
Abstract
The purpose of this study was to determine by immunohistochemistry the expression of estrogen and progesterone receptors in ovarian follicular structures from cows with cystic ovarian disease (COD) and to compare these with normal ovarian structures. Secondary, tertiary, atretic, and cystic follicles were evaluated. The follicular cysts of animals with COD presented a significantly higher expression of estrogen receptor α in all follicular layers than secondary, tertiary, and atretic follicles in both groups (P < .05). The intensity of estrogen receptor β in the granulosa cell layer was stronger in tertiary than in secondary and atretic follicles in normal animals (P < .05) and in growing and cystic follicles in animals with COD (P < .05). Theca cells were scarcely stained in the 2 groups. Growing follicles and cysts from COD animals were less stained than tertiary follicles from normal animals (P < .05). Differences did not exist between the 2 groups with regard to the progesterone receptor. Ovaries of animals with COD exhibited altered estrogen receptors expression compared with that in normal animals.
Keywords
Cystic ovarian disease (COD) in dairy cattle is defined as the presence of 1 or more ovarian follicular structures measuring at least 20 mm in diameter, persisting in the ovary for 10 days or more in the absence of corpus luteum (CL), and interrupting the normal reproductive cycle.18 The pathogenesis of COD is a complex process that involves dysfunction in ovulation. The primary cause of this disease has not been elucidated.7,14 Receptors for estrogen (ER) are expressed as 2 structurally related subtypes in mammals, ERα and ERβ, which are encoded by 2 distinct genes. The existence of these 2 subtypes may partly explain the selective action of estrogen in different target tissues and in the same tissue in different physiological states.5 Studies on several species have confirmed the differential distribution of these 2 receptors in the ovary: ERβ was detected mainly in granulosa cells, whereas ERα was localized in theca cells, interstitial glands, stromal cells, and germinal epithelium.1,2,6,15,20 The progesterone receptor (PR) has at least 3 isoforms, all originating from the same gene.3,11 In bovine tissues, all 3 isoforms (PR-A, PR-B, and PR-C) have been detected.17 The PR-A and PR-C isoforms may have an inhibitory influence on the transcriptional activity of PR-B.5,11
It is well known that folliculogenesis is a process in which the strait equilibrium in hormonal secretion at the hypothalamic-pituitary-gonadal axis level allows the normal development of the reproductive cycle in females. Any factor that alters this delicate balance could lead to alterations in fertility, leading to anestrous, silent estrous, and ovarian cysts.14,18 The purpose of the present study was to localize and to quantify by immunohistochemistry the expression of ER and PR in ovarian follicular structures from cows with cystic ovarian disease and to compare it with the expression in ovarian structures from cows with regular cycles.
Ovaries with cystic follicles (n = 10) were collected at a local abattoir in Rafaela city (Santa Fe, Argentina), within 20 minutes of death, from mixed breeds of Bos taurus cows, assessed visually as being nonpregnant. Follicular cysts were diagnosed when the follicles were more than 20 mm in diameter, in the absence of a functional CL in either the right or the left ovary.18
Four nulliparous Argentinean Holstein heifers with regular estrous cycles were used as normal controls. Estrous cycles were synchronized by Ovsynch protocol, and the time of first detection of estrous behavior was designated as Day 0 of the cycle.13 Ovarian ultrasonographic examinations were performed, and the ovaries were removed surgically by flank laparotomy, when the dominant follicle reached a diameter of >10 mm, in the absence of an active CL (Day 18 approximately).19
The ovaries were conditioned, fixed in 10% buffered formalin for 8 hours at 4°C, and then washed in phosphate-buffered saline; processed in the routine manner; and embedded in paraffin. Serial sections 5 μm in thickness were mounted on 3-aminopropyl triethoxysilane (Sigma, USA)-coated slides and dried for 24 hours at 37°C.16
