
Abstract
The equine putative chloride channel protein eCLCA1 is thought to be critically involved in the pathogenesis of recurrent airway obstruction (RAO) via modulation of the hydration of airway mucins. A recent study revealed a strong increase of eCLCA1 messenger ribonucleic acid (mRNA) in the lungs of horses with RAO. In this study, eCLCA1 protein and mRNA expression were quantified in airway goblet cells of 9 horses affected with RAO and 9 control horses by using immunohistochemistry and laser microdissection followed by real-time quantitative reverse transcription polymerase chain reaction, respectively. Horses affected by RAO had strong goblet cell metaplasia in bronchioles and goblet cell hyperplasia in bronchi and the trachea. Expression of the eCLCA1 protein was tightly linked to all airway goblet cells in both groups. No differences were detected in the ratio of eCLCA1 mRNA copy numbers to the mRNA copy numbers of the housekeeping gene EF-1a per goblet cell between horses affected with RAO and unaffected horses, suggesting that the increase in eCLCA1 expression is because of increased numbers of goblet cells and not transcriptional upregulation of the eCLCA1 gene. In addition, biochemical analyses of the eCLCA1 protein after in vitro translation and heterologous expression in cultured cells revealed that eCLCA1 is a secreted glycoprotein and not an integral membrane protein. Taken together, the results suggest that eCLCA1 mediates its effect as a soluble constituent of airway mucins that is overexpressed in RAO airways because of goblet cell hyperplasia and metaplasia, not transcriptional upregulation.
Keywords
The term recurrent airway obstruction (RAO) (chronic bronchiolitis, heaves) is used for a condition in mature horses characterized by airway obstruction that can be reversed by a change in the environment or the use of bronchodilators. 19 This definition separates RAO from inflammatory airway disease, which occurs in young horses, as well as from respiratory diseases induced by primary viral or bacterial infections. The principal etiology of RAO is thought to be inhalation of organic stable dusts that contain pollen, molds, endotoxins, or forage mites. 2 The key mechanism is based on allergen-induced hypersensitivity with Th2 cytokine–mediated chronic-airway pathology. On histologic examination, the disease is characterized by lymphocytic bronchiolitis, with variable eosinophilic infiltrates, smooth-muscle hyperplasia, and goblet cell metaplasia in small bronchioles, with overproduction of mucins. 6, 14 Mucus overproduction and increased numbers of neutrophils are key cytologic diagnostic features in smears of tracheobronchial aspirates. Equine RAO shares many clinical and morphologic features with human asthma and chronic obstructive pulmonary disease (COPD), despite some dissimilarities that make it a less than perfect model for the 2 human entities. 3, 21
We recently identified and cloned the first equine member of the family of calcium-activated chloride channels, eCLCA1, which is strongly upregulated in small airways of horses with RAO. 1 Chloride channels in general are prime regulators of epithelial secretion, and many diseases with unbalanced epithelial secretion involve defective or dysregulated chloride channels. 11 Members of the CLCA gene family were recently shown to be key regulators of the asthma phenotype, particularly mCLCA3 (alias gob-5) in mice, and hCLCA1 in people. The rationale for this hypothesis is based on observations that i) hCLCA1 and its murine orthologue, mCLCA3 (alias gob-5), are strongly upregulated in human patients with asthma and murine asthma models, respectively 10, 23 ; ii) experimental overexpression of hCLCA1 and mCLCA3 results in goblet cell metaplasia, mucin overproduction, and exacerbation of the asthma phenotype 10, 18 ; iii) antisense-mediated repression of mCLCA3 expression reduces the asthma phenotype 18 ; iv) functional blocking of the hCLCA1 and mCLCA3 chloride channel activity inhibits goblet cell metaplasia, mucin production, and reduces airway inflammation25; and v) single nucleotide polymorphisms in the hCLCA1 gene affect the susceptibility to human asthma. 12
The equine eCLCA1 is the direct orthologue of mCLCA3 and hCLCA1, and is expressed in the same cell types, namely, in goblet cells of the respiratory and intestinal tracts. 1, 8, 15 However, structural differences between the mCLCA3 protein in mice and the hCLCA1 protein in humans may suggest functional differences between these direct orthologues. 7, 17 Thus, studies on the role of this cluster of CLCA proteins may provide further insight into the mechanisms, similarities and dissimilarities of RAO, asthma, and COPD. Moreover, CLCA proteins have been suggested as potential targets for therapeutic intervention. 24
In this study, we further characterized the role of eCLCA1 in equine RAO. Specifically, we tested the hypothesis of whether eCLCA1 is transcriptionally upregulated in RAO airway epithelia 1 and whether the eCLCA1 protein actually is an integral membrane molecule that could serve as an ion channel in RAO airways. Surprisingly, we failed to identify transcriptional upregulation, despite strong overexpression of the eCLCA1 messenger ribonucleic acid (mRNA) and protein in whole RAO airways. Instead, the increased eCLCA1 mRNA copy numbers in RAO airways appear to be solely because of increased numbers of goblet cells that express this molecule. Moreover, eCLCA1 is a secreted, soluble glycoprotein and has no integral membrane domain and thus more likely acts as a regulator rather than a channel protein on its own.
Materials and Methods
Horses and tissue processing
Lung and trachea specimens were derived from 9 warm-blooded horses affected with RAO (age 6–28 years, median 16 years; weight 439–750 kg, median 550 kg) and 9 warm-blooded control horses (age 7–17 years, median 15 years; weight 248–646 kg, median 500 kg) that were euthanatized or died because of unrelated reasons, and had no clinical history or macroscopic or histologic lesions suggestive of respiratory disease. The diagnosis of RAO was based on clinical history and histologic lesions as defined by Robinson. 19 Tissue specimens of 1 ml were collected from 4 different areas of the cranial and main lobes, respectively, and from 6 defined segments of the airways: central to the cranial, middle and caudal thirds of the trachea; left main bronchus 2 cm caudal of the bifurcation; bronchus from the center of the left main lobe, with approximately 1.0 cm in diameter; and small bronchus from the caudal fourth of the left main lobe, approximately 0.3 cm in diameter. Samples were immersion fixed for 24 hours in 10% neutral-buffered formalin and were routinely embedded in paraffin. From each sample, three 3-μm serial sections were mounted on adhesive glass slides and stained with hematoxylin and eosin or the periodic acid–Schiff (PAS) reaction or were processed for immunohistochemistry. Adjacent tissue samples of similar size were freshly snap frozen in liquid nitrogen and stored at −80°C until further use for laser microdissection and mRNA expression analyses.
