
Abstract
A 2 and a half-year-old male California sea lion (Zalophus californianus) presented with a history of intermittent generalized seizures. Magnetic resonance imaging revealed a large focal mass occupying the right cerebral hemisphere with moderate dilatation of the contralateral lateral ventricle. At necropsy, the right cerebral hemispheric white matter was expanded by numerous irregularly shaped, pale pink nodules up to 10 mm in diameter. The overlying cortex was characterized by increased numbers of small, poorly developed gyri with shallow, often indistinct, sulci (polymicrogyria). Microscopically, nodules were composed of neurons, oligodendroglia, microglia, and supporting neuropil and were well delineated from the surrounding white matter. The gross, histological, and immunohistochemical features of this lesion are consistent with a neuronal migration defect resulting in unilateral subcortical heterotopia.
Keywords
In humans, cortical dysplasia is relatively common in patients with epilepsy and mental retardation. Disruption of neuronal migration due to a fundamental fault in the migratory process or an early insult to neuronal progenitor cells during development of the cerebral cortex has been suggested for many of these conditions. 11, 15 The formation of neuronal heterotopias within cerebral white matter is one such manifestation. Traditionally, heterotopias due to disruption of migration were classified as nodular or laminar. Clinically, it was found more useful to divide heterotopias into 3 categories: 1) subependymal, 2) subcortical, and 3) band heterotopia. 3 Subcortical heterotopias are defined as “irregularly lobulated masses of gray matter that are located primarily in the subcortical white matter of the cerebral hemispheres.” 2
Reports of developmental lesions or malformations resulting in cortical dysplasia are not uncommon in the veterinary literature. Although often idiopathic, frequently reported etiologies include in utero infections, maternal toxin exposure, and those with a genetic basis. 8, 17 Well-characterized examples with distinctive lesions include ovine orbiviral hydranencephaly, holoencephaly in lambs caused by maternal ingestion of Veratrum californicum, and lissencephaly in Lhasa Apso dogs. 8, 17 Sporadically, hamartomatous lesions are also described. 7, 13 To our knowledge, there are no reports in the veterinary literature of subcortical heterotopia as a result of a neuronal migration defect.
At 1 year of age, a captive-born male California sea lion had a single, self-limiting, generalized seizure. The subsequent physical exam, neurological exam, complete blood cell count, and clinical chemistry were unremarkable. For approximately 1 year, no additional seizures were observed and the animal progressed in training and development along with his peers. At 2 years of age, another self-limiting, generalized seizure was observed. Again the physical exam, neurological exam, complete blood cell count, and clinical chemistry were unremarkable. Protozoal and morbilliviral serology and domoic acid assays were negative. To explore a possible anatomical basis for the seizures, a magnetic resonance imaging (MRI) study was performed. The T2-weighted MRI study done without contrast revealed a large focal mass occupying or replacing much of the right cerebral hemisphere. The overlying cortex was thin and irregular, and there was moderate dilatation of the left lateral ventricle (Fig. 1). Within weeks, multiple generalized seizures were observed, and the animal was placed on appropriate anticonvulsant therapy. Over the following months, the animal was observed as becoming progressively more ataxic during land-based training sessions and had brief periodic episodes of altered behavior and apparent awareness interpreted as petit mal seizures. A second MRI was ordered to better establish the limits and anatomical relationship of the lesion. However, the animal expired during this procedure.