Follicles were classified according to the criteria listed in the Nomina Histologica into the following groups: secondary, tertiary, atretic, and cystic follicles.12 Tertiary follicles containing less than 1% apoptotic cells were regarded as vital tertiary follicles, whereas follicles containing more than 1% apoptotic cells were classified as atretic.6 The latter were further classified as obliterative or cystic.10
An indirect immunohistochemistry method was used. Antibody against ERα (clone 1D5) was purchased from Cell Marque (Santa Bárbara, CA, USA); antibodies against ERβ (polyclonal) and PR (clone PR88) were purchased from BioGenex (San Ramón, CA, USA). The secondary antibody goat polyclonal anti-mouse IgG was purchased from Zymed (San Francisco, CA, USA), and goat polyclonal anti-rabbit IgG was purchased from Chemicon (Temecula, CA, USA). Each primary antibody was assayed in at least 5 sections of each ovary from each individual. A streptavidin-biotin immunoperoxidase method was used as described earlier.16 In brief, sections were deparaffinized and hydrated, and then microwave pretreatment (antigen retrieval) was performed. Endogen peroxidase activity was inhibited with 1% H2O2, and nonspecific binding was blocked with 10% normal goat serum. All sections were incubated with primary antibodies for 18 hours at 4°C, and after being washed in phosphate-buffered saline, the samples were incubated for 30 minutes at room temperature with preabsorbed biotinylated secondary antibodies selected specifically for each of the 2 types of primary antibodies used (monoclonal or polyclonal). The visualization of antigens was achieved by the streptavidin-peroxidase method (BioGenex, San Ramon, CA, USA), and 3.3-diaminobenzidine (Liquid DAB-Plus Substrate Kit—Zymed, San Francisco, CA, USA) was used as chromogen. Finally, the slides were washed in distilled water and counterstained with Mayer's hematoxylin, dehydrated, and mounted. Positive control tissues were used. Negative control sections were subjected to the same immunohistochemical method, replacing primary antibodies by rabbit and mouse nonimmune serum.
Image analysis was performed using an Image Pro-Plus 3.0.1 system (Media Cybernetics, Silver Spring, MA, USA). Images were digitized by a CCD color video camera (Sony, Montvale, NJ, USA) mounted on a conventional light microscope (Olympus BH-2, Olympus Co., Japan), using an objective magnification of ×40. Briefly, using a color-segmentation analysis tool, the total positively stained nuclear area (brown reaction product) was measured and expressed as a ratio (%) of the total area of cell nuclei (brown reaction product + blue hematoxylin). The image analysis score was calculated separately in each follicular wall layer (granulosa, theca interna, and theca externa) from at least 50 images of the following structures: secondary, tertiary, atretic, and cystic follicles from normal and COD ovaries.
The results obtained were analyzed with de SPSS software (version 11.0 for Windows, SPSS Inc., Chicago, IL, USA). All the data were expressed as the mean ± standard error of mean. The percentage of positivity for each antibody in different areas and structures was analyzed by means of an ANOVA test and a Duncan post test.
Morphologically, ovaries from normal cows exhibited follicles in various stages of development, including primary follicles, secondary follicles, tertiary follicles, corpora albicantia, and late CL, as well as follicles with different degrees of atresia. In cows with COD, the development of healthy secondary and tertiary follicles could be observed, in addition to others showing evidence of atresia and 1 or 2 large cystic structures with a thickened or absent granulosa cell layer. A CL was not present in any of these cases. The cystic follicles showed no signs of luteinization.
A summary of the immunohistochemical expression of different antibodies is given in Tables 1, 2, and 3. The reactions were negative in all negative controls and positive in the positive tissue controls.
Image analysis score for ERα immunoexpression measured as ratio (%) of positively stained nuclear area (brown reaction product) on the total area of cell nuclei (brown reaction product + blue hematoxylin) in the follicular wall of normal and cystic bovine ovaries.