Immunohistochemistry
Identification of the eCLCA1 protein on tissue sections was performed as described. 1 Briefly, after dewaxing the mounted tissue samples in xylene and rehydration in isopropanol and graded ethanol, sections were treated for 20 minutes with 0.05% pronase E (Merck, Darmstadt, Germany) in phosphate-buffered saline solution (PBS) at 37°C. Endogenous peroxidase activity was blocked by 0.5% H2O2 in 85% ethanol for 30 minutes at room temperature, followed by 2 washes in PBS that contained 0.05% Tween-20 (PBS/Tween-20) and blocking in PBS/Tween-20 that contained 20% heat-inactivated normal goat-serum. After repeated washes, sections were incubated overnight with anti-eCLCA1 antibody α-eCa1 1 or the respective pre-immune serum diluted 1 : 500 in PBS/Tween-20 that contained 1% bovine serum albumin (Serva, Heidelberg, Germany) in a humid chamber at 4°C. Sections were washed repeatedly in PBS/Tween-20 and incubated at room temperature for 30 minutes with biotinylated goat anti-rabbit immunoglobulins (5 μg/ml; Vector Laboratories, Burlingame, CA) diluted in PBS/Tween-20, followed by repeated washes in PBS/Tween-20. Color was developed for 30 minutes by using freshly prepared ABC solution (Vectastain Elite ABC Kit; Vector Laboratories) diluted in PBS according to the manufacturer's protocol, followed by repeated washes in PBS. Incubation for 5 minutes in PBS that contained 0.05% freshly prepared 3,3′-diaminobenzidine-tetrahydrochloride-dihydrate (Fluka-Chemika, Buchs, Switzerland) and 0.1% H2O2 was followed by final rinsing in PBS and tap water for 15 minutes each. The slides were counterstained with hematoxylin, dehydrated through ascending graded ethanol, cleared in xylene, and cover slipped. Smears of tracheobronchial aspirates of horses affected with RAO were fixed in acetone for 10 minutes followed by immunohistochemical staining as described above.
Morphometry
The percentages of PAS- and eCLCA1-positive cells were determined per total epithelial cells in total cross sections of airways. The diameters of bronchi and bronchioles with goblet cell metaplasia were determined by using an Olympus BX41 microscope equipped with a Colorview II digital camera and the analysis software program (SIS Soft Imaging System, Münster, Germany).
Laser-capture microdissection and reverse transcription
Of each region of interest, 3 consecutive sections of 6-μm thickness from the frozen tissue samples were mounted on glass slides, with the first and third sections covered with a polyethylene naphthalate membrane of 1.35-μm thickness (PALM Microlaser Technologies, Bernried, Germany). These 2 sections were fixed for 2 minutes in 95% ethanol at −20°C. After air-drying for 5 minutes, the sections were stained with hematoxylin in diethylpyrocarbonate (DEPC) treated water for 2 minutes, rinsed in DEPC-treated water for 2 minutes, and stained with eosin in DEPC-treated water for 30 seconds. Sections were dehydrated in ascending graded ethanol (30, 50, 70, 99%), air-dried at room temperature, and stored at −80°C. The section between the 2 sections prepared for laser-capture microdissection was mounted on Superfrost adhesive glass slides (Menzel-Gläser, Braunschweig, Germany) and stained with the PAS reaction according to routine protocols, followed by dehydration and freezing as described above. Goblet cells were identified and digitally marked on the PAS-stained sections, followed by excision and catapulting of the same areas from the preceding and subsequent serial sections mounted on polyethylene naphthalate membranes (PALM MicroBeam C; PALM Microlaser Technologies, Martinsried, Germany). This procedure was found necessary because only brief hematoxylin and eosin staining but not PAS staining was compatible with subsequent isolation of high-quality RNA. For each area of interest, 2.5 × 106 μm2 were excised from both normal horses and horses with RAO, which contained approximately 1 × 103 goblet cells. Cells were laser-pressure catapulted into the caps of 0.5-ml Eppendorf tubes that contained 20 μl of digestion buffer RLT (RNeasy Mini Kit; Qiagen, Hilden, Germany). Tubes were inverted and filled up to 350 μl with RLT buffer, and total RNA was extracted and purified by using the manufacturer's protocol, including digestion with RQ1 RNase free DNase (Promega). Total RNA was reverse transcribed in 20 μl by using 200 U SuperScript III reverse transcriptase (Invitrogen), 100 nmol MgCl2, 50 ng random hexamers, 0.2 mmol DTT, 40 U RNase OUT (Promega, Madison, WI) and 10 nmol of each dNTP at 25°C for 10 minutes followed by 50 minutes at 50°C. Finally, samples were digested with 2 U Escherichia coli RNase H for 20 minutes at 37°C to remove RNA from the cDNA : RNA duplexes.
Quantitative real-time polymerase chain reaction
Primer sequences for eCLCA1 were 5′-GAGGAGTGAACGCAGCCAGAC-3′ (forward) and 5′-TCGGTGATTTGACAAGGTGGGAAG-3′ (reverse) with a calculated amplicon size of 238 bp. To correct for variations in RNA amounts and (cDNA) synthesis efficacy, the cDNA encoding elongation factor-1a (EF-1a) was amplified in parallel in all experiments as a housekeeping gene. 20 Primers for EF-1a were (forward) 5′-CAATGT-CAAGAACGTGTCC-3′ and (reverse) 5′-ACGACGATCAATCTTCTCCTTCAG-3′, with a predicted amplicon length of 213 bp. Calculated melting temperatures ranged between 60.5 and 60.9°C for eCLCA1 primers and 54.8 and 55.0°C for EF-1a primers.
Real-time quantitative reverse-transcription PCR and data analyses were performed by using the MX 3000P Quantitative PCR System and MX Pro software (Stratagene, La Jolla, CA). The reactions were carried out in 96-well polypropylene plates covered with Optical caps (8× strips; Stratagene). In addition to the cDNA samples from respiratory tissues, 10-fold serial dilutions of cDNA from equine rectal mucosa that ranged from 1 : 50 to 1 : 5,000,000 were used to generate a standard curve for estimation of relative copy numbers on each plate for eCLCA1 and EF-1a. The plates contained triplicates of each cDNA sample and no-template controls with water instead of cDNA templates. Every sample was measured in 2 separate PCR runs. During initial optimization runs, the exact primer concentrations and PCR time and temperature conditions were determined. The optimized 25-μl reaction mix contained 2 μl cDNA, 12.5 μl Brilliant SYBR Green QPCR Master Mix (Stratagene), 30 nM reference dye (ROX), and 300 nM (eCLCA1) or 100 nM (EF-1a) of each primer. Cycling conditions used in the final experiments were 10 minutes at 95°C, followed by 40 cycles of 30 seconds at 95°C, 1 minute at 56°C, and 30 seconds at 72°C. The cDNA of interest and the EF-1a cDNA were amplified on the same plate to ensure equal amplification conditions. Specificity of amplification products was confirmed by melting curve analyses. Results were quantified by using the MX Pro Stratagene analysis software, applying the adaptive baseline, amplification based threshold, and moving average algorithm enhancement. For each sample, the cycle threshold (CT) value was calculated based on the normalized baseline corrected fluorescence (ΔRn).
Quantification of target gene expression
For each sample analyzed, the mean CT value based on the results of all experiments was given with the corresponding standard deviation (SD). Relative cDNA copy numbers (means and SD) were calculated on the basis of the results of the standard curves of the same run. Correlation coefficients were always above 0.95. In case the excised tissue areas contained unavoidable nongoblet cell epithelial cells, the EF-1a cDNA copy number per goblet cell was corrected by multiplying the total number of EF-1a cDNA copies per excised sample with the fraction of goblet cells in that sample. 20
Statistical analysis
For percentages of PAS-positive and eCLCA1-expressing cells, as well as the relative copy numbers of eCLCA1 cDNA per copy number of EF-1a cDNA per goblet cell, arithmetic means (M), SDs, and standard errors of the mean were calculated. Differences were considered statistically significant for P values ≤ 0.05 in an unpaired t-test.
Sequence analyses of the eCLCA1 polypeptide
The eCLCA1 amino acid sequence (Genbank accession no. AY524856) was screened for a signal sequence by using the SignalP 3.0 software. Hydrophobicity analyses were carried out by using the software by Kyte and Doolittle 13 to identify putative transmembrane regions.