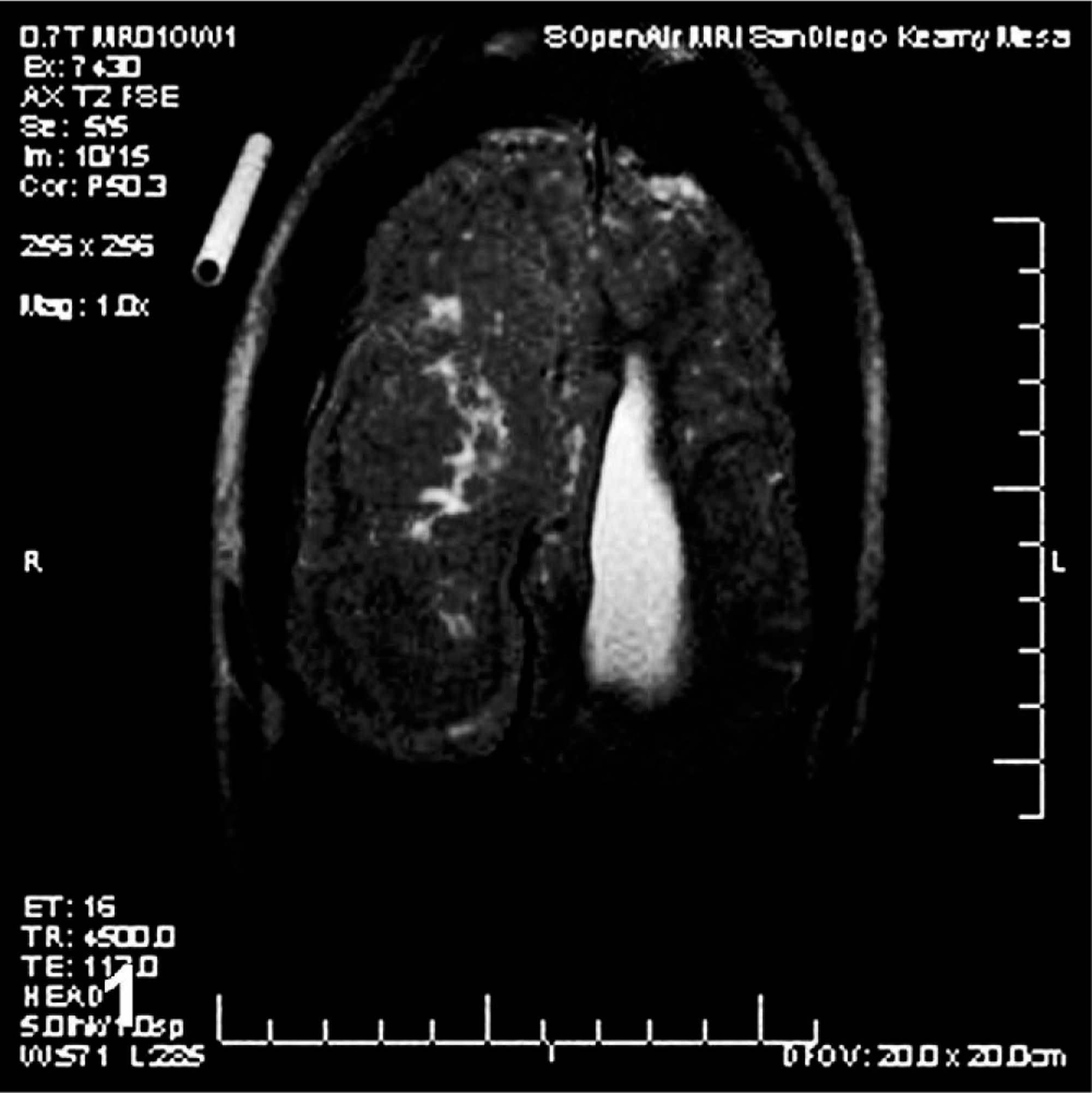
Brain; California sea lion. T2-weighted horizontal magnetic resonance image without contrast. Note the mass occupying the right hemisphere, the thin overlying cortex, and dilatation of the left lateral ventricle.