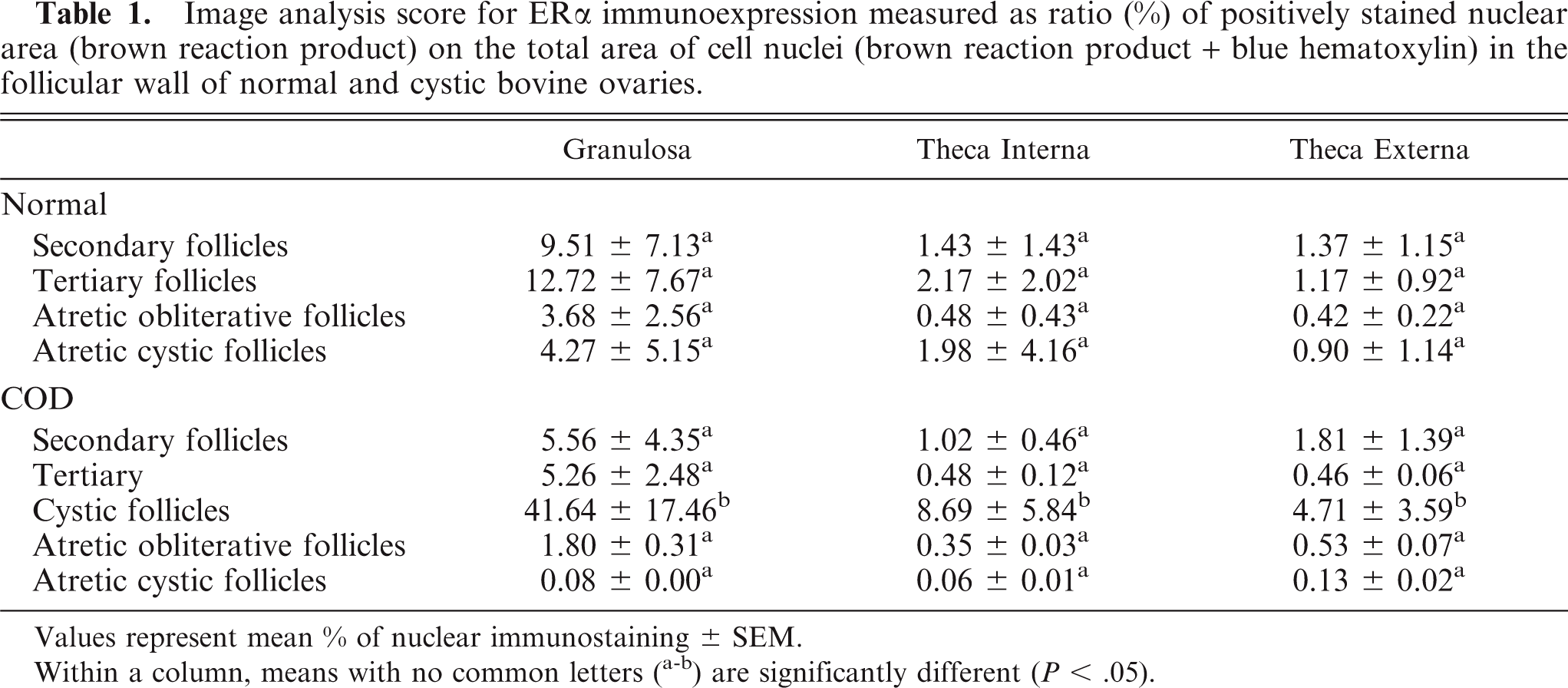
Values represent mean % of nuclear immunostaining ± SEM.
Within a column, means with no common letters (a-b) are significantly different (P < .05).
Image analysis score for ERβ immunoexpression measured as ratio (%) of positively stained nuclear area (brown reaction product) on the total area of cell nuclei (brown reaction product + blue hematoxylin) in the follicular wall of normal and cystic bovine ovaries.
Values represent mean % of nuclear immunostaining ± SEM.
Within a column, means with no common letters (a-b-c) are significantly different (P < .05).
Image analysis score for PR immunoexpression measured as ratio(%) of positively stained nuclear area (brown reaction product) on the total area of cell nuclei (brown reaction product + blue hematoxylin) in the follicular wall of normal and cystic bovine ovaries.
Values represent mean % of nuclear immunostaining ± SEM.
Within a column, means with no common letters (a-b) are significantly different (P < .01).