In vitro translation and protease protection assay
The eCLCA1 open reading frame cloned into pcDNA3.1 was transcribed and translated with the TNT T7 Coupled Reticulocyte Lysate System (Promega) in the presence of L-[35S]methionine as described. 1 Reactions of 25 μl were carried out at 30°C for 90 minutes, with or without 2 μl canine pancreatic microsomal membranes (Promega). A protease protection assay was performed by addition of proteinase K (25 μg/ml; Sigma) in the presence of microsomal membranes on ice for 20 minutes. The reaction was stopped by boiling and analyzed by autoradiography after 10% sodium-dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE).
Identification of glycosylation patterns and Western blotting
HEK293 cells were transfected with the eCLCA1-containing pcDNA3.1 plasmid by using the Lipofectamine TM 2000 method (Invitrogen, Carlsbad, CA), as described. 17 At 48 hours after transfection, the cells were washed twice with PBS and incubated with medium for 7 hours. The cells and the medium were analyzed separately. The medium was centrifuged for 5 minutes at 1,000 × g and proteins were precipitated from the supernatant with 2 volumes of ethanol. The cells were washed twice with PBS and solubilized in 0.5 ml lysis buffer for 30 minutes on ice as described. 17 The cell lysate and the medium were treated with Endo H and PNGase F (New England Biolabs, Frankfurt, Germany) according to the manufacturer's protocol. The samples were subjected to 10% SDS-PAGE and electroblotted onto nitrocellulose membranes. Membranes were blocked with Tris-buffered saline solution (pH 7.5) that contained 0.1% Tween-20 and 5% non-fat milk and probed with antibody α-eCa1 (1 : 1,000). 1 Membranes were then incubated with peroxidase-conjugated goat anti-rabbit immunoglobulin (1 : 10,000; Jackson ImmunoResearch, West Grove, PA) and were developed by using enhanced chemiluminescence (Amersham, Arlington Heights, IL).
Results
Histopathology
All 9 horses diagnosed with RAO had various degrees of lymphocytic and plasmacytic bronchiolitis with smooth-muscle hyperplasia and marked goblet cell hyperplasia in hematoxylin and eosin (HE) stained tissue sections. Clogging of small airways with mucus and occasional neutrophils and cellular debris was a consistent finding. Infiltrating eosinophils were found in 3 horses. Alveolar emphysema was virtually absent. Larger bronchi and trachea had minimal to no lymphocytic infiltrations. The 9 control horses had none of these findings and were free of any other lesions in the lung and airways. Digital morphometric image analyses identified goblet cells in bronchioles of down to 78 μm in diameter in horses affected with RAO, whereas the smallest airway that contained goblet cells in control horses was 441 μm in diameter.
