
Abstract
The progression of tracheal lesions induced by vaccination of day-old specific pathogen-free chicks with the La Sota strain of Newcastle disease virus (NDV) was examined by relating surface changes as observed by scanning electron microscopy with subcellular changes seen by transmission electron microscopy. NDV infection resulted in hypertrophy of goblet cells, their rupture, and the formation of excess mucus. Activation of goblet cells peaked within 4 days postvaccination. Afterward, the activation levels gradually decreased. At the level of the ciliated cells, a marked increase in the proportion of nonciliated to ciliated cells and later an almost complete deciliation of the tracheal surface were observed because a simple squamous to cuboidal epithelium replaced the original pseudostratified epithelium. Fifteen days postvaccination, all epithelial damage was restored. Because the observed vaccination-induced lesions are detrimental to epithelial integrity and function as a barrier against invading microorganisms, they might explain at the ultrastructural level the secondary complications of vaccination with the La Sota strain against NDV
Newcastle disease (ND) is a highly contagious viral disease of poultry and other bird species caused by specified viruses of the avian paramyxovirus type I (APMV-I) serotype of the genus Avulavirus belonging to the family Paramyxoviridae. 14 The virus has a wide host range, and >250 species of birds have been reported as susceptible to infection. 2 NDV isolates are classified into forms or pathotypes of disease on the basis of clinical signs in chickens. 4 The strains of low virulence can only replicate in areas with trypsinlike enzymes, such as in the respiratory and intestinal tract, whereas the virulent viruses can replicate in a range of tissues and organs. Infection with NDV strains of high virulence hence frequently results in a fatal systemic infection, whereas strains of low virulence cause transient damage to the respiratory and intestinal epithelia. The latter affects the role of the epithelium as a barrier against invading microorganisms and is one of the causes of vaccination-associated secondary complications. 1, 5, 17
The control of ND is based on complementary hygienic and prophylactic measures. 15 Immunity against ND is usually associated with the presence of neutralizing and hemagglutination-inhibiting (HI) antibodies. The HI test is a practical and reliable serologic indicator of immunity. 3 Vaccination against ND can be performed with either live or inactivated vaccines. Live vaccines induce higher protection and have been used by the poultry industry for >50 years. They are based on the use of lentogenic strains, among which Hitchner B1 and La Sota are the most popular. 2 The efficiency of a live vaccine depends on its potency to multiply enough within the chicken to induce a satisfactory immune response. 15 In general, because they are more invasive, La Sota vaccines give better protection than B1 strains 22 and a have greater tendency to spread from bird to bird within a house. 21 However, because the intensity of the protective immune response is related to virulence of the viral strain, La Sota is more likely to cause a mild to severe respiratory reaction in young birds and to predispose the birds to secondary infections. 2
The progression of NDV-induced tracheal lesions was examined at tissue level by standard histological methods in turkeys exposed to the La Sota strain 1 and in chickens exposed to the B1 strain of NDV. 17 NDV-induced pathological changes at the surface of the tracheal mucosa were studied by scanning electron microscopy (SEM) in chickens 7, 8, 17 and in embryos and their tracheal explants. 9 These changes comprise hypertrophy of the goblet cells, disorientation and deformation of the cilia, deciliation of the epithelium, and plaques of lysed cells.
In this report, we examine whether the NDV La Sota strain–induced tracheal lesions are more severe than the lesions observed for the Japanese Ishii NDV strain, 7 which in phylogenetic analyses clusters together with the Ulster NDV strain, known as an enteric, asymptomatic pathotype. 12 We relate the La Sota NDV–induced changes of the surface of the tracheal epithelium, as observed by SEM with cellular and sub-cellular changes seen by transmission electron microscopy (TEM).
Materials and Methods
NDV virus
The seed stock of the La Sota strain of NDV was obtained from Dr. D. J. Alexander (International Reference Laboratory, Weybridge, UK). This strain was propagated in the allantoic cavity of embryonated specific pathogen–free (SPF) eggs as described. 16
Treatment of chickens
Fertilized eggs of SPF White Leghorn chickens (Valo-CF) were obtained from Lohmann (Cuxhaven, Germany) and incubated at 37.8°C and a relative humidity of ∼55%. Hatched chicks were housed in isolation units under SPF conditions with free access to food and water. One group of 1-day-old SPF chicks was vaccinated intranasally with 106 mean egg infective dose (EID50) of the La Sota strain. A second group of chicks were sham inoculated with PBS and housed in a separate isolator. Serum and the trachea were isolated from three chicks at each time point until 15 days posthatch.
Quantification of NDV-specific immunoglobulin M and G
NDV-specific immunoglobulin (Ig)M and IgG were measured in serum samples by indirect enzyme-linked immunosorbent assay (ELISA). The virus is represented by antigen prepared from the NDV–La Sota strain as described 11 and coated to ELISA plates by overnight incubation with 100 μl of the NDV–antigen solution (1 μg/ml) in coating buffer (pH 9.6) containing 50 mM sodium bicarbonate. After 60 minutes incubation of these plates with the serum sample, bound IgM or IgG was detected with isotype-specific monoclonal antibodies. 6 The latter were detected with a peroxidase-labeled goat anti-mouse total Ig conjugate and TMB substrate (Kirkegaard & Perry Laboratories, Gaithersbury, MD). Between incubations, ELISA plates were washed with washing buffer (pH 7.4) containing 0.15 M NaCl, 0.01 M phosphate, 50 mM EDTA, and 0.1% Tween 20. Absorbance at 510 nm was recorded with a Multiscan MCC ELISA reader. Optical densities of serum samples diluted 800 times for IgG and 200 times for IgM with washing buffer were usually situated in the linear part of the serum dilution curve and considered representative for NDV-specific humoral responses. Two replicates were included for each sample. In addition to the test samples, a positive and a negative control serum were included on each plate at indicated dilutions.
The HI titers of the sera were determined as described. 16
Electron microscopy
The tissue samples were processed for EM analysis by standard methods. Briefly, tissue specimens were fixated in 2.5% glutaraldehyde–2% formaldehyde in 0.1 M cacodylate buffer (pH 7.4) and postfixed in 1% OsO4.
For SEM, the samples were afterwards dehydrated in increasing concentrations of ethanol and, finally, acetone for subsequent critical point drying in carbon dioxide. They were glued with carbon cement on aluminum stubs and coated with gold in a vacuum evaporator. Observations were made with the Philips scanning electron microscope (FEI, Einhoven, The Netherlands). Analog images were converted to digital images (8 bit, 2,800 × 3,488 pixels) with the Orion Frame Grabbing System for SEM (E.L.I. sprl, Brussels, Belgium).
For TEM, fixated samples were stained overnight in a freshly prepared 2% uranyl acetate solution at room temperature and dehydrated in increasing concentrations of ethanol and, finally, in anhydrous propylene oxide. After embedding in Spurr medium, the specimen blocks were trimmed to obtain a cutting face of 0.5 × 1 mm2 to 1 × 2 mm2, and ultrathin sections in the gold to mat the silver interference color range were cut with the Ultracut microtome and brought on pioloform-coated copper grids (200 mesh). The sections were stained with Reynolds lead citrate for examination with the Philips EM208S transmission electron microscope (FEI). 18
Evaluation of the severity of tracheal lesions
SEM allowed analysis of the effects of NDV vaccination on goblet cell activation and ciliated cells separately, and their respective earlier described degrees of activation and deciliation could readily be discriminated. 7 For each bird, lesions were classified by these criteria and scored accordingly as indicated in Table 1. Proximal, middle, and distal parts of the trachea were examined separately, but a combined, mean lesion score was retained because little variation between parts was observed. The severity index at each time point was calculated by dividing the sum of lesion scores by the total number of chickens subjected to evaluation.
