
Abstract
Three marine viral hemorrhagic septicemia virus isolates were used to bath challenge turbot with the purpose of studying mortality and the pathology and antigen distribution over time. Two high-virulence isolates, 860/94, 4p168 and a low-virulence isolate 1p3 from a Baltic Sea herring were used. Organ samples were collected sequentially at 2, 4, 7, 10, 15, 20, 25, and 45 days postinfection. Specimens were processed for virology, histopathology, and immunohistochemistry. Organs during the early stages of infection (from 2 to 7 days) had virus isolation from all three groups only on day 7. Virus titer in kidney and heart sampled at day 25 was higher for the two virulent isolates compared with the low-virulence isolate. The viral distribution in situ of the two more virulent isolates from turbot (860/94) and herring (4p168) resembled viral hemorrhagic septicemia in rainbow trout with regard to the target organs. Early infection of endothelial cells in both kidney and heart was observed. Accumulated mean mortality was 41.5% for the turbot isolate 860/94, 48% for the herring isolate 4p168, and 3.5% for the herring isolate 1p3. This study revealed that the isolates from turbot (860/94) and herring (4p168) induced significantly higher mortality compared with the virus-free control and the herring isolate (1p3). The onset of mortality is markedly later in turbot compared with what is seen in rainbow trout.
Keywords
The first detection of viral hemorrhagic septicemia virus (VHSV) in marine fish was from ulcer lesions in cod. 10 VHSV has since been identified from several other fish species from the northern European marine environment, such as cod Gadus morhua; 10 , 12 , 15 , 18 , 23 , 24 herring Clupea harengus; 5 , 15 , 18 haddock Melanogrammus aeglefinus; 24 sprat Sprattus sprattus, blue whiting Micromesistius poutassou, fourbearded rockling Rhinonemus cimbrius, lesser argentine Argentina sphyraena; 18 Norway pout Trisopterus esmarkii, whiting Merlangius merlangus; 15 , 18 poor cod Trisopterus minutus; 15 and common dab Limanda limanda, flounder Platichthys flesus and plaice Pleuronectes platessa (H. Mortensen, unpublished).
Outbreaks of VHS have occurred in farmed turbot (Scophthalmus maximus) in Germany, 22 Scotland, 19 , 21 and Ireland (J. McArdle, unpublished). Studies by Snow 26 and King 14 demonstrated 60% mortality by immersion challenge of juvenile turbot with the Scottish isolate from a farm outbreak at Gigha, Scotland.
Investigations of the pathology of marine VHSV infection in marine fish species have been limited. Histologic sections from farmed turbot in an outbreak in Scotland showed that diseased fish had widespread necrosis in cardiac muscle and hepatocytes and hemorrhage behind the retina and within the muscle. 21 Viral hemorrhagic septicemia (VHS) pathogenesis is more thoroughly described in rainbow trout. 3 , 7 VHS is a hemorrhagic disease with the hematopoietic tissue of the kidney and spleen as the main target. Macrophages are subject to infection, and in a more advanced stage, melanomacrophage are destroyed. 7 Leukopenia is evident, as is vascular and sinusoidal congestion. 28 Although pathology after infection of turbot with marine VHSV has been documented under field conditions in Germany 22 and Scotland, 19 , 21 no experimental sequential pathology studies have been published to date.
The aim of this study was to describe the sequential development of pathologic changes in experimental infections with marine VHSV isolates in turbot using isolates of known high and low virulence.
Materials and Methods
Fish
Turbot (mean weight 10 g) were virologically tested in cell culture (12 pools of 5 fish) and were found negative for VHSV, infectious pancreatic necrosis virus, and infectious hematopoietic necrosis virus (IHNV). Fish were starved for 24 hours before infection and put into a 30-liter aquarium lined with polythene bags and containing 10 liters of sea-water (sand filtered from Nigg Bay, Aberdeen) with aeration. Duplicate tanks each holding 30 fish (mortality study) or 50 fish (pathogenesis study) were set up for each test group.
VHSV propagation
Stocks of virus were grown on the bluegill fry caudal trunk cell line (BF-2, pass number 3–6) cultivated in Glas-gow minimum essential medium (GMEM; Gibco, Invitrogen, Groningen, The Netherlands), plus 10% fetal bovine serum (FBS, Sigma, St Louis, MO) (GMEM-10). Each virus was diluted in GMEM-10 to give a dose of 109 TCID50 per isolate to be added to each tank, hence, giving a final dose of 105 TCID50/ml per isolate in a volume of 10 liters. Negative control groups received GMEM-10 alone.