At necropsy, the right cerebral cortex was characterized by increased numbers of small, poorly developed gyri with shallow, often indistinct, sulci (polymicrogyria [PMG]). Focally, there was an area of cortical thinning and loss near the midline. The gyri and sulci of the left cerebral cortex were normal (Fig. 2). There was visible deformation of the calvarium with deviation of the crista galli and internal sagittal crest to the left. On transverse sectioning, examination of the brain revealed asymmetrical cerebral hemispheres with enlargement of the right cerebral hemisphere relative to the left cerebral hemisphere. The right cerebral hemisphere was characterized by numerous irregularly shaped, pale pink nodules that extended from the ventricular surface through the hemispheric white matter to the cerebral cortex running from the cranial aspect of the frontal lobe to the caudal aspect of the occipital lobe. Nodules were variably sized, up to 10 mm in diameter, and well demarcated from the surrounding white matter. There was compression and ventromedial displacement of basal nuclei and diencephalic structures. The right lateral ventricle was compressed and displaced ventrally and laterally. There was moderate dilatation of the left lateral ventricle (Fig. 3).
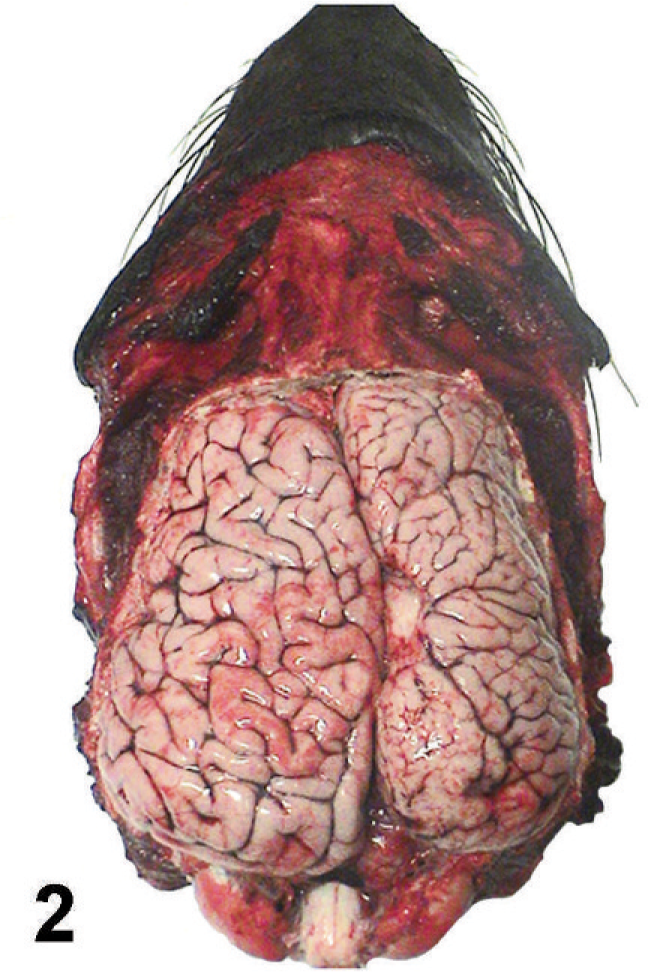
Cerebral hemispheres; California sea lion. Gyri of the right hemisphere are small and increased in number. Sulci are shallow and often indistinct. Note the focal loss of cortical tissue.