Quantification of goblet cell hyperplasia and eCLCA1 expressing cells
The numbers of goblet cells identified in PAS reactions were markedly increased in the bronchioles of horses affected with RAO (Fig. 1A) when compared with bronchioles of unaffected horses (Fig. 2A). In fact, none of the control horses had any goblet cells in bronchioles of approximately 400 μm in diameter or less, which had significant numbers of goblet cells in horses affected with RAO. Bronchioles were defined anatomically by the absence of hyaline cartilage that accompany the airway. In contrast to the HE-stained tissue sections, goblet cell hyperplasia was more easily detected by using the PAS reaction in bronchi and all segments of the trachea (Fig. 3A) when compared with unaffected horses (Fig. 4A).
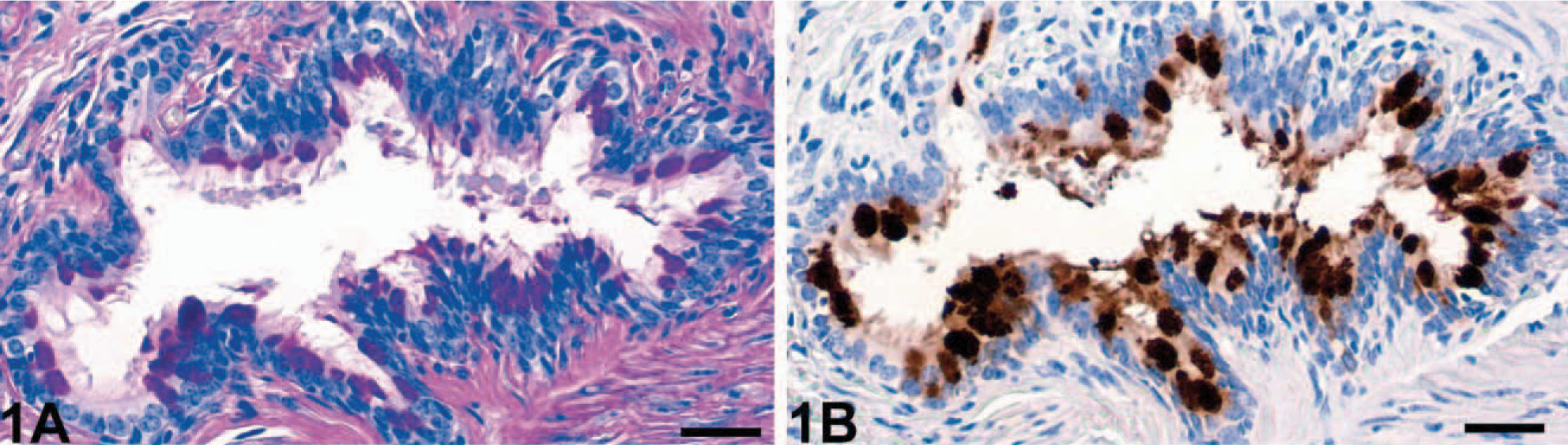
Bronchiolus; RAO-affected horse No. 4. Goblet cell hyperplasia (
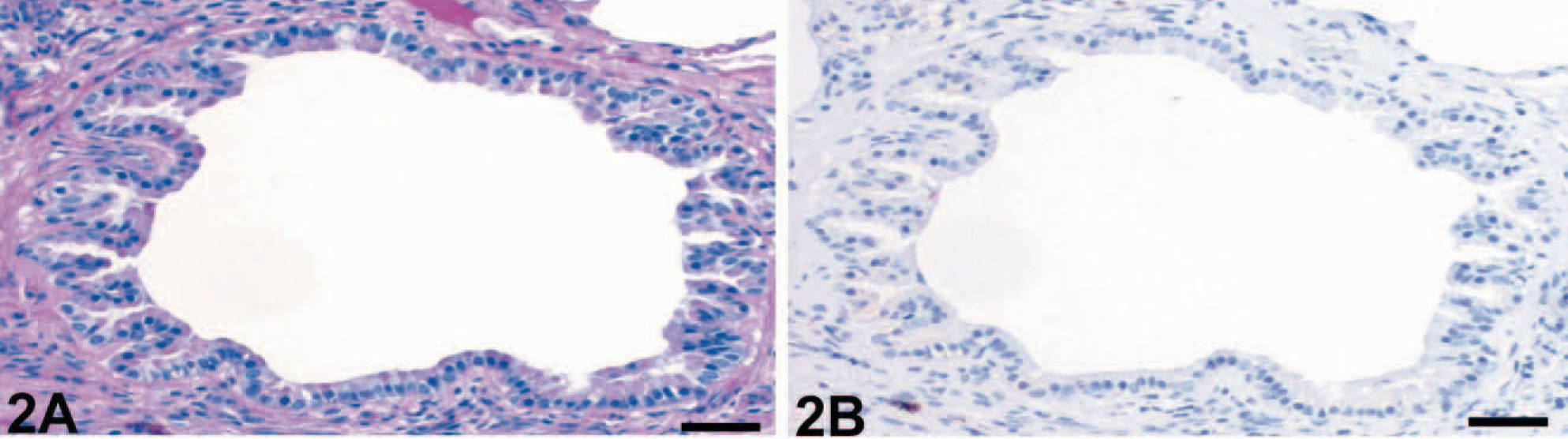
Bronchiolus; control horse No. 15. No lesions were observed. PAS reaction (
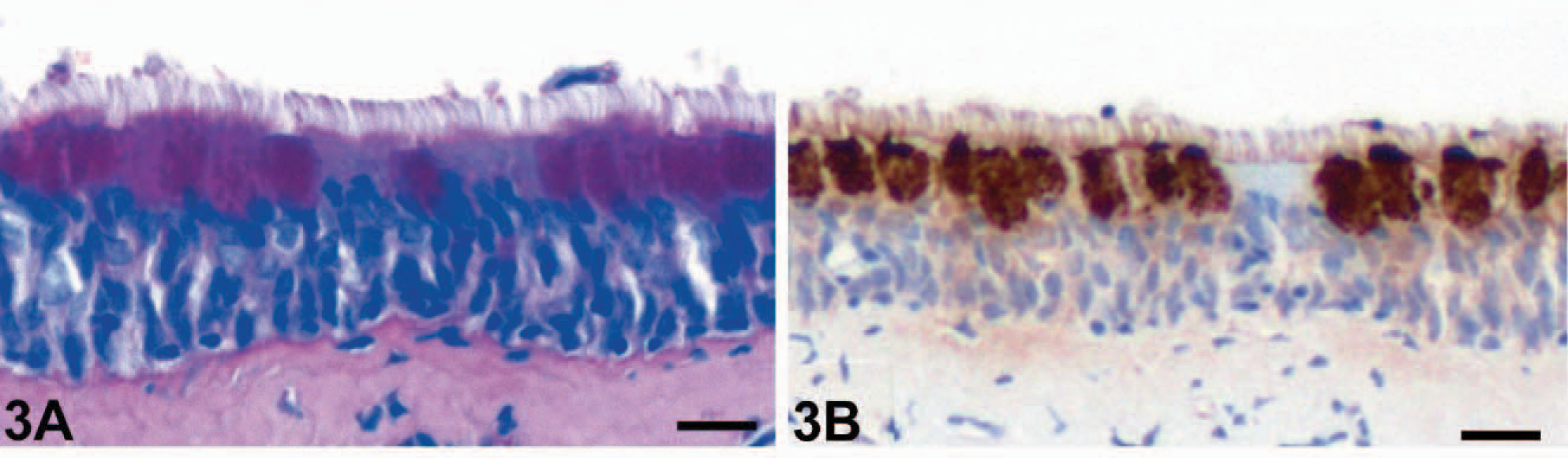
Trachea, middle segment; RAO-affected horse No. 4. Goblet cell hyperplasia (
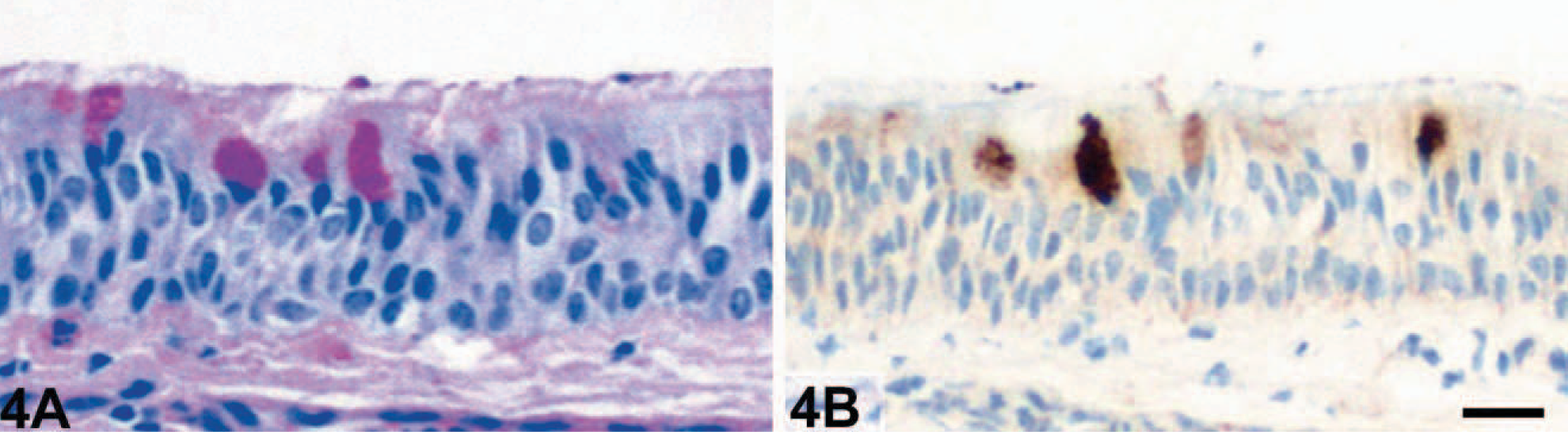
Trachea, middle segment; control horse No. 15. No changes were observed. PAS reaction (
Immunohistochemical detection of eCLCA1 confirmed exclusive localization of the protein in cytoplasmic granules of goblet cells in both horses affected with RAO and control horses, as reported previously. 1 Cells that stained positive for eCLCA1 were virtually identical with PAS-positive goblet cells. Consequently, bronchioles of horses affected with RAO had a strong expression of eCLCA1 (Fig. 1B), whereas no staining was observed in bronchioles of control horses (Fig. 2B). Again, all cross sections of larger bronchi and the trachea had more eCLCA1 positive cells in the epithelium in horses affected with RAO when compared with unaffected horses (Figs. 3B, 4B).
Counting of PAS-positive goblet cells revealed statistically increased percentages of goblet cells in all airway segments of horses affected with RAO compared with unaffected horses (Fig. 5A). The strongest difference was found in bronchioles, with mean goblet cell percentages of 33% in horses affected with RAO and virtually no goblet cells in control horses. The highest percentages of goblet cells were found in the small- and medium-sized bronchi. All 3 segments of the trachea had less goblet cells than the bronchi in horses affected with RAO, as well as in control horses.