Scoring system to quantify ultrastructural changes of the ciliated cells and activation of the goblet cells as observed by SEM.

Results
Humoral responses
Intranasal vaccination of the 1-day-old SPF chickens with 106 EID50 of the NDV La Sota strain resulted in the generation of NDV-specific humoral responses. Peak IgM responses were recorded at 10 days post-immunization (dpi), whereas an IgG response was observed from 15 dpi. Mean high HI titers of 7.3 and 6.3 were observed 10 and 15 dpi, respectively, as expected because Ig of both isotypes contribute to the inhibition of hemagglutination. In control chicks, no specific IgM, IgG, and HI levels were observed (not shown).
Electron microscopy
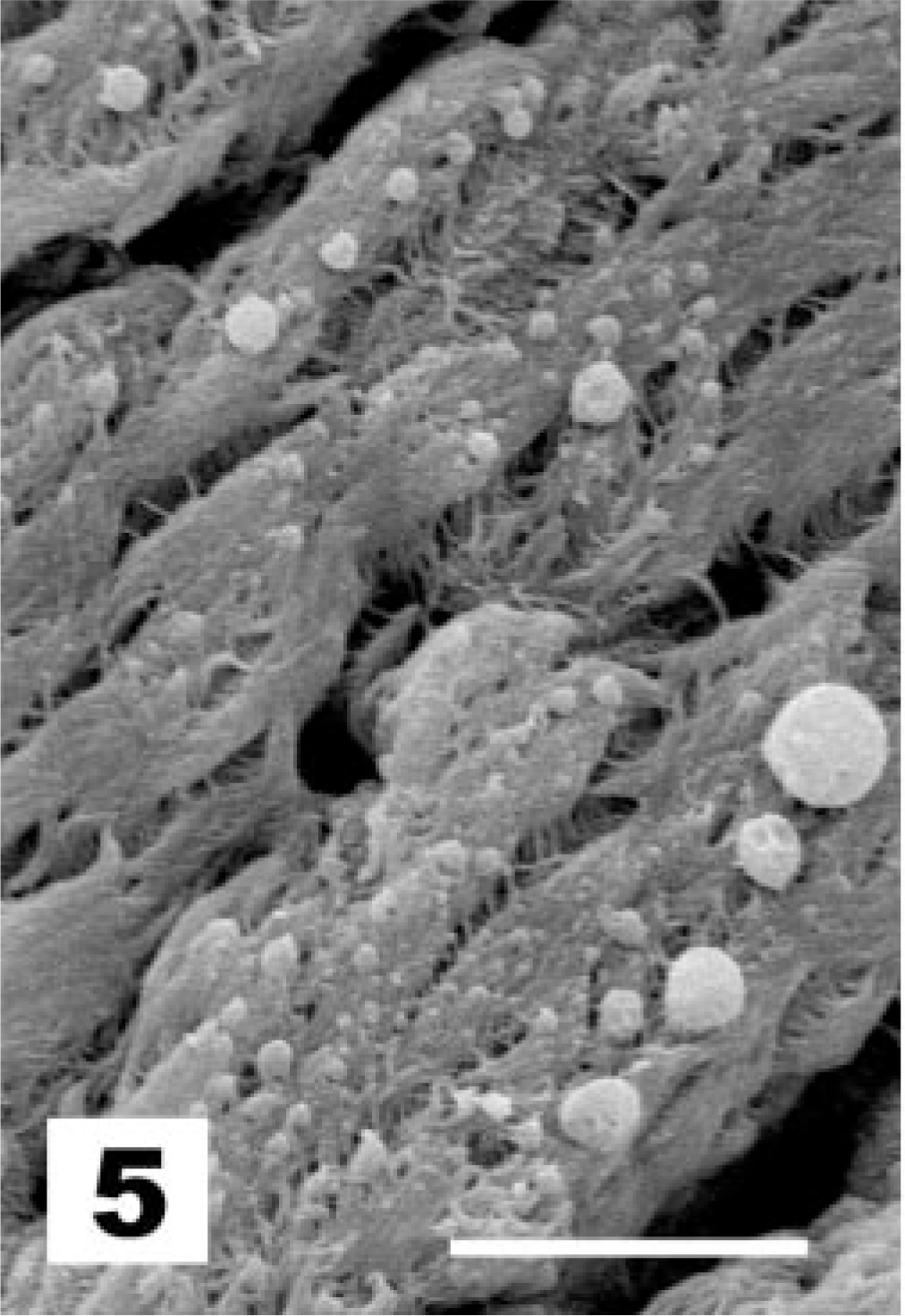
In sham-treated chicks and under normal conditions, as at the moment of vaccination and after recovery from infection, the tracheal surface of the chickens was lined by a pseudostratified columnar epithelium. Figure 1 demonstrates the presence of nuclei situated at different levels, suggesting that the epithelium has multiple layers of cells. However, all cells maintain contact with the basal membrane, although not all reach the free surface. In this epithelium, ciliated and goblet cells predominate. In normal conditions in which the goblet cells are not activated, the mucus layer that covers the tracheal epithelium is largely washed away during the preparative steps. The surface of the underlying epithelial cells is thus revealed for SEM examination. Figure 2 shows the luminal surface of the epithelium on which patches of nonciliated cells are surrounded by masses of ciliated epithelial cells. Both ciliated and goblet cells are covered by microvilli. The apical surface of goblet cells is visible, but the cells are not activated or swollen and do not or rarely expose their mucus content. This situation represents normal conditions and received the score of 1 (Table 1). Under these conditions, cell types other than goblet and ciliated cells, such as immature goblet cells and basal cells, could be readily observed by TEM. Basal cells stand out because their nuclei lie close to the basal lamina and because these cells do not extend to the luminal surface (Fig. 1). All chicks of the sham-treated control group received lesion score 1 at all examined time points.