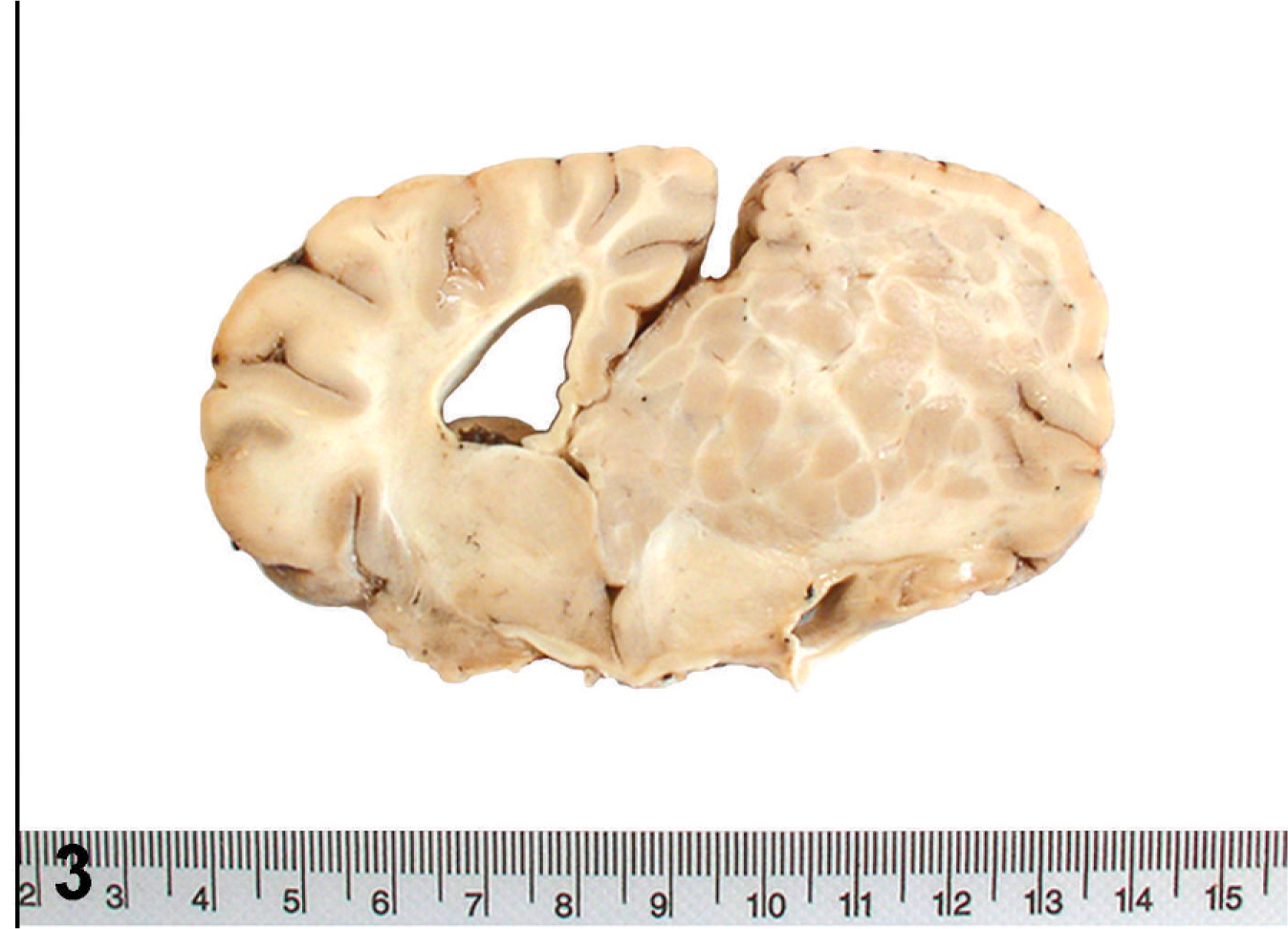
Subcortical heterotopia; California sea lion. A single nodular mass occupies the right cerebral hemisphere, expanding the centrum semiovale, and extending across midline compressing the contralateral hemisphere.
The entire brain was fixed in 10% neutral buffered formalin. Multiple representative sections, including sections through the frontal lobe, occipital lobe, parahippocampal gyrus, caudate nucleus, thalamus, mesencephalon, and cerebellum, were processed routinely; embedded in paraffin; cut at 4-μm-thick sections; and stained with hematoxylin and eosin. Serial sections were also examined by immunohistochemistry using the avidin-biotin peroxidase method for glial fibrillary acidic protein (GFAP) (Dako, dilution 1 : 4000), neuron-specific enolase (NSE) (Dako, dilution 1: 800), and myelin basic protein (MBP) (Dako, dilution 1 : 400).