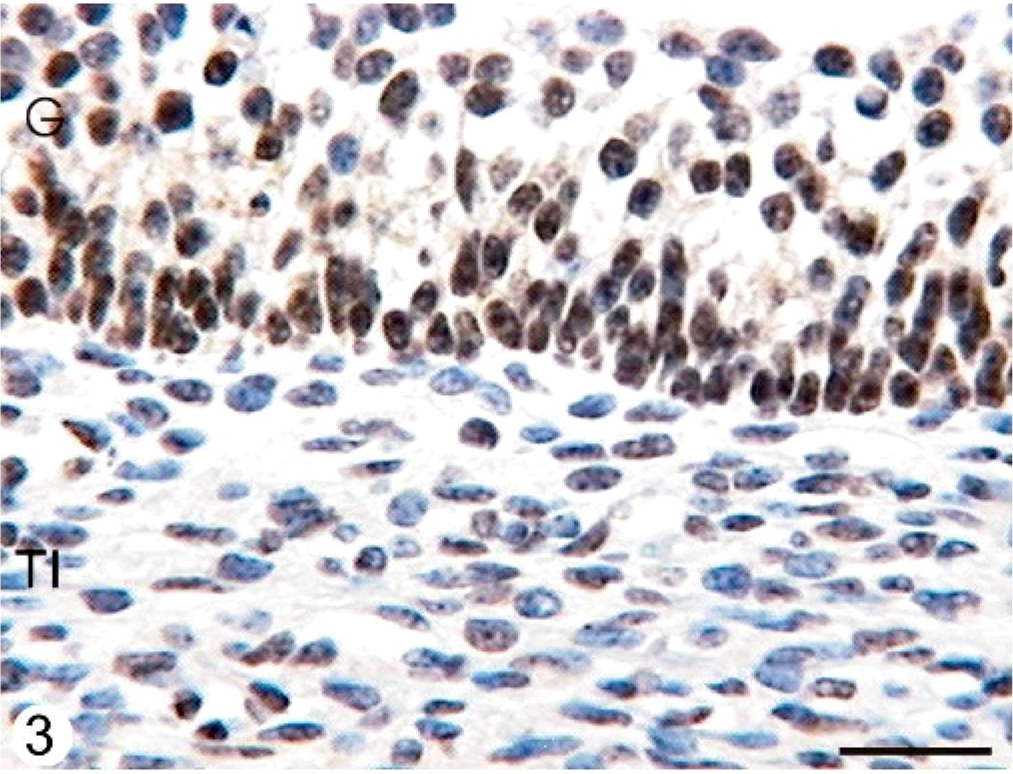
ERα was expressed in the cellular nuclei and cytoplasm of the granulosa, theca interna, and theca externa layers from all follicular categories studied. Granulosa cells were moderately stained with 1D5 clone. The reaction products in this cell type were granular in the nucleus. They were diffuse in the cytoplasm. ERα was widely dispersed in the theca interna and showed moderate staining intensity with homogeneous distribution in the cellular nucleus. The immunostaining was low in the theca externa. There were differences in the expression between normal animals and animals with COD. The follicular cysts presented a significantly higher expression of this protein in the nuclei than in the secondary, tertiary, and atretic follicles in both groups (P < .05) (Figs. 1, 2, Table 1).
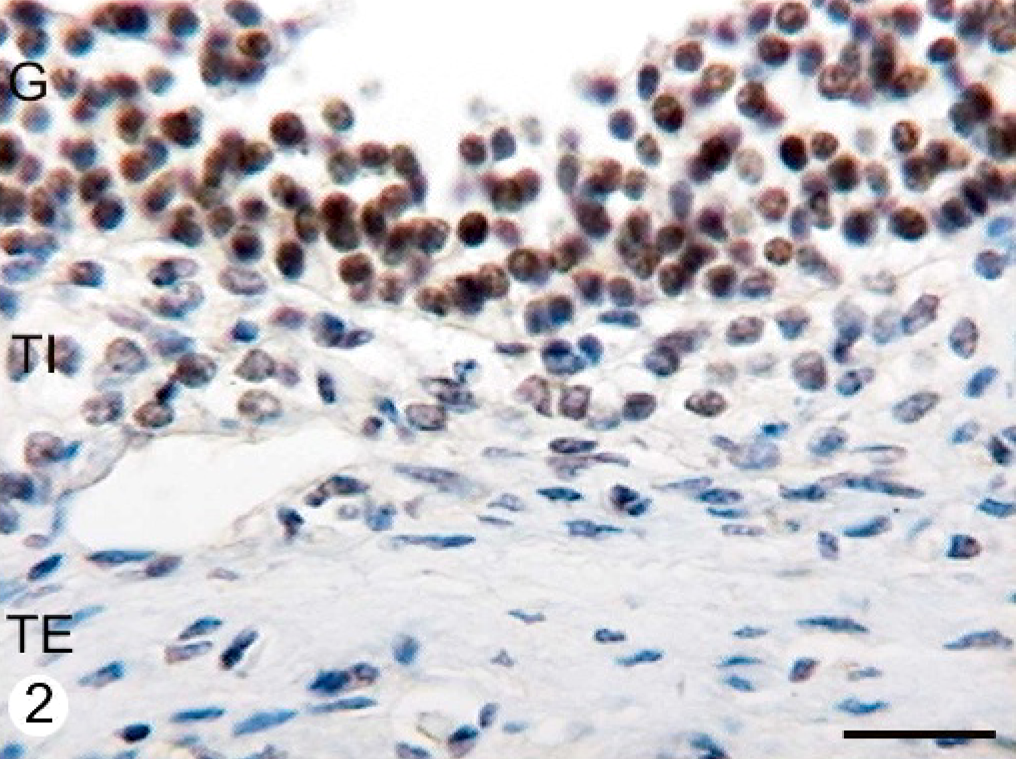