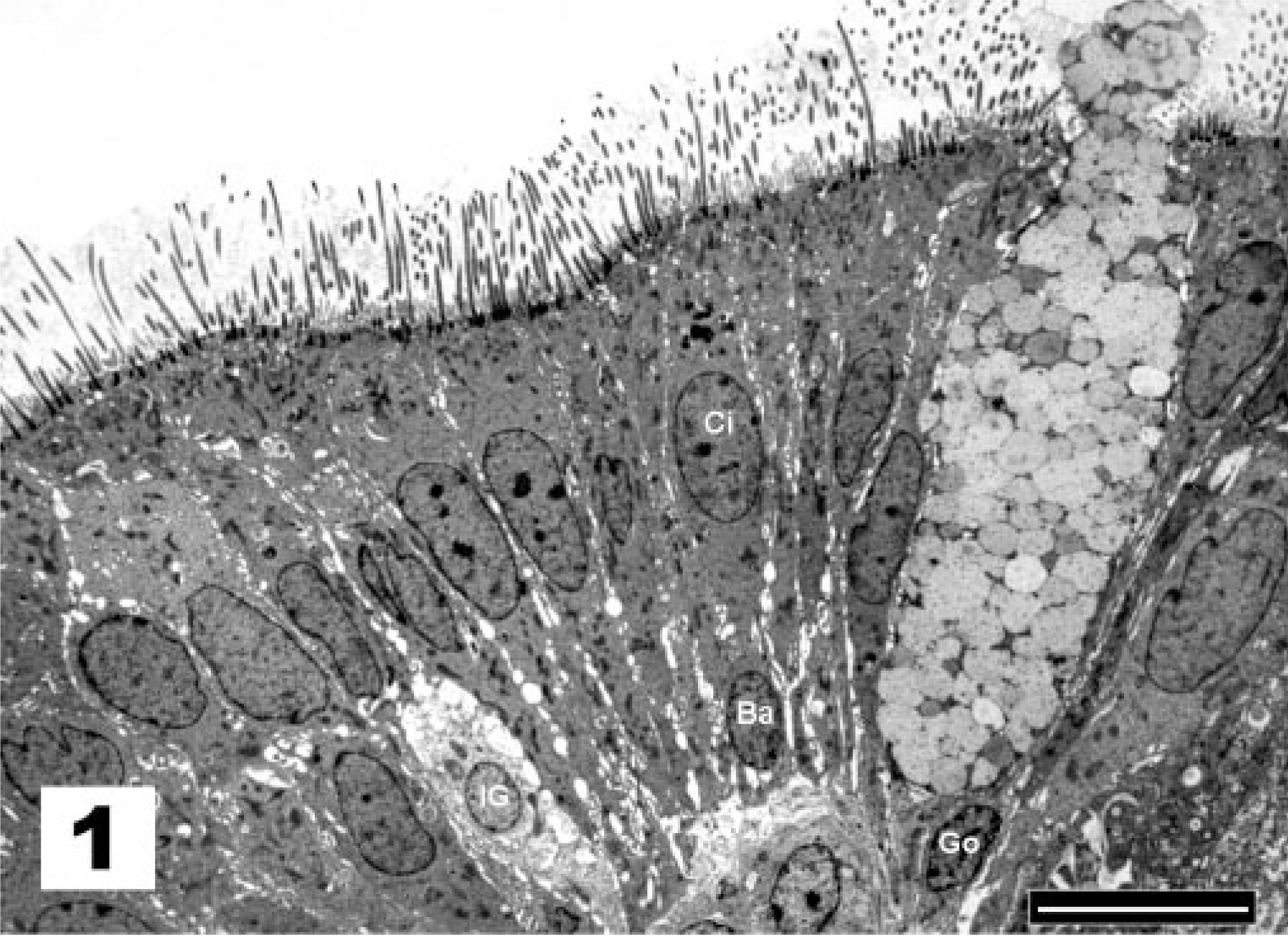
Tracheal epithelium; sham-inoculated chicken. The nuclei of ciliated cells (Ci) of the pseudostratified epithelium are situated at a more apical level than the nuclei of goblet cells (Go), basal cells (Ba), and immature goblet cells (IG), suggesting that the epithelium has multiple layers of cells. TEM, Reynolds lead staining. Bar = 10 μm.

Tracheal epithelium; sham-inoculated chicken. Patches of nonciliated cells are surrounded by masses of ciliated epithelial cells, and no or limited goblet cell activation is observed (score 1). SEM. Bar = 10 μm.
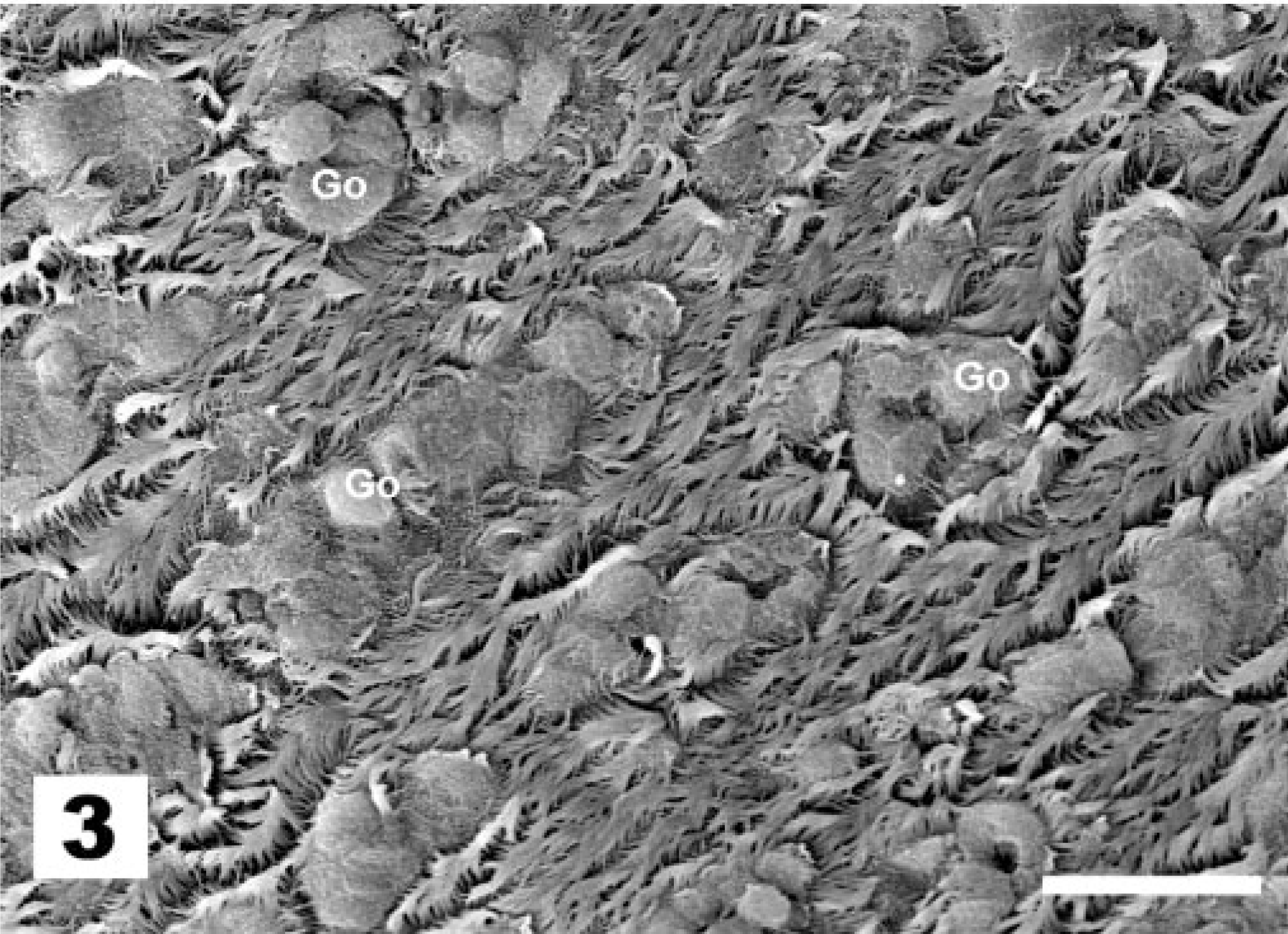
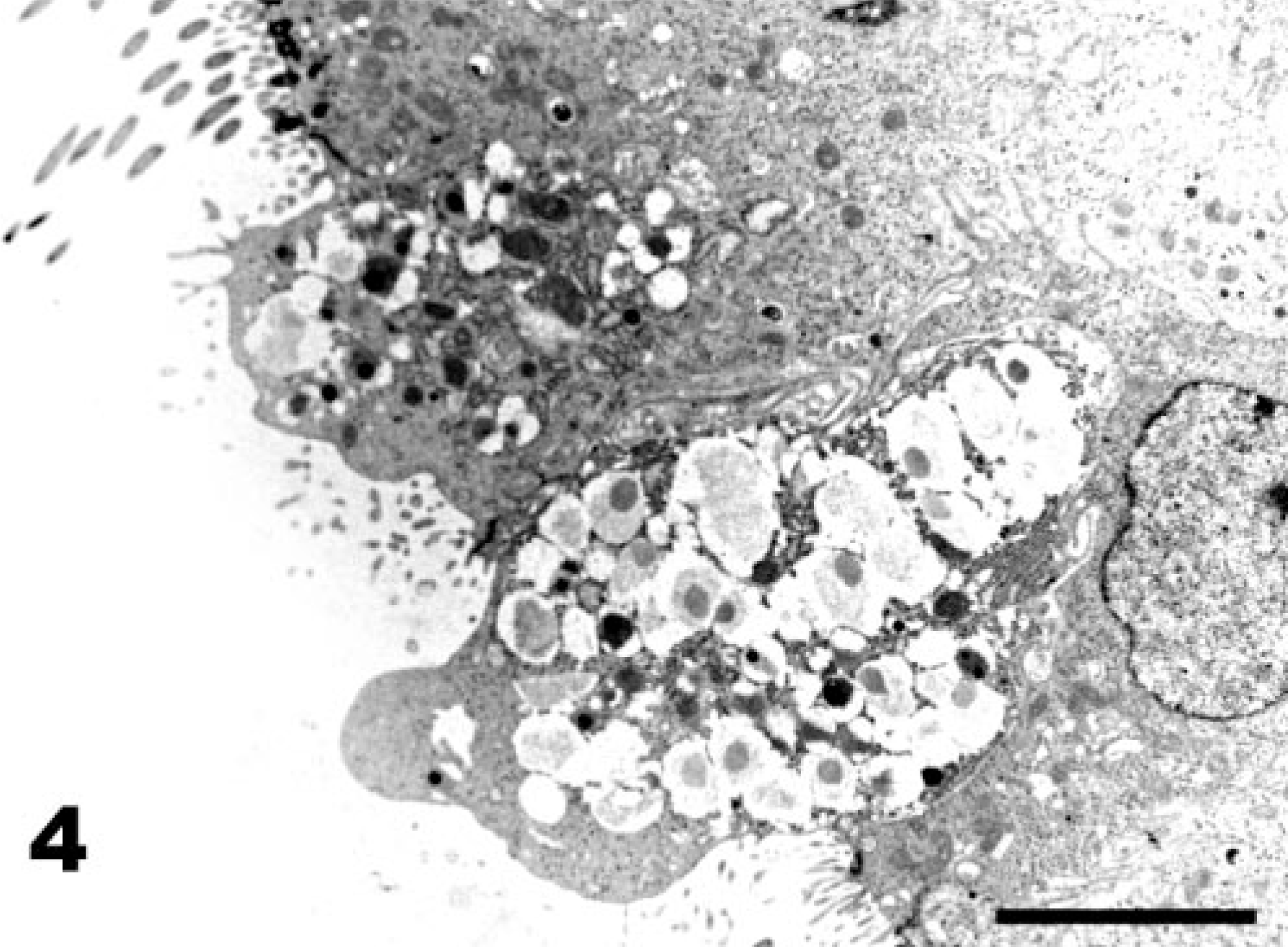
The first observed sign of ultrastructural change of the tracheal epithelium after La Sota vaccination is the hypertrophy of goblet cells, which exposes them at the epithelial surface (Fig. 3, score 2). Globular processes of mucus bulge into the lumen of the trachea and are shed, wrapped in Golgi-derived membranes (Fig. 4). Afterwards, goblet cells produce an excess of mucus, with globular mucus particles (Fig. 5) or blankets covering the epithelium (Fig. 6). The end result of goblet cell activation is the rupture of goblet cells when the openings of ruptured cells are exposed (Fig. 7). The conditions exemplified by Figs. 5–7 all receive lesion score 3. TEM analysis clearly illustrates the disruption of the hypertrophied goblet cells. Initially, secretion of mucus results in a gap between the globules of the mucus-filled cup of the goblet cell (Fig. 8). Afterwards, secretion continues, leaving the nuclei and residual cytoplasm of the ruptured goblet cell covered with mucus and bulging into the lumen of the trachea (not shown).
Tracheal epithelium; chicken 3 DPI with the La Sota NDV strain. Goblet cells are hypertrophied (some are marked Go) and bulge into the lumen of the trachea. SEM. Bar = 10 μm.
Tracheal epithelium; chicken 3 DPI with the La Sota NDV strain. Goblet cells are hypertrophied and their globular processes of mucus bulge into the lumen of the trachea. (score 2). TEM. Reynolds lead staining. Bar = 10 μm.
Tracheal epithelium; chicken 4 DPI with the La Sota NDV strain. Formation of globular mucus particles. SEM. Bar = 5 μm.