Microscopically, nodules were well delineated from the surrounding white matter and composed of numerous irregularly arranged neurons, oligodendroglia, and microglia along with supporting neuropil (Fig. 4). There was strong NSE immunoreactivity for the numerous neurons within the nodules (Fig. 5). No cells within nodules exhibited GFAP immunoreactivity (Fig. 6), and there was only scattered immunoreactivity for MBP (Fig. 7). The lack of GFAP staining within nodules not only demonstrates an absence of mature astrocytes but also suggests an absence of radial glial cells. Within the surrounding white matter, there was strong multifocal immunoreactivity for GFAP-positive astrocytes (Fig. 6) and strong diffuse immunoreactivity for MBP (Fig. 7). Rare NSE–positive neurons were also observed within the white matter (Fig. 5). The overlying cerebral cortex was hypocellular and of variable thickness. Cortical neurons were irregularly arranged and lacked any apparent lamination or organization.
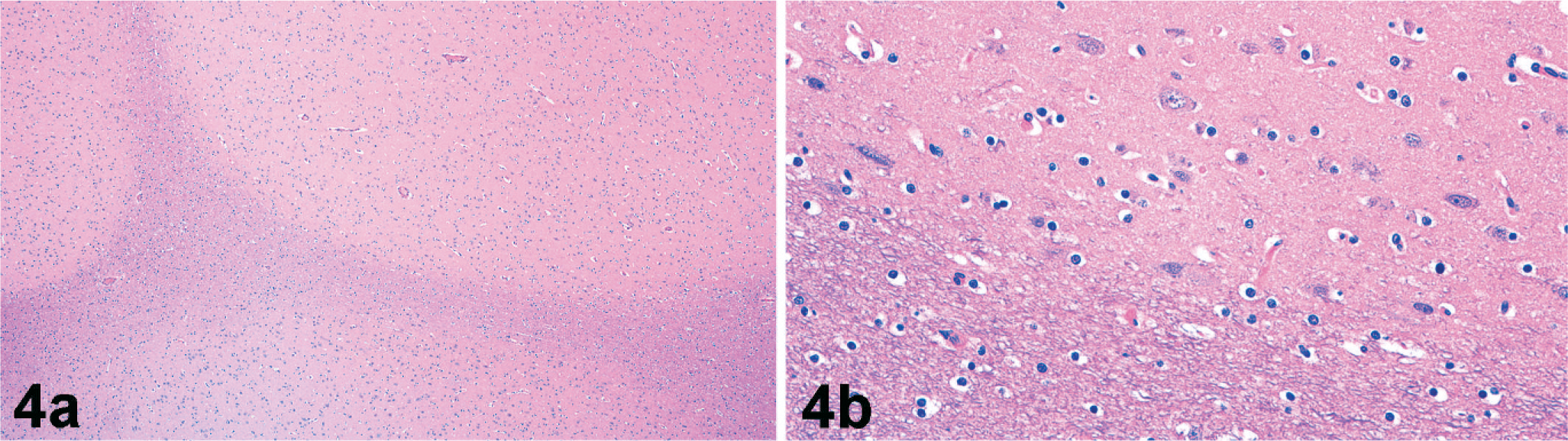
Subcortical heterotopia; California sea lion. Nodules are well delineated from the surrounding white matter tracts (
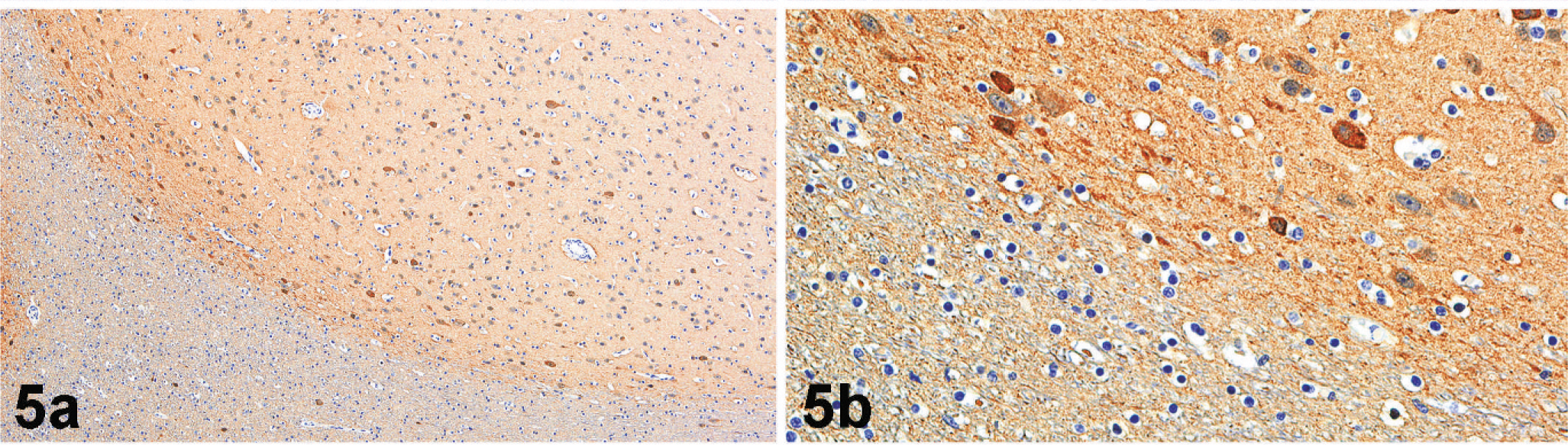
Subcortical heterotopia; California sea lion. Neurons within nodules (upper half of the photomicrograph) express strong immunoreactivity for NSE (
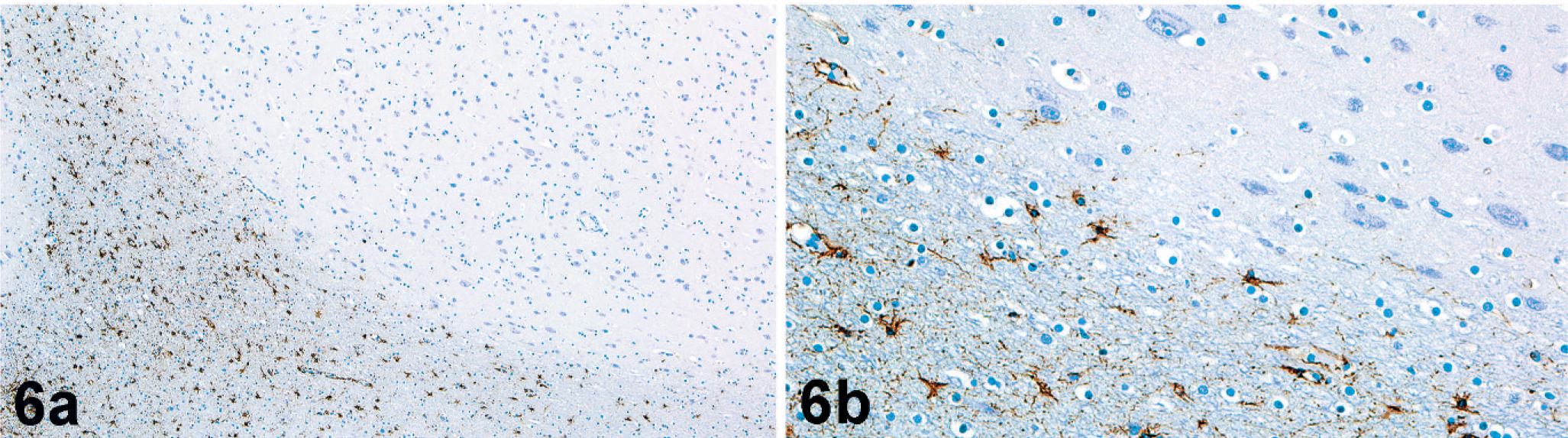
Subcortical heterotopia; California sea lion. Astrocytes within white matter tracts (lower half of the photomicrograph) exhibit strong GFAP immunoreactivity (
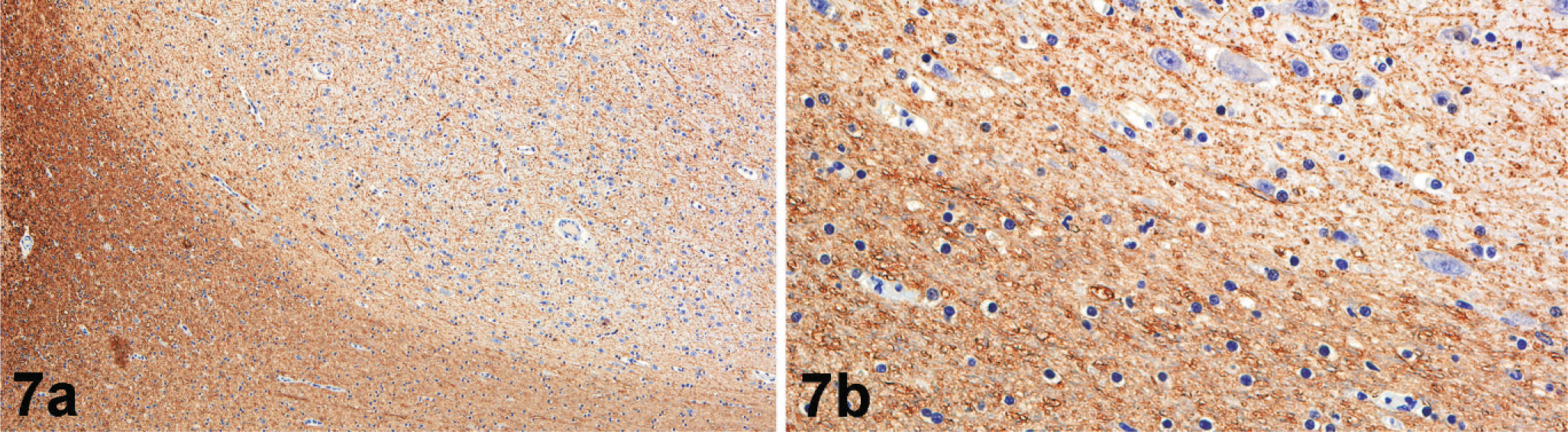
Subcortical heterotopia; California sea lion. There is strong MBP immunoreactivity within white matter tracts (lower half of the photomicrograph) (
Development of the central nervous system is a complex choreographed process requiring cortical neurons to migrate relatively long distances. As the neural plate closes, forming the neural tube, primitive neuroectoderm (or neuroepithelium) is positioned along the ventricle or lumen of the neural tube in an area referred to as the ventricular zone. 5, 9, 15 It is here that neuroectodermal cells replicate becoming neuroblasts (immature differentiated neurons capable of growth but not division) and spongioblasts, which are the progenitors of astrocytes and oligodendrocytes. 