Infection protocol
The virus solutions were poured into duplicate tanks for each group and thoroughly mixed. The polythene bags were tied to contain aerosols and reduce the risk of cross-contamination between tanks. After a 6-hour exposure to the virus, the fish were gently tipped out of the bags into the tanks. The water volume was raised to 30 liters, and water flow rates were set to 50 liters per hour. The water temperature was maintained at 11 C ± 1 C, and aeration was provided throughout the duration of the experiment. The photoperiod was set for 12 hours of daylight, and the fish were fed a commercial pellet diet to satiation. Wastewater was disinfected according to standard protocol.
Mortality study
Group 1 fish were challenged with isolate 860/94, from an outbreak on a turbot farm in Scotland. 19 , 21 Group 2 was exposed to isolate 4p168, sampled from herring in the Skagerrak. Group 3 was exposed to isolate 1p3, sampled from herring in the Baltic Sea. The Danish Veterinary Laboratory 18 kindly supplied isolates 4p168 and 1p3, and the virulence of these isolates for turbot has previously been determined by King. 14 Group 4 was a negative control. Fish were checked daily, and dead fish were removed and stored at − 80 C.
Pathogenesis study
After infection tissue sampling was performed as described below.
Tissue sampling
In groups 1, 2, 3, and 4, organ samples from four fish from each group were randomly collected sequentially at days 0 (before onset of experiment), 2, 4, 7, 10, 15, 20, 25, and 45 postinfection (p.i.). The fish were sampled and submerged whole in 10% phosphate-buffered formalin (pH 7.4). Tissue specimens from individual fish of skin, gills, brain, liver, spleen, kidney, heart, and intestine were dissected from fixed whole fish. The samples were embedded in paraffin wax according to standard procedures. 17
Immunohistochemical identification of VHSV
The sections were processed according to the method described by Evensen and Olesen. 8 After deparaffinization, the sections were incubated with 5% (wt/vol) bovine serum albumin in Tris-buffered saline (TBS, pH 7.4) for 20 minutes. The solution was removed from the slides, and a monoclonal antibody specific for nucleoprotein from VHSV (IP5B11) was applied to the slides at a dilution of 1 : 700 for 1 hour at 37 C. After washing for 5 minutes in TBS, the secondary antibody, biotinylated anti-mouse immunoglobulin diluted 1 : 300 was added. The solution and tissue were incubated for 30 minutes at room temperature. The sections were washed for 5 minutes in TBS before incubation with avidin–biotin alkaline phosphatase diluted 1 : 500 for 1 hour at 37 C. After washing, Fast Red (1 g/liter, Sigma) and Naphthol AS-MX-phosphate (0.2 g/liter, Sigma) with 1 mM levamisole (Sigma) in 0.1 M TBS were added to develop for 20 minutes. Sections were washed in tap water before being counterstained with Mayer's hematoxylin and mounted in aqueous mounting medium (Aquamount, BDH Laboratory, Dorset, UK).
VHSV isolation by cell culture
In groups 1, 2, 3, and 4, six turbot from each group were collected at days 0 (before onset of experiment), 2, 4, and 7 p.i. and frozen whole at − 80 C. Tissue specimens from skin, gills, intestine, kidney, and heart were sampled after thawing. Samples were diluted 1/10 in Eagle's minimum essential medium with Earle's balanced salt solution supplemented with 16.4 mM Tris buffer, 5.3 mM NaHCO3, 10% FBS, 4 mM L-glutamine, and 50 µg/ml gentamicin. While immersed in an ice solution, the pellet was homogenized by a pellet mixer (kidney and heart) or a homogenizer (skin, gills, and intestine). The homogenate was centrifuged (1,500 × g, 4 C, 15 minutes, Beckmann GS-6R). The supernatant was added to BF-2 cells cultured on 96-well plates (20–48 hours old). The final dilution of tissue material in cell culture medium was 1 : 100. The inoculated cells were incubated at 15 C and were inspected for up to 7 days for the occurrence of cytopathic effect.