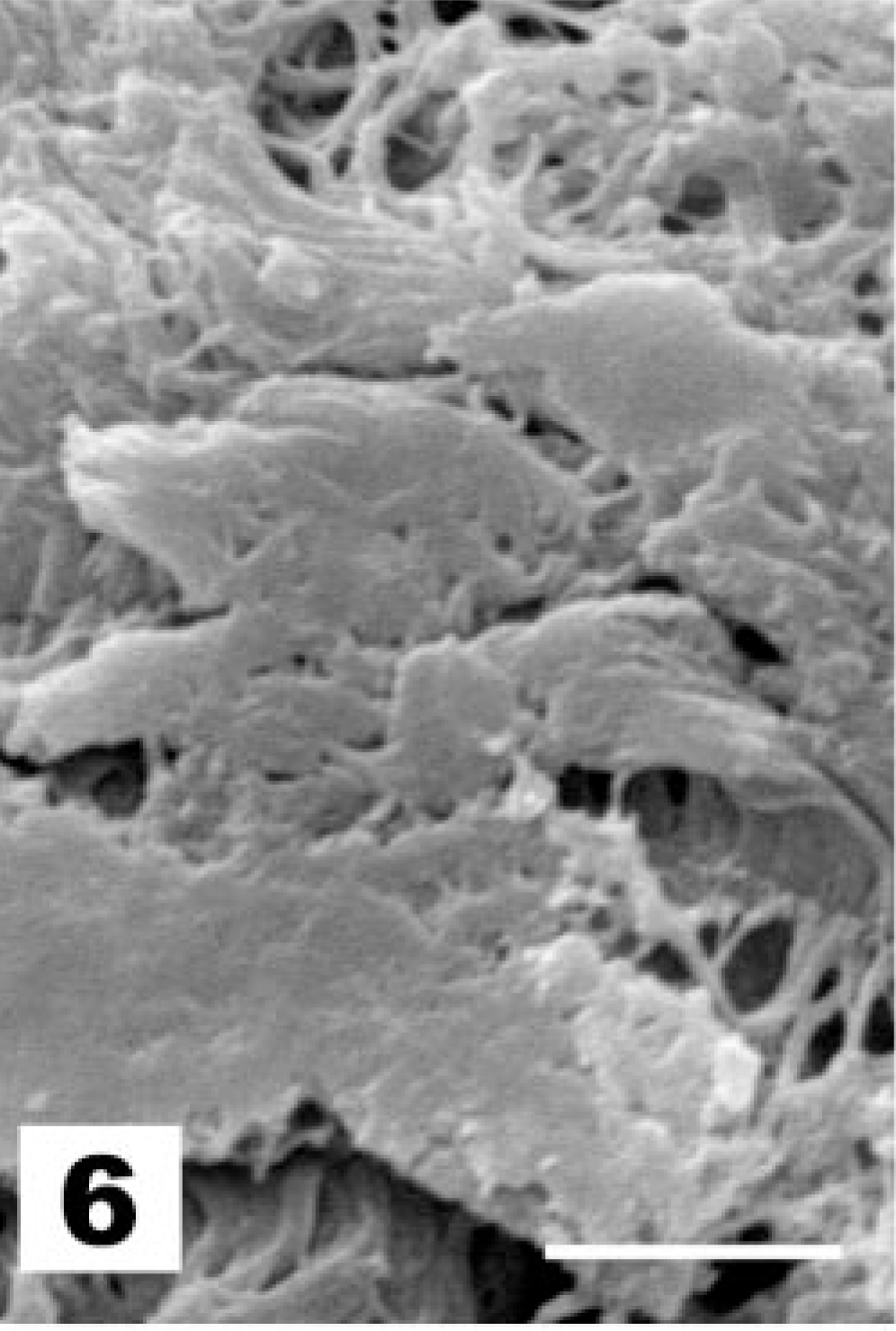
Tracheal epithelium; chicken 4 DPI with the La Sota NDV strain. Blankets of mucus covering the underlying epithelium. SEM. Bar = 5 μm.

Tracheal epithelium; chicken 4 DPI with the La Sota NDV strain. Rupture of epithelial goblet cells. SEM. Bar = 10 μm.
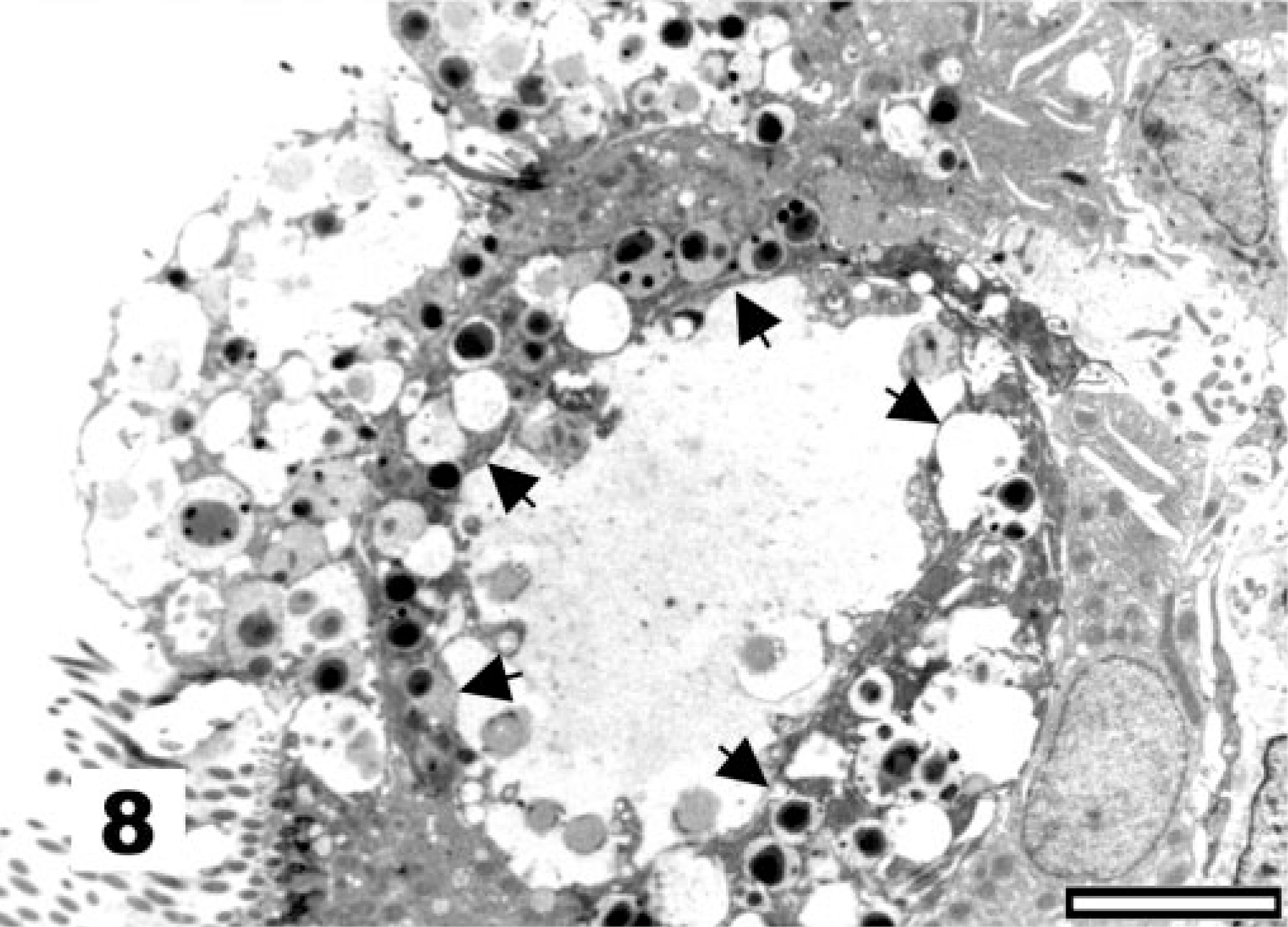
Tracheal epithelium; chicken 4 DPI with the La Sota NDV strain. Formation of a gap, demarcated by arrows, between the globules of the mucus-filled cup of activated goblet cells. TEM, Reynolds lead staining. Bar = 5 μm.
At the level of the ciliated cells, vaccination with the NDV La Sota strain results first in a marked increase in the proportion of nonciliated to ciliated cells (Fig. 9, score 2) and later in an almost complete deciliation (Fig. 10, score 3) of the tracheal surface. In these cases of deciliation, the original pseudostratified epithelium was replaced by a simple squamous (Fig. 11) to cuboidal epithelium (Fig. 12). In some instances, diapedesis of cells with lymphoid morphology was observed (Fig. 12).