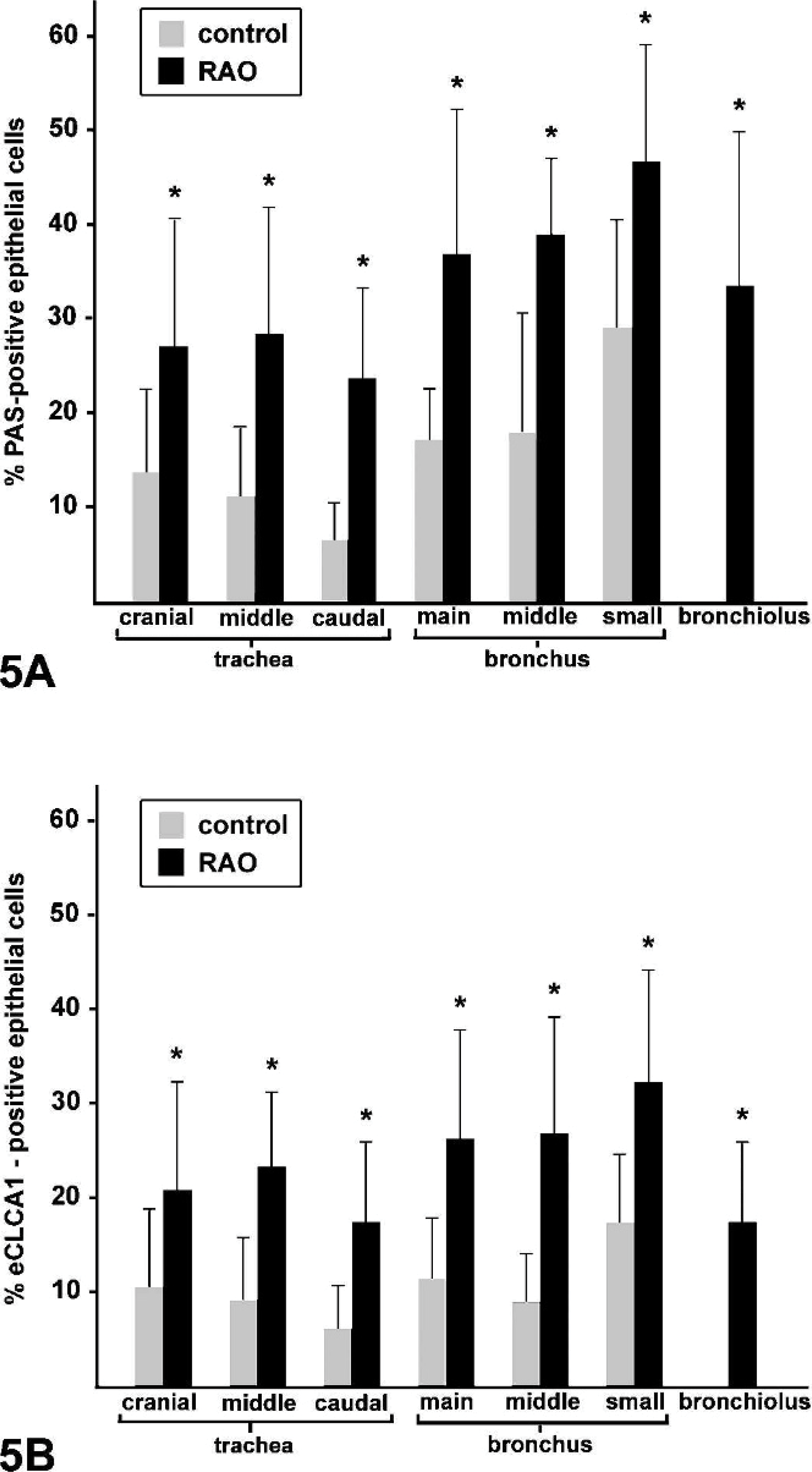
Quantification of the percentage of PAS positive cells (
The percentages of eCLCA1 expressing cells within the epithelial lining of the different airway segments were very similar to the percentages of PAS-positive cells (Fig. 5B). The only difference was that eCLCA1-positive cells were slightly less than PAS-positive cells, resulting in ratios of eCLCA1 expressing cells to PAS-positive cells of between 0.56 and 0.89 (Fig. 6). When the ratios of eCLCA1 to PAS-positive cells were compared between horses affected with RAO and control horses, no significant differences were observed in any of the locations analyzed (Fig. 6). In conclusion, the number of eCLCA1 expressing cells was similarly correlated with the number of PAS-positive goblet cells in both groups, suggesting no change in eCLCA1 expression in RAO airways on the single-cell level.
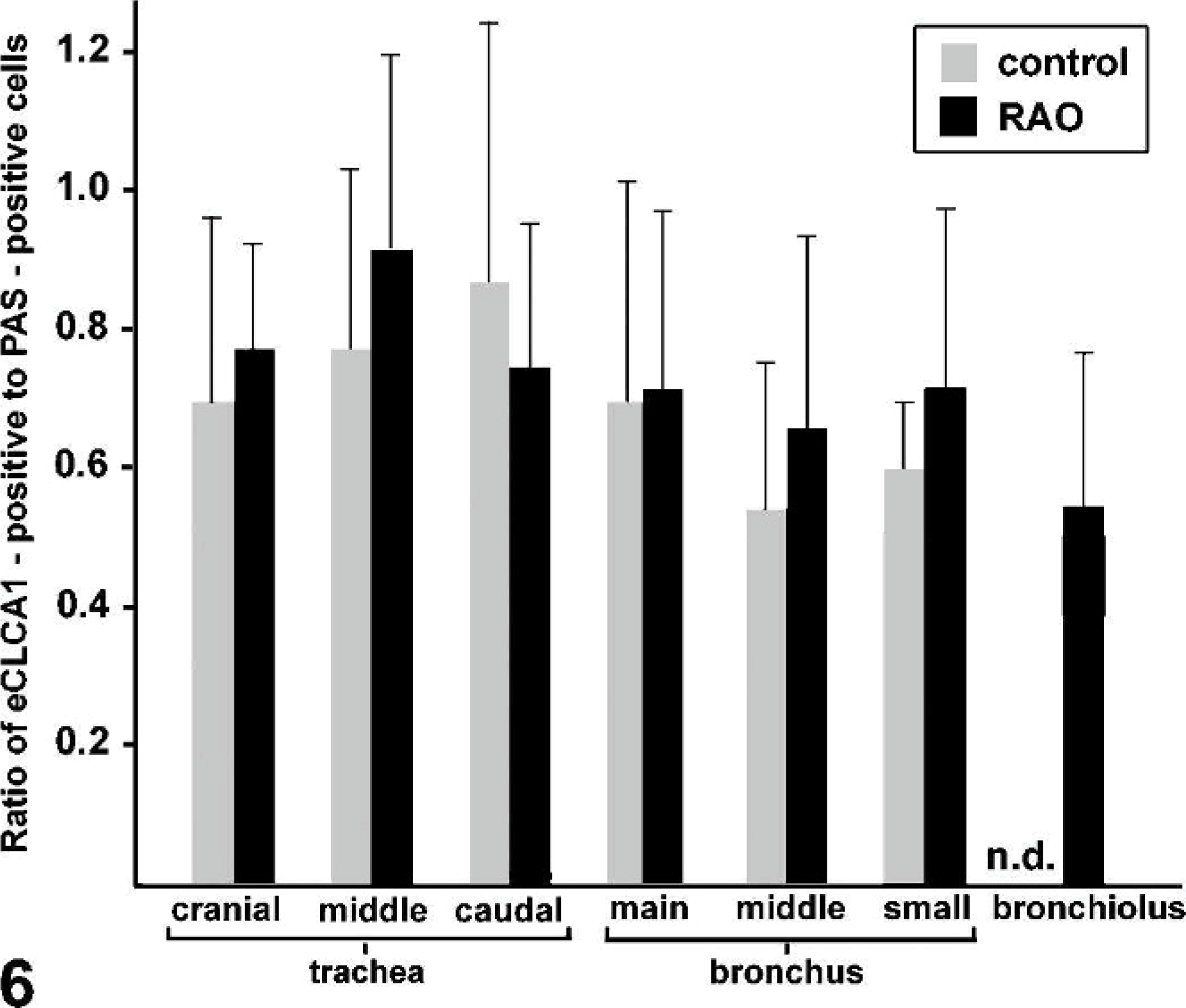
The ratios between the percentages of eCLCA1 expressing cells and PAS-positive cells were calculated in control and horses affected with RAO. No statistically significant differences were observed between the groups in any of the locations analyzed. Columns and bars indicate mean values and standard deviations, respectively. n.d., not doable because of the absence of goblet cells.