Immunohistochemical staining with ERα antibody in the follicular wall of a cyst from a COD bovine ovary. A high number of granulosa cells show an intense granular immunoreaction with this antibody, whereas some theca interna cells show moderate or weak diffuse immunostaining. Streptavidin-Biotin method, Mayer's hematoxylin counterstain. G = granulosa; TI = theca interna; TE = theca externa. Bar = 25 μm.
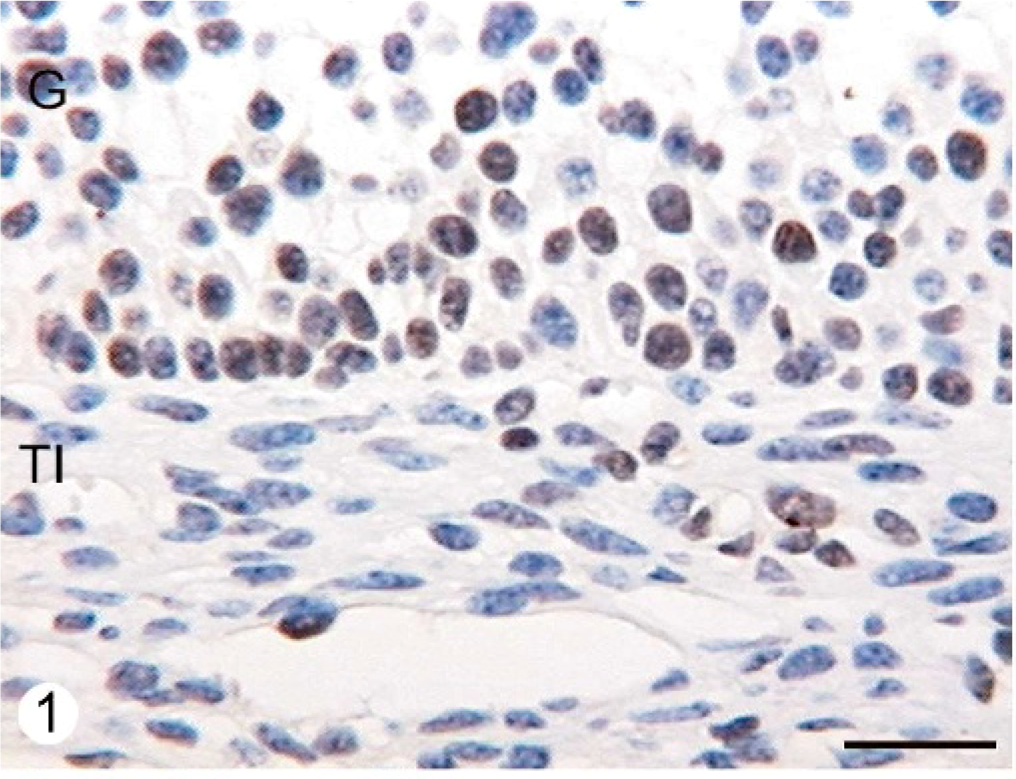
Follicular wall of a healthy tertiary follicle from a normal bovine ovary immunostained with ERα antibody. Some granulosa cells shows moderate nuclear staining, whereas others shows weak or no staining. Theca interna show a few nuclei with light reaction to the antibody. Streptavidin-Biotin method, Mayer's hematoxylin counterstain. G = granulosa; TI = theca interna. Bar = 25 μm.