Tracheal epithelium; chicken 7 DPI with the La Sota NDV strain. Marked increase of the proportion of non-ciliated to ciliated cells. SEM. Bar = 10 μm.
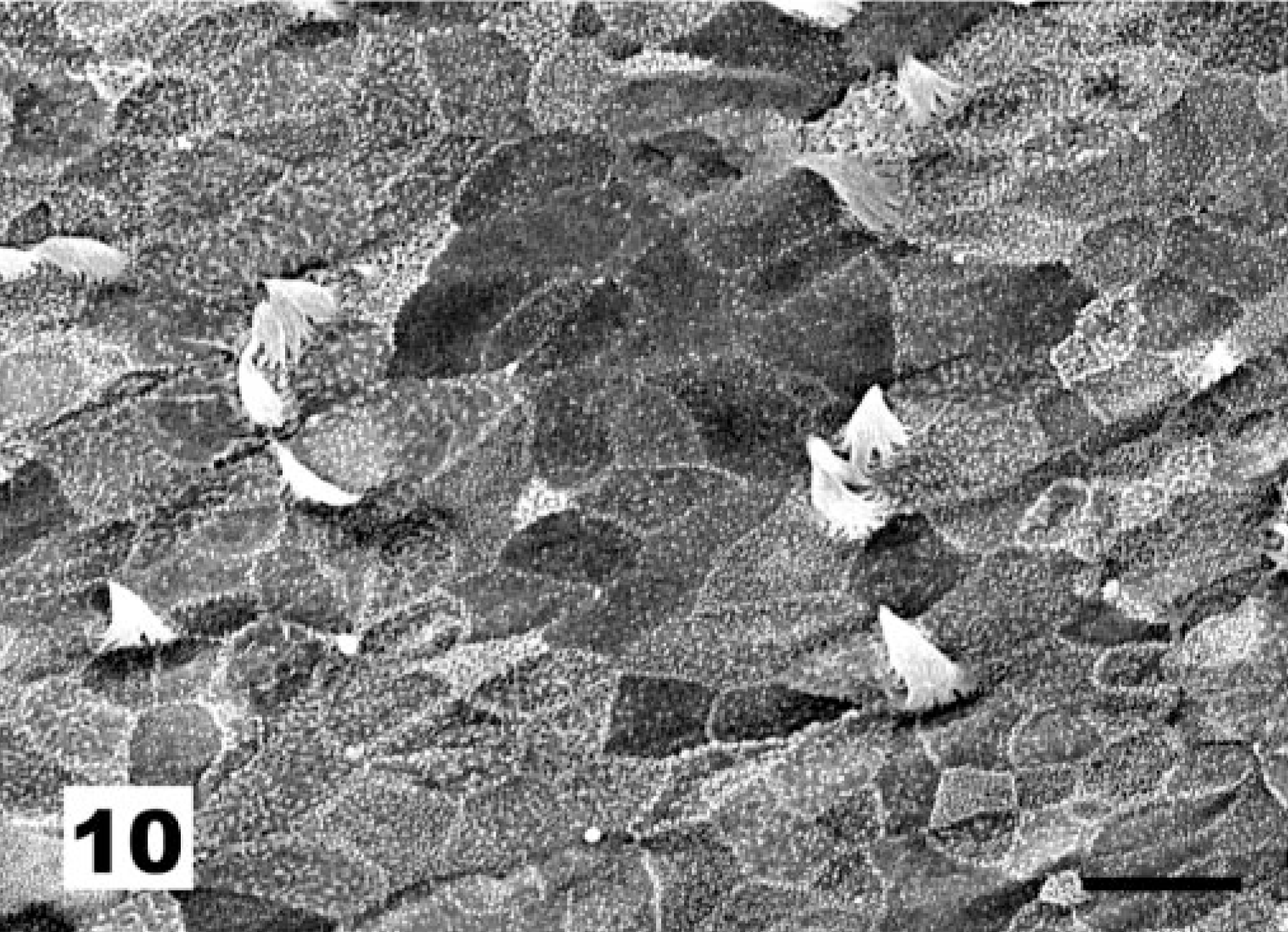
Tracheal epithelium; chicken 10 DPI with the La Sota NDV strain. Almost complete deciliation of the tracheal surface. SEM. Bar = 10 μm.
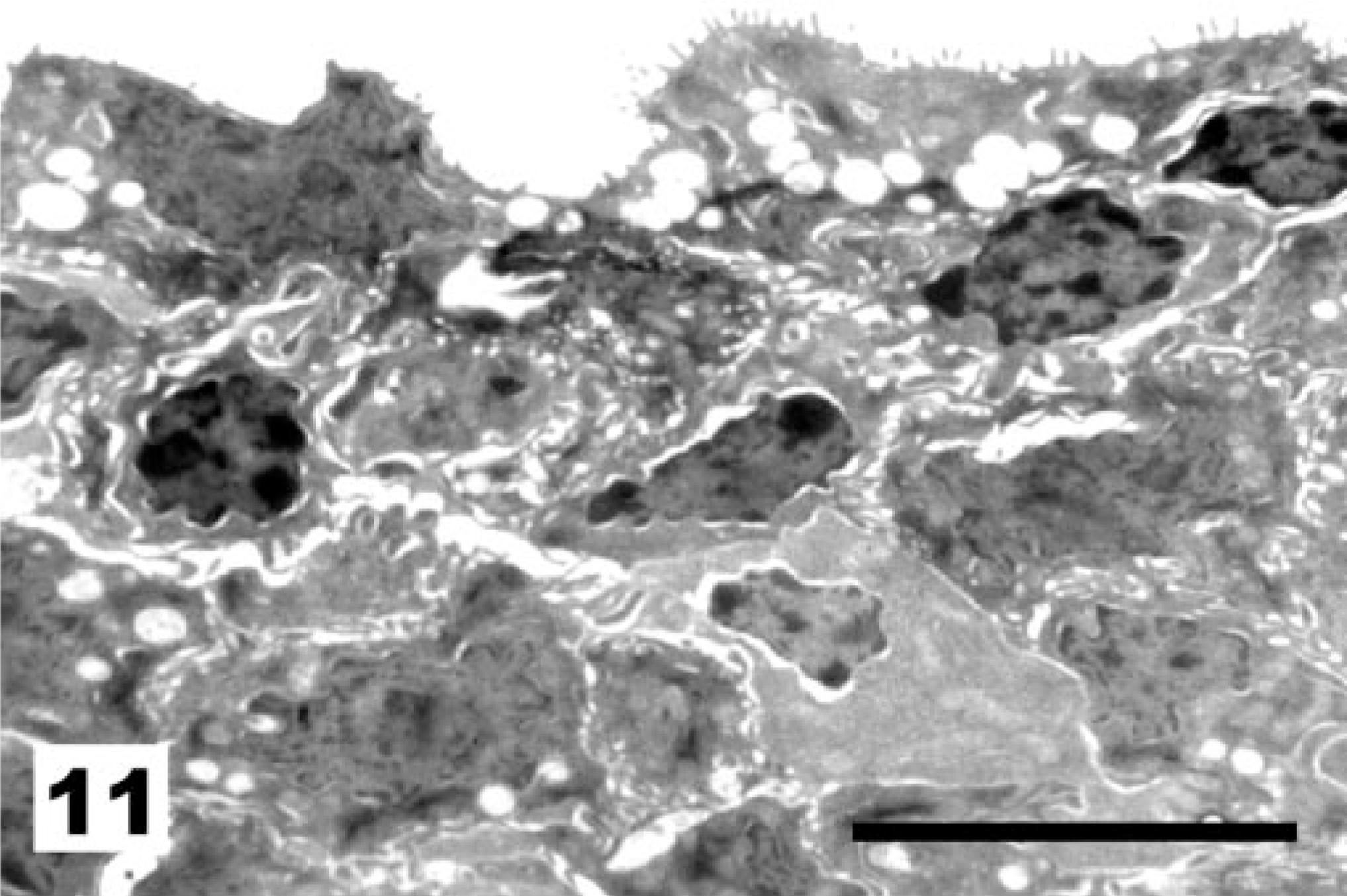
Tracheal epithelium; chicken 10 DPI with the La Sota NDV strain. Replacement of the original pseudostratified epithelium by a simple squamous epithelium. TEM, Reynolds lead staining. Bar = 10 μm.