8 Telencephalic neuroblasts populate the subcortical-basal nuclei and the cerebral cortex. 8 Population of the cerebral cortex begins with the outward migration of neuroblasts through an intermediate zone along radial glial cell processes that extend from the ventricular surface to the pial surface. 9, 15 The first 2 cell types to migrate out of the ventricular zone to populate the future cortex form what is referred to as the preplate. The outer layer of the preplate is composed primarily of Cajal-Retzius neurons and the inner layer, subplate neurons. 9, 15 Subsequent migrating neurons come to rest between these 2 preplate cell layers, completing the 6-layered cortex in an “inner to outer” fashion. That is, earlier migrating cells reside in deeper cortical layers, whereas later migrating cells pass through the deeper layers to occupy more superficial layers. 10, 15
Proper neuronal migration requires the successful interaction between cell ligands, cell adhesion molecules, and extracellular matrix molecules and the coordinated movements of cytoskeletal proteins. 5, 15 In humans, studies of neuronal migration defects with a familial basis, identification of their associated genes, and observations of their characteristic phenotypes have led to an understanding of the mechanisms underlying neuronal migration. As a result, 4 key steps have been identified: 1) onset of neuronal migration, 2) ongoing neuronal movement, 3) neuronal migration into the preplate, and 4) neuronal migration completion. 9 Conditions best illustrating these 4 key steps include periventricular heterotopia, classical lissencephaly and double cortex, lissencephaly with cerebellar hypoplasia, and cobblestone lissencephaly, respectively. 5, 9, 15 Filamin 1 (FLN1) was identified as the gene underlying periventricular heterotopia. Filamin 1 protein, an actin-binding/cross-linking protein, plays a critical role in the onset of neuronal migration. Mutation results in neuronal nodules lining the lateral ventricle. 