Samples of posterior kidney and heart from six fish from each group at day 25 p.i. were subjected to virologic examination. Titration of virus was performed in 96-well plates with Bluegill sunfish cells (BF-2, ATCC CCl 91) TCID50 standard procedures for quantification of virus. 13 Virus collected from the cell culture supernatant was identified according to standard procedures by the indirect fluorescent antibody technique 11 using monoclonal antibodies IP5B11 VHSV N protein (reference strain F1)
Statistical analyses
Identification of virus
The statistical analyses were performed in the SAS-PC System® Version 8 for Windows (SAS Institute Inc., Cary, NC). The descriptive analyses were conducted using PROC FREQ. The odds ratios (OR) calculated from the estimated coefficients in the final models were used to express the chance of getting a positive sample. The three experiments with repeated measurements over time conducted in the study were analyzed separately with two regression models for both. The repeated effect in all models was the variable, day of sampling. The independent variables, that is, type of infection (1 = 860/94, 2 = 4p168, 3 = 1p3) and the organs that were investigated in each fish, were included as fixed effects. The best model fit was found by forcing the variable type of infection into the model and by including the organs investigated.
The outcome variables in the experiment were the results from the immunohistochemical and cell culture identification of VHSV in the specimens (1 = positive, 0 = negative). Two repeated measurements regression models using PROC GENMOD were assessed, 2 , 27 and the tissue specimen was the statistical unit in both. The type III chi-square value was used to test the significance (P < 0.05) of the variables. The interaction terms between the two independent variables were also tested for significance.
One important assumption for traditional statistical methods is that the observations are independent of each other, which is assumed not to be the case in this study. In a repeated measurements analysis it is assumed that the observations within each cluster (fish in this case) over time (sampling day) are correlated. It is also possible to specify the structure of the correlations among the observations within each cluster. 2 In this study, we assumed that the best-fitting approach was a first-order autoregressive model in which one measurement is correlated with the previous one.
Mortality
Mortality incidence was subjected to Fisher's exact test (two-tailed), with a significance level of P = 0.01. The analysis was done relative to virus-free control 4A. Mortality was recorded for 45 days p.i.
Results
Sequential pathology study
Group 1—turbot (860/94) isolate
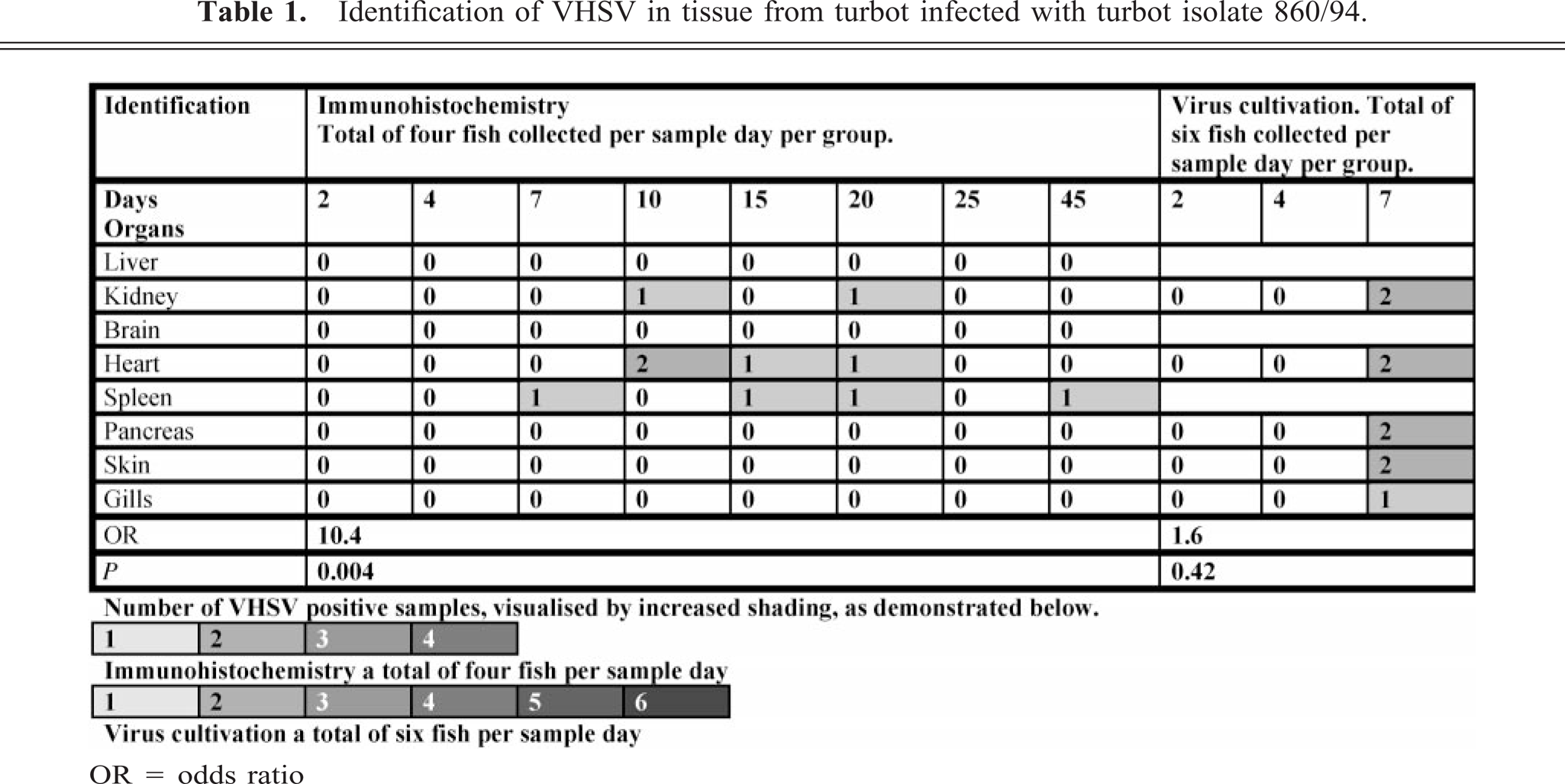
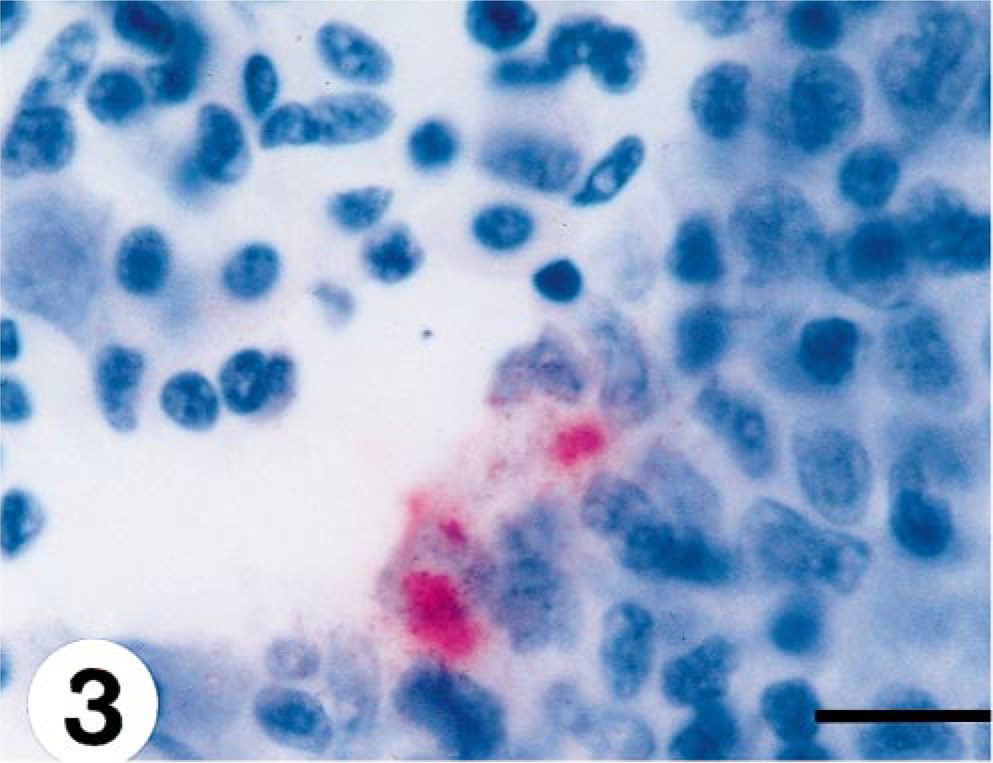