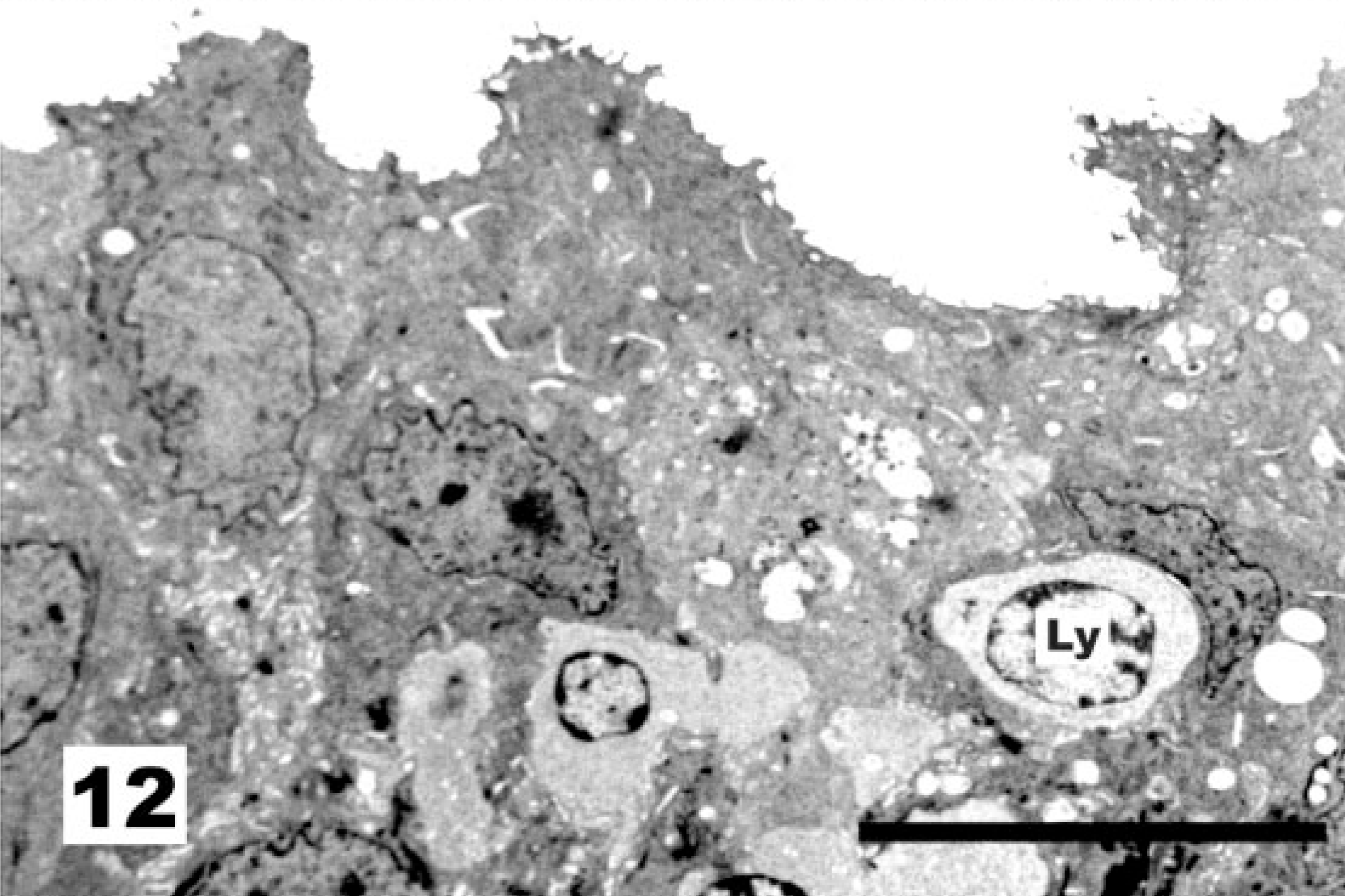
Tracheal epithelium; chicken 10 DPI with the La Sota NDV strain. Replacement of the original pseudostratified epithelium by a simple squamous to cuboidal epithelium. Diapedesis of cells with lymphoid morphology (nucleus marked Ly) was observed. TEM, Reynolds lead staining. Bar = 10 μm.
Following the above-described scoring system, as summarized in Table 1, the ultrastructural changes observed by SEM were scored and averaged by age. A peak activation of the goblet cells of all birds (lesions score 3) vaccinated at 1 day of age with 106 EID50 of the La Sota NDV strain was observed within 4 days postvaccination. Afterwards, lower goblet cell activation levels with mean lesion scores of 2.2, 2.3, 1.5, and 2 were observed. The peak of goblet cell activation is followed by a NDV-induced deciliation of the tracheal epithelium, when a simple squamous to cuboidal epithelium replaces the original pseudostratified epithelium. One of three chicks shows a denuded epithelium on day 5 already, whereas all 7- and 10-day-old chicks demonstrated deciliation ranging from a marked decrease of ciliated cells to a complete deciliation and desquamation of the epithelium (mean lesion scores of 2.8 and 2.5, respectively).
At 15 days of age, all epithelial damage and activation was restored. The proportion of ciliated versus nonciliated cells appears somewhat higher than in newly hatched chickens (not shown). Many of the goblet cells are, however, not visible from the luminal side of the trachea as demonstrated by TEM (Fig. 13).