5, 9, 10 Classical lissencephaly and double cortex are the result of defects in ongoing neuronal migration associated with mutations in lissencephaly 1 (LIS1) and doublecortin (DCX). LIS1 and DCX are microtubule-binding proteins key in the microtubule stability and reorganization necessary for proper migration. Mutations result in collections of neurons within subcortical white matter. 5 Lissencephaly with cerebellar hypoplasia is the result of a reelin (RELN) gene mutation identified in some families and the Reeler strain of mouse. The reelin protein appears to be responsible for the entry of migrating neurons into the preplate and their proper positioning. 14, 15 In the absence of the reelin protein, migrating neurons populate the cortex in an inverted “outer to inner” fashion. 14, 15 Cobblestone lissencephaly appears to be a defect in the signal responsible for halting neuronal migration. Mutations in the fukutin gene result in the loss of the fukutin protein, a protein expressed by layer 1 neurons, where it is believed to produce a stop signal. Without the appropriate stop signal, migrating neurons pass the cortical plate, penetrate the pia, and present as follicular heterotopias. 9, 15
PMG is defined as excessive cortical infoldings. Grossly, gyri are small and separated by shallow or indistinct sulci. 4, 6 The defect can be focal or widespread. Histologically, PMG has been described as layered, characterized by a cell-sparse zone in layer 5, or unlayered where there is disorganization of cortical layers and nearly random orientation of neurons. 4 The pathogenesis of PMG in humans is less well understood than neuronal migration defects. Most authors suggest ischemia as a major cause, but genetic, viral, and maternal toxicities have also been reported. 4, 18 PMG is also associated with seizure activity and mental retardation and often accompanies other malformations, including nodular heterotopias, enlarged lateral ventricles, and schizencephaly. 1, 6, 18 Schizencephaly, a condition defined as a full-thickness defect of the cortical mantle lined by gray matter, is thought to have etiologies similar to PMG. Although not observed in this case, schizencephaly is commonly observed concurrently with PMG in humans. 12, 19