Relative expression levels of eCLCA1 mRNA in goblet cells
To test whether the cellular expression levels of eCLCA1 were differentially regulated in horses affected with RAO compared with unaffected horses, the relative eCLCA1 mRNA copy numbers per mRNA copy numbers of the housekeeping gene EF-1a 9 were determined in goblet cells by using real-time quantitative reverse transcription polymerase chain reaction (PCR) after laser microdissection. The relative copy numbers of eCLCA1 cDNA per goblet cell were virtually identical between the different localizations analyzed in each horse. Moreover, no statistically significant differences were recorded in relative eCLCA1 mRNA expression per goblet cell among control horses and horses affected with RAO (Fig. 7).
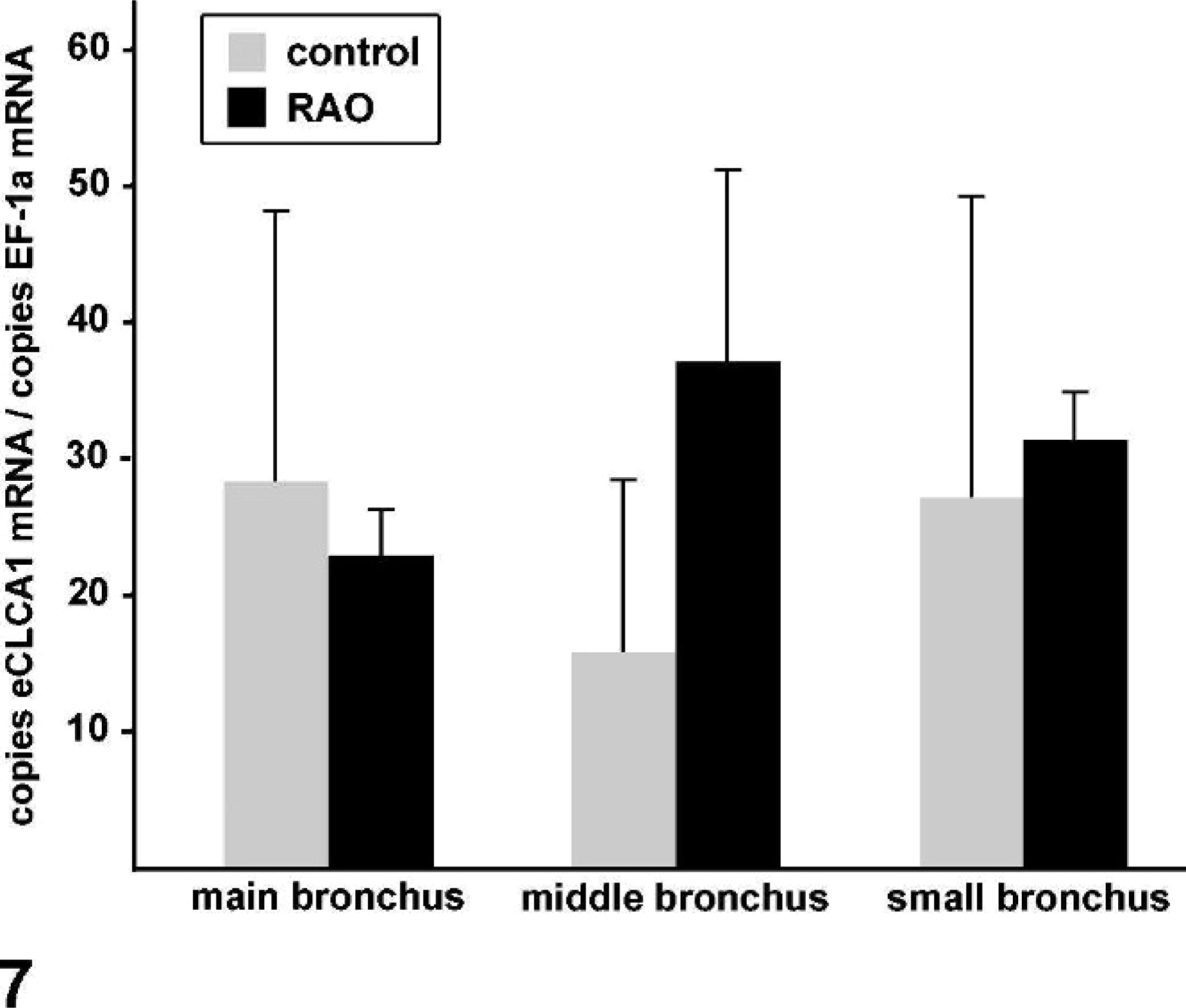
Quantification of eCLCA1 mRNA copy numbers per mRNA copy numbers of the housekeeping gene EF-1a 9 in the bronchi of control and horses affected with RAO by using real-time quantitative reverse transcription PCR following laser microdissection of individual goblet cells or small groups of goblet cells. No statistically significant differences were observed between control and horses affected with RAO. Columns and bars indicate mean values and standard errors of the mean, respectively.
Localization of the eCLCA1 protein
Although initially thought to form transmembrane ion channels, some members of the CLCA family of proteins were recently shown to be secreted, soluble proteins that may regulate ion secretion indirectly rather than acting as channel proteins on their own. 7, 18 In this study, immunohistochemical detection of the eCLCA1 protein did not only stain goblet cells but also strongly labeled accumulated mucins in the lumen of small bronchioles in horses affected with RAO (Fig. 8A). Similar mucin labeling for eCLCA1 expression was sporadically observed in normal horses (Fig. 4B). In addition, immunohistochemical detection of eCLCA1 on smears of tracheobronchial aspirates from horses affected with RAO strongly stained virtually all mucin components but no cells (Fig. 8 B). These findings resulted in the hypothesis that eCLCA1 may, at least in part, be secreted with other goblet cell products.