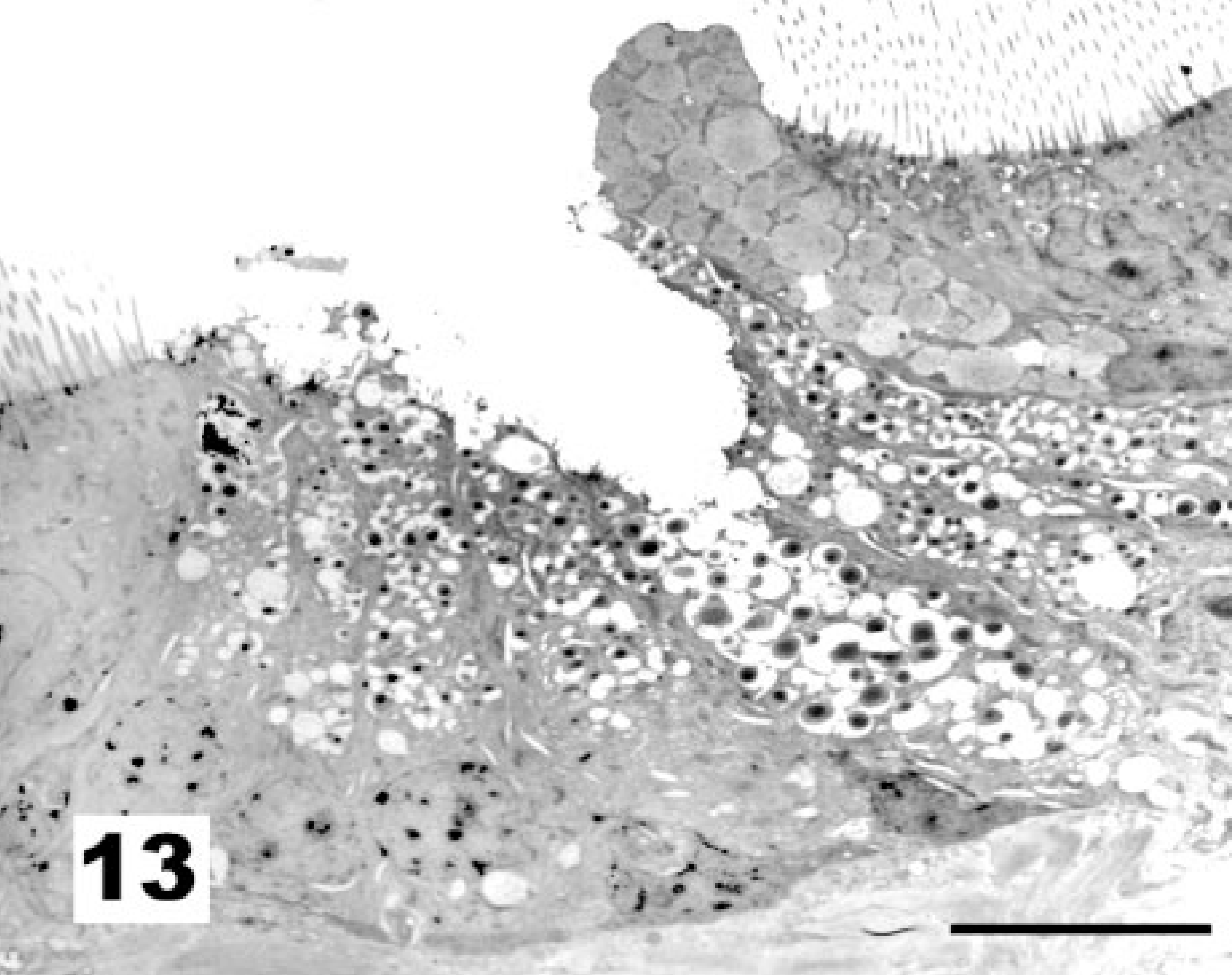
Tracheal epithelium; chicken 15 DPI recovered from vaccination with the La Sota NDV strain. Compared with unvaccinated chicks, goblet cells tend to group in epithelial folds. TEM, Reynolds lead staining. Bar = 10 μm.
Discussion
SEM analysis confirmed that vaccination of 1-day-old SPF chickens with live virus of the lentogenic La Sota NDV strain results in damage to their tracheal epithelium, as observed earlier in chickens 7, 8, 17 and in embryos and tracheal explants. 9 In our hands, TEM allowed the visualization and interpretation of the effects of NDV on goblet cells and ciliated cells.
An early strong activation of the goblet cells was observed in all vaccinated chicks. This most likely reflects the initial NDV infection of these cells. The subsequent decrease of the goblet cell activation score can be interpreted as the result of goblet cell exhaustion and virus replication eliminating goblet cells because severe ultrastructural decay and rupture of goblet cells was observed after the initial activation.
The increase in the lesion score of ciliated cells between 5 and 10 days postvaccination reflects the replacement of the highly differentiated, pseudostratified epithelium containing ciliated cells by a simple squamous to cuboidal epithelium without cilia. This replacement should be interpreted as the regeneration phase during which the basal cells at the periphery of a lesion slide across the underlying fibroblastic connective tissue to close the lesion. 21 It makes sense to consider that basal cells survive infection with lentogenous NDV strains on the basis of findings that the Sendai virus, a rodent paramyxovirus, infects ciliated cells and secretory cells but not basal cells of the rat tracheal epithelium. 13
As expected, proliferation of the La Sota virus induced a typical primary humoral response, with peak IgM responses preceding the rise of IgG. The rise of the titers of HI antibodies, shown to be necessary in providing protection from NDV, 19 coincided with restoration of the mature airway epithelium by further differentiation, including ciliogenesis and formation of secretory cells. Recovery was complete within 15 days postvaccination when high NDV-specific IgG titers were observed. Kotani et al. 7 observed this correlation between histopathologic changes in affected trachea and levels of antibody too: viral antigen was observed in the regressive phase of the infection, and HI antibodies were observed in the regenerative phase. Furthermore, comparable transient tracheal damage was observed with the low-virulence Ishii 7 and B1 strains of NDV, 17 whereas a highly virulent Malaysian strain caused a complete exfoliation of the tracheal epithelium and death of the infected chicks. 8 In agreement, immunohistochemistry demonstrated antigen of the Roarkin NDV strain in the epithelial cells of the trachea 5 days, but not 2 and 7 days, after vaccination. 10
The vaccination of day-old chicks with live virus of the La Sota strain of NDV induced a uniform, strong humoral immune response. This, and the absence of responses in sham-treated birds, indicates a uniform NDV infection of all birds, which is expected for intranasal vaccination. On one hand, comparable kinetics and intensity of responses are desirable in field conditions in which La Sota and its derived clones are widely used for vaccination of poultry. On the other hand, considering the residual pathogenicity of the La Sota vaccine, one can assume that similar concomitant subclinical lesions to the trachea that were brought about here experimentally might be induced in commercial poultry exploitations as well. However, it should be stressed that this vaccine is not used for day-old chicks in the field and that most chicks possess maternally derived antibodies that might partially interfere with vaccination in the field. A further similar study should be performed in 2-week-old SPF chickens. Nevertheless, even at a later age, NDV vaccination is a well-known factor for secondary infections like colibacillosis, which could be substantiated in experimental conditions. 1, 5, 17 The observed NDV-induced replacement of the mature pseudostratified epithelium by a nondifferentiated, simple squamous to cuboidal epithelium explains this at the ultrastructural level: because of its lack of mature goblet cells and cilia, the cuboidal epithelium cannot trap particles, including bacteria, in mucus and clear the mucus-associated particles by ciliary movement. Therefore, our results reinforce the recommendation that La Sota vaccine should not be used in young unprimed chickens.
Footnotes
Acknowledgements
This research was granted by the Belgian Ministry of Agriculture and Middle Classes (Project DG6-S6037, section 1). We thank Ms. Nadine Dubois and Ms. Marina Ledecq for their expert technical assistance.