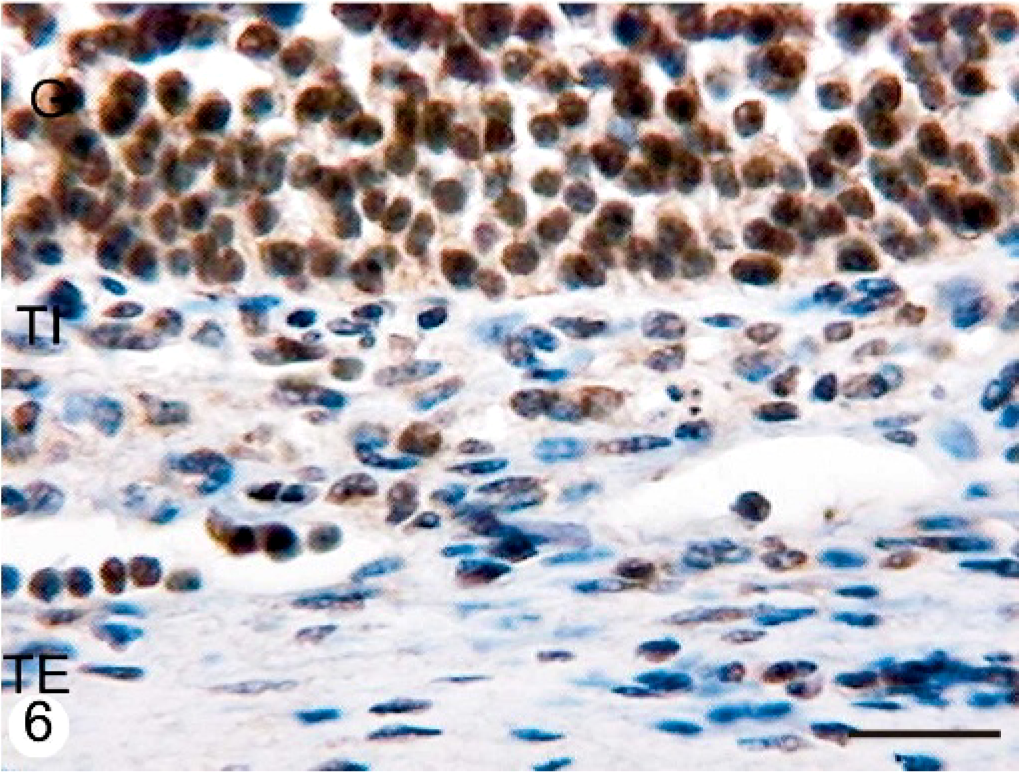
ERβ protein was detected in the nuclei of granulosa, theca interna, and theca externa of all follicles studied, although the intensity of ERβ in the granulosa cell layer was stronger in tertiary follicles than in secondary and atretic follicles in normal animals (P < .05). The reaction product was granular in the nuclei of all follicular layers. In animals with COD, the immunoexpression of ERβ diminished in all growing follicles, inclusive in cystic follicles. Theca cells (both interna and externa) were scarcely and diffusely stained in the 2 groups, but all growing follicles and cysts from animals with COD were less stained than tertiary follicles from normal animals (P < .05) (Figs. 3, 4, Table 2). Diffuse cytoplasmic immunoreactivity was also observed in the granulosa cells of the follicles.