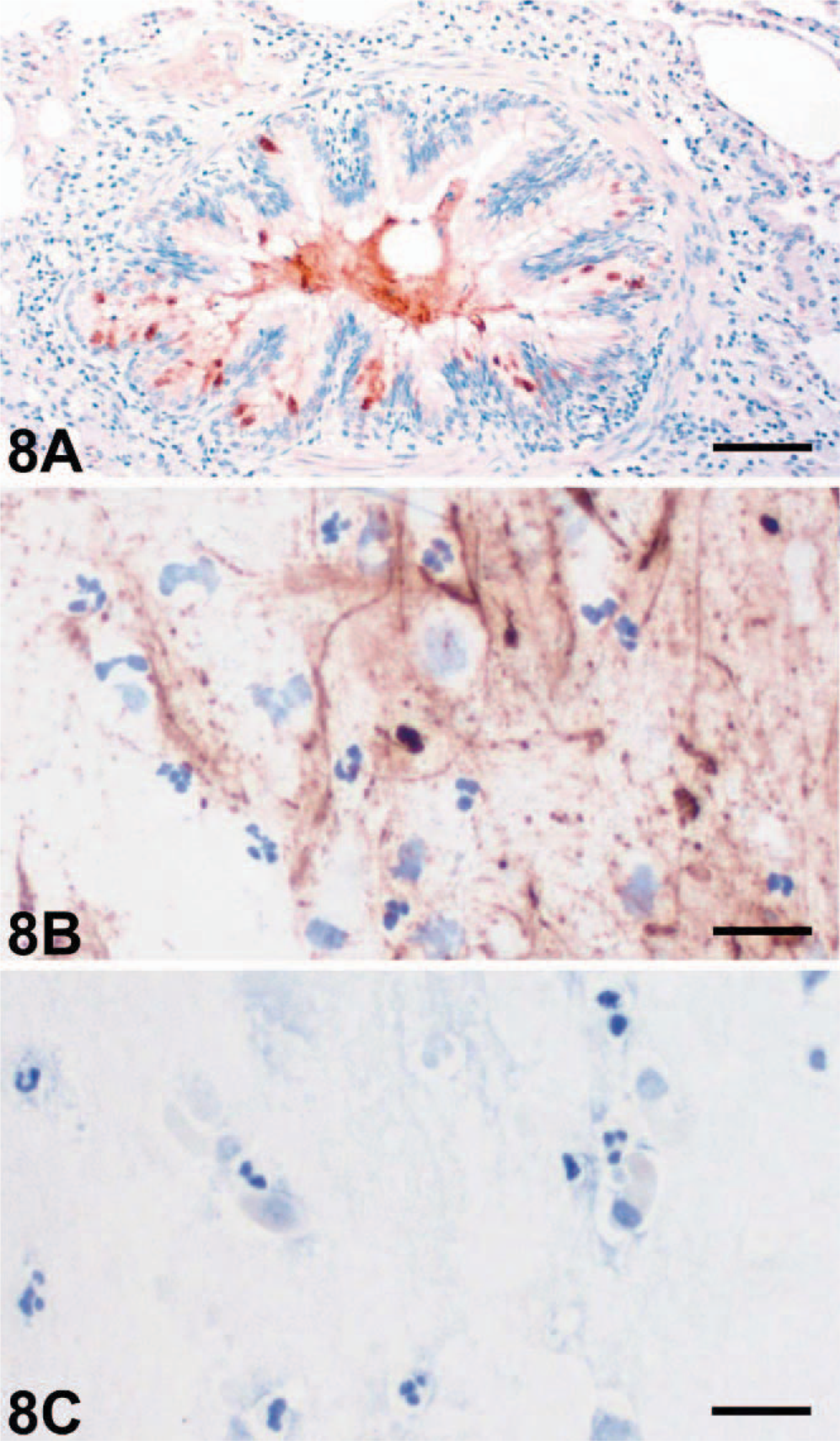
Lung, bronchiole (
To test this hypothesis, we first analyzed the primary eCLCA1 913 amino acid sequence for potential membrane spanning hydrophobic domains by using the Kyte-Doolittle transmembrane prediction program. 13 This program failed to identify any significant hydrophobic domain that could constitute a transmembrane domain other than the signal sequence that targets the polypeptide to the secretory pathway of the cell (Fig. 9A).
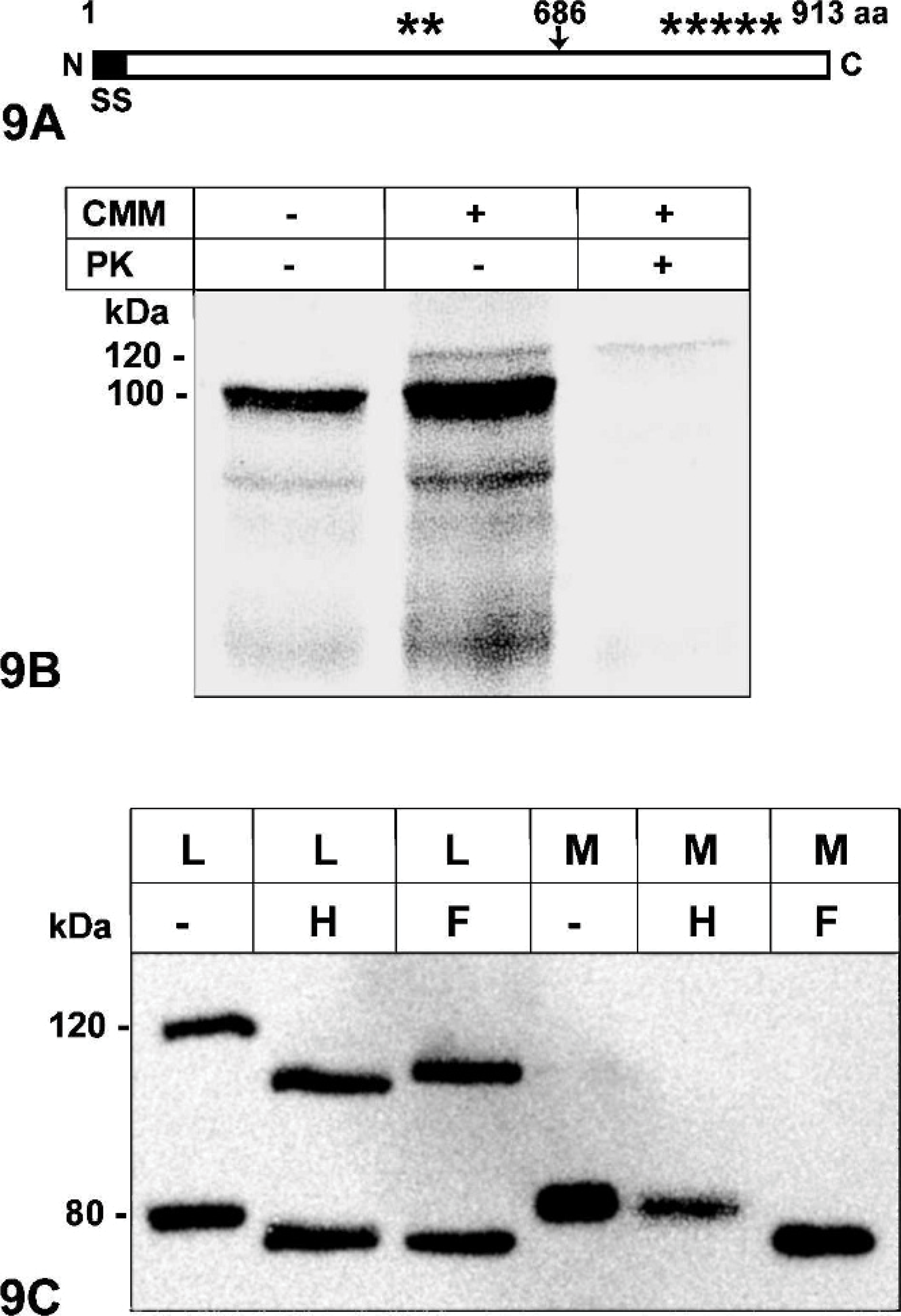
Characterization of the eCLCA1 protein by sequence analysis (
A protease protection assay was further used to identify the size of extracellular fragments of the eCLCA1 proteins. First, in vitro translation of the polypeptide resulted in an approximately 100 kDa primary translation product, consistent with the predicted molecular weight of the 913 amino acid chain (Fig. 9B). The addition of sugar residues by canine microsomal membranes increased the size of the protein to approximately 120 kDa, consistent with 7 predicted sites for asparagine-linked glycosylation (see asterisks in Fig. 9A). Further addition of proteinase K that degraded all proteins not translocated into the microsomal vesicles corresponding to the extracellular space resulted in a complete loss of all protein bands except for the 120 kDa band. This observation clearly indicated that the entire polypeptide chain entered the secretory pathway without any transmembrane domain.
