of pigs and horses in modern farming systems; 23 however, the farm in which this pig was raised did not provide contact with horses. The failure to isolate microorganisms from the tissues could be due to one or more of the following: autolysis, antibiotic use, presence of cellular components that are inhibitory to bacteria growth, or focal rather than bacterial colonization. The postmortem interval was estimated to be from 10 to 15 hours. Inflammatory changes present in multiple organs were not difficult to observe despite moderate-to-advanced autolysis. Organs in which autolysis significantly affected microscopic assessment were the pancreas and intestines.
A. equuli and A. suis produce similar lesions in pigs. 25 However, A. equuli differs from A. suis by being nonhemolytic, producing acid from mannitol, and not hydrolyzing esculin. 23 Attempts to isolate pathogenic bacteria from various tissues, including the heart valve, were unsuccessful in the present case. A molecular approach was sought as an alternative to detect and identify the bacteria in the affected heart valve. Bacterial 16S rRNA has highly conserved nucleotide sequences that are shared by all bacterial species and are interspersed with variable sequences that are genus or species specific. Primers 16SFa/16SFb and 16SR have been proved able to amplify a 320 base-pair hypervariable region at the 5′ end of 16S rRNA and are also complimentary to a wide range of different bacteria genera and species. 9 Therefore, primers 16SFa/16SFb and 16SR can be used in a broad range of PCRs as well as specific 16S rRNA PCR for detection and differentiation of bacterial 16S rRNA in clinical specimens. 6, 9 As shown in the present case, bacterial DNA extracted from the paraffin-embedded valvular lesions was successfully amplified by using PCR with the primers 16SFa/16SFb and 16SR to generate a 320 base-pair amplicon from bacterial 16S rRNA. Analysis of the amplified nucleotide sequences revealed 100% homology with 2 A. equuli isolates (GenBank AY634640 and AY465359). In addition, PCR typing of the RTX gene ruled out A. suis and confirmed A. equuli subsp. equuli. The lack of detection of aqxA gene also ruled out A. equuli subsp. haemolyticus. 18 Although porcine actinobacillosis as the result of A. equuli has not been confirmed in pigs in the United States in the last 30 years, diagnosticians should consider this pathogen in the differential diagnosis of septicemic conditions of pigs.