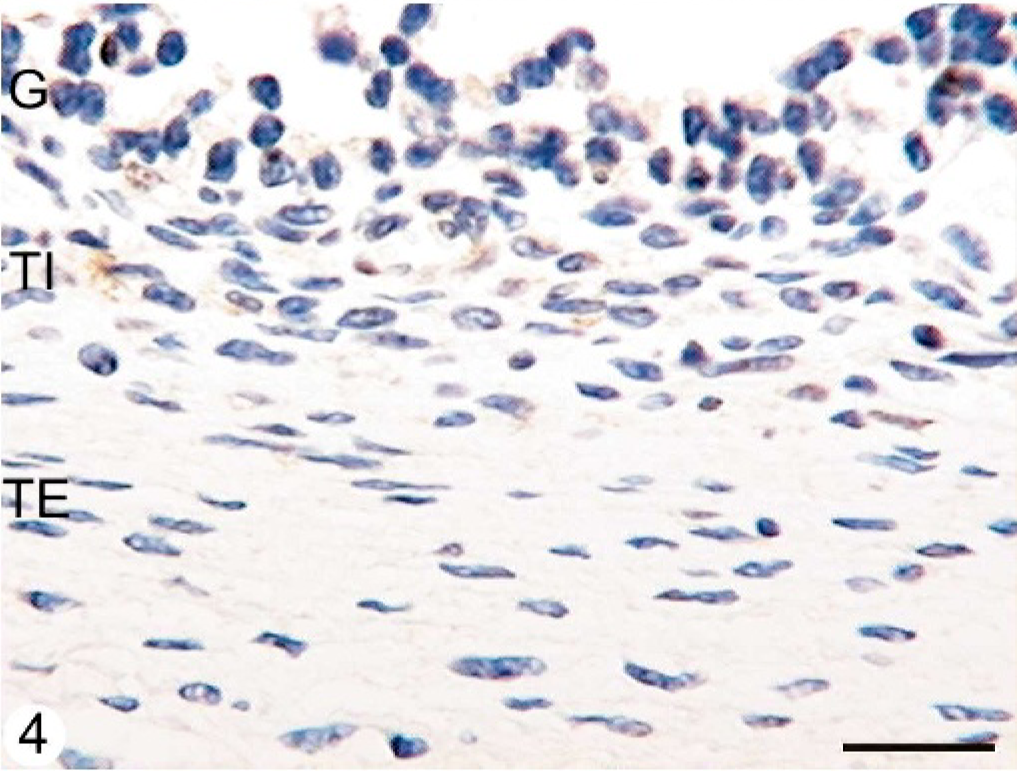
Follicular wall of a cystic follicle of an animal with COD. A very scarce proportion of granulosa and theca cells show a weak immunoreaction to the antibody anti-ERβ. Streptavidin-Biotin method, Mayer's hematoxylin counterstain. G = granulosa; TI = theca interna; TE = theca externa. Bar = 25 μm.
ERβ immunostaining in the wall of a tertiary follicle from normal bovine ovary. Granulosa cell layers show intense granular nuclear staining in a high percentage of cells. Granular and intense reaction can be also observed in theca cells. Streptavidin-Biotin method, Mayer's hematoxylin counterstain. G = granulosa; TI = theca interna. Bar = 25 μm.
PR protein was observed in the nuclei of different ovarian cell groups, namely in granulosa, theca interna, and theca externa cells from secondary, tertiary, atretic, and cystic follicles. The nuclei showed a granular reaction in the 3 studied follicular layers. Diffuse cytoplasmic immunostaining was observed in the granulosa cells of the 2 groups. The expression of PR was lower in the granulosa cell layer from atretic cystic follicles of COD and normal ovaries than in tertiary follicles from normal ovaries (P < .01) (Figs. 5, 6, Table 3).