To test whether the mature eCLCA1 glycoprotein was indeed secreted by living cells, the cloned eCLCA1 cDNA was transfected into cultured HEK293 cells and Western blot analyses were performed to characterize the glycosylation pattern in cell lysates and in the supernatant, i.e., the tissue culture medium. Western blot analysis of cell lysates identified the 120 kDa glycosylated primary translation product and the 80 kDa amino terminal cleavage product (Fig. 9C), which is thought to represent the functional protein. 1 Removal of immature, high-mannose–type sugar residues added in the endoplasmic reticulum by endoglycosidase H reduced the sizes to approximately 100 kDa and 75 kDa, respectively, indicating that both protein forms are still immature. Additional incubation with PNGase F that removed both high-mannose–type and mature complex glycoresidues also reduced the sizes of the 2 proteins. These results suggested that the 2 proteins had not passed the Golgi apparatus, and no mature protein was associated with the cells, including the cell-surface membranes. When the medium was tested 48 hours after transfection, only the 80 kDa protein was found secreted by the cells but not the primary translation product. However, the 80 kDa protein was resistant to endoglycosidase H treatment and only susceptible to PNGase F treatment, indicating that this was the mature, complex glycosylated protein after final processing in the Golgi apparatus.
Discussion
Obstruction of small airways by hypersecreted mucins is among the key mechanisms that lead to impaired respiratory function in horses suffering from RAO. 2 Recent studies identified the human hCLCA1 and its murine orthologue, mCLCA3, as key mediators of the goblet cell hyperplasia observed in both asthmatics and in mouse models of asthma. 10, 12, 18, 22, 24 Several issues, however, remained unclear, including the questions of whether these CLCA proteins are upregulated on the transcriptional level and whether these proteins are genuine channel proteins or regulators of other molecules. The equine orthologue of hCLCA1 and mCLCA3, eCLCA1, was identified and cloned recently, and was shown to be strongly upregulated in the lungs of horses suffering from RAO. 1
The results of this study demonstrate that the eCLCA1 protein was expressed in the hyperplastic mucin-producing goblet cells throughout all airway segments in horses with RAO. Thus, the increased numbers of eCLCA1 mRNA molecules in total lungs of horses with RAO 1 could either be because of transcriptional upregulation of the protein on the cellular level or merely because of increased numbers of eCLCA1 expressing cells. This distinction is critical for understanding the role of CLCA proteins either as primary regulators of goblet cell hyperplasia and mucin production, as suspected earlier, 18, 24 or as bystander molecules, secondary to goblet cell hyperplasia. Relative quantification of eCLCA1 mRNA copy numbers in goblet cells by using laser-capture microdissection and quantitative reverse transcription PCR failed to identify differential expression of eCLCA1 in goblet cells of horses with RAO compared with unaffected horses. Moreover, immunohistochemical detection of eCLCA1 failed to identify the protein in cell types other than goblet cells under conditions of RAO. Taken together, these data clearly show that increased numbers of eCLCA1 mRNA in lungs of horses with RAO 1 is a bystander effect, secondary to goblet cell hyperplasia and not because of transcriptional upregulation in goblet cells or other cell types. This is consistent with previous findings in mouse models of cystic fibrosis with intestinal mucus obstruction where the increase of mCLCA3 mRNA copy numbers was very similar to the level of goblet cell hyperplasia. 16 Nevertheless, the more than twofold increase of goblet cells expressing eCLCA1 under conditions of equine RAO and a similar increase in goblet cells expressing mCLCA3 in cystic fibrosis mice may still be consistent with a significant functional role of this molecule in RAO or cystic fibrosis, respectively. However, it appears less obvious how vector-induced overexpression of the murine and human orthologues leads to goblet cell hyperplasia and their antisense-suppression results in reduced numbers of goblet cells. 10, 18
As a noteworthy methodological aspect, the results of this study show that gene-expression profiling by using whole-tissue extracts may be misleading in terms of identifying different transcriptional expression levels of particular genes. Our previous observations on increased copy numbers of eCLCA1 mRNA in lungs of horses with RAO had initially been interpreted as true upregulation of the gene, 1 which, in this study, turned out to be a misinterpretation. Clearly, the number of cells expressing the gene of interest has to be taken into account with caution when mRNA copies are quantified in a tissue of interest, particularly when this number changes under condition of disease. Laser-capture microdissection of single cells or small groups of cells, followed by quantification of mRNA copy numbers relative to a housekeeping gene has become a valuable tool to evade this problem.
Initial observations on the immunostaining of secreted mucins for eCLCA1 prompted us to biochemically test whether the eCLCA1 protein was at least in part secreted from the cell. Both experimental approaches, the protease protection assay, as well as the characterization of glycosylation patterns in cell lysates and tissue-culture medium unequivocally revealed that the protein is entirely secreted by the cells without any detectable association with the plasma membrane. Furthermore, the complex glycopattern of the fully processed protein was detectable only in the supernatant of the cultured cells and not within the cell lysates, including the plasma membrane. These observations were consistent with the previous electron immunolocalization of the orthologous murine protein, mCLCA3 alias gob-5, in secretory vesicles of goblet cells and in the secreted mucus layer in mouse intestines. 15 Moreover, the results on eCLCA1 in this study are very similar to the results of a more recent systematic biochemical analysis of the mCLCA3 protein, revealing that mCLCA3 is entirely secreted without any association with the cell membrane. 17 Thus, eCLCA1 seems to resemble its murine orthologue, mCLCA3, more closely than its human orthologue, hCLCA1, which is linked to the outer plasma membrane, probably via binding to an as yet unidentified second protein. 7 Whether this significant difference between the equine and human orthologues of the CLCA gene family are of relevance with regard to differences between certain secretory disorders in the 2 species remains to be shown. Clearly, because of its secreted nature eCLCA1 cannot act as a channel protein on its own and the chloride secretory response that is associated with expression of eCLCA1 1 is much more likely because of an indirect effect, possibly via regulatory functions of this protein on other ion channels.
Taken together, the results of this study indicate that eCLCA1 is a fully secreted soluble protein that is similarly secreted by goblet cells, both in normal equine airways and in hyperplastic goblet cells in RAO without any differential expression on the level of transcriptional regulation. These results, together with recent reports on hCLCA1 7 and mCLCA3, 7, 17 rectify the previously hypothesized universal model of CLCA proteins being transmembrane proteins with genuine channel function. Still, the clearly increased expression of this protein may play an important role in RAO airways as a mediator of chloride secretion or other goblet cell functions via as yet unknown pathways. Several other soluble constituents of the mucin layer are thought to act as signaling molecules on epithelial cells, such as the trefoil factor family proteins that also act on secretory pathways. 5 It will be challenging to identify molecules that interact with eCLCA1 and to explore the mechanism of how this group of CLCA proteins exerts its seemingly well-established regulatory role in goblet cell hyperplasia and mucin secretion. 10, 18, 22– 24 Only with these data at hand will it be reasonable to assess any potential diagnostic or therapeutical value of eCLCA1 in equine RAO.
Footnotes
Acknowledgement
This study was supported by funds from the German Research Council (Deutsche Forschungsgemeinschaft grant GR1491/3).