In addition to germline mutations, other identified causes of heterotopia in humans include injury, ischemia, and infection in the late first trimester or early second trimester. 2 The unilateral distribution of the lesion observed in this young sea lion, as well as the apparent absence of this condition in any siblings, suggest a sporadic insult as the cause rather than a genetic or germline mutation. The nature of the lesion and the accompanying cranial deformity suggest the lesion's presence since before parturition, the likely pathogenesis being an early insult to the ventricular zone. Possibilities in this case include an in utero infection, maternal exposure to a toxin, and an ischemic event. In humans, the age of seizure onset is variable (usually during the first or second decades of life), and the seizure severity is often progressive in nature. 3, 9 Patients with large bilateral heterotopias were more likely to have severe developmental delays or motor dysfunction, whereas those with focal or even extensive unilateral heterotopias were more likely to have less severe (if any) mental retardation with variable motor function. 2, 3 Our case was similar to human reports in that the seizure onset was delayed and the seizure activity was progressive in frequency and severity. In humans with neuronal migration defects or PMG, mental retardation is usually present, although the degree is variable. 3, 9 Interesting in this case was the degree, or lack thereof, of any perceived reduction in the animal's learning or motor abilities, especially for an animal in an intensive and active training program.
Footnotes
Acknowledgements
We thank Dr. Joseph Gleeson for his critical review of this manuscript; Drs. James Barkovich, Lawrence Hansen, and Henry Powell for assistance with microscopic interpretation; Drs. Dale Dunn, Judy St. Leger, and Mark Xitco for their invaluable input; and Steve Van Adestine for photographic assistance.