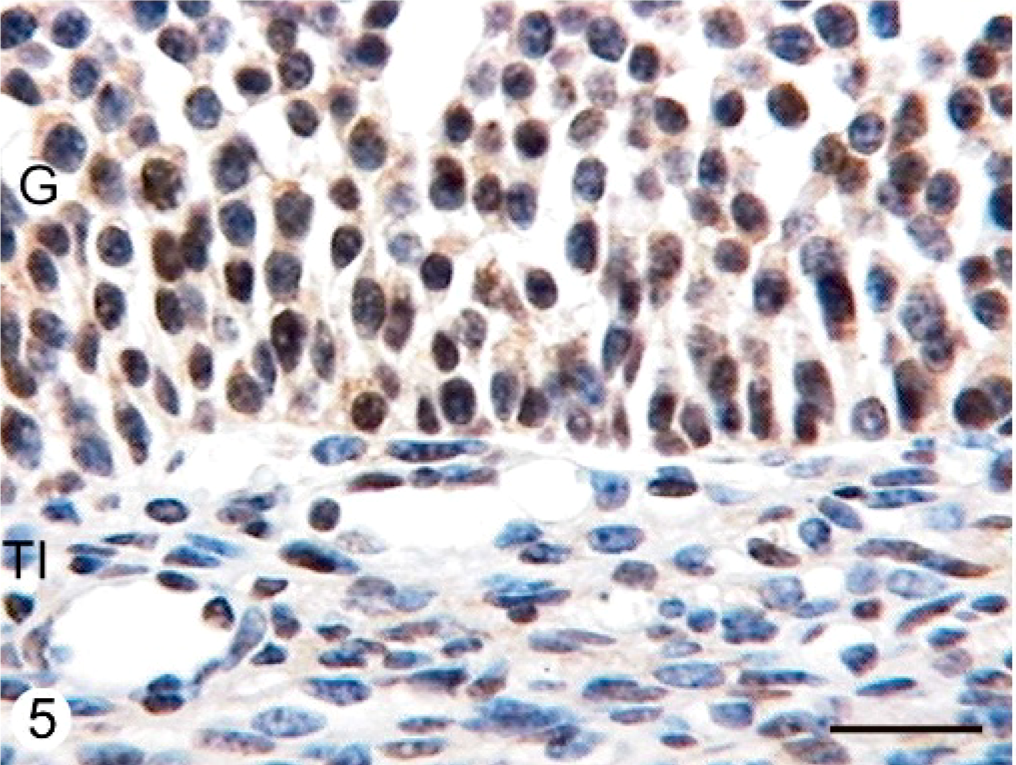
Microphotographs showing the staining with the antibody anti-PR in the follicular
wall of a tertiary follicle of a normal ovary (
Microphotographs showing the staining with the antibody anti-PR in the follicular
wall of a tertiary follicle of a normal ovary (
We observed a decrease in the expression of ERβ in growing, atretic, and cystic follicles in animals with COD compared with tertiary follicles from normal animals. At the same time, we also found a high expression of ERα in cystic follicles in relation to other follicular categories. The decrease in the levels of ERβ in the follicles of animals with COD could be due to high circulating levels of luteinizing hormone that usually are present in animals with this illness. It is known that gonadotropins and estrogen downregulate granulosa expression of this ER isoform. Both ERs show a tendency to upregulate together with increasing estrogen levels in the follicular fluid, and this upregulation is correlated to an upregulation in the luteinizing and follicle-stimulating hormone receptors.2 The change in the concentration of different types of receptors in granulosa and theca cells of follicles from animals with COD could alter the ERα/ERβ ratio, causing modifications in the action or effects of estrogen on its target cells. Because ERα and ERβ have different sensitivities to estradiol and because of the possibility of cross-talk between receptors or for heterodimer formation to occur, small modifiers in the ERα/ERβ relation may perturb normal folliculogenesis.15,21 Then, the effects of the estradiol could be an alteration in the balance of proliferation/apoptosis, an alteration in the expression of gonadotropins receptors, or a disturbance of enzyme action and metabolism.4,8,9
The distribution and localization of PR in the ovary remained without significant changes in animals with and without the disease. Because the antibody used by us could not distinguish between isoforms, the selective detection of diverse localization of PR-A, PR-B, or PR-C was not possible. In vitro studies have shown that the A and B forms may have different functions in the same cell and also that the activity of the individual isoform of the receptors may vary among distinct cell types.3,11
In summary, ovaries from animals with COD exhibited altered ER expression compared with normal ovarian structures of synchronized animals. It is reasonable to suggest that in conditions characterized by altered ovulation, such as COD, changes in the expression of ovarian ERs could play a role. More studies are necessary to fully understand and appreciate the implications of these observations.
Footnotes
Acknowledgements
We are very grateful to the Meat Inspection Office in Rafaela Alimentos (Rafaela, Santa Fe, Argentina) for supplying us with bovine ovaries and to the members of the Animal Health Hospital (FCV-UNL) for the animal care. This study was supported by grants from the Argentine National Agency for the Promotion of Science and Technology (ANPCyT) (BID OC-AR 1201—PICT 08-14734). NRS is a Fellow and HHO and EJG are Research Career Members of the National Scientific Research Council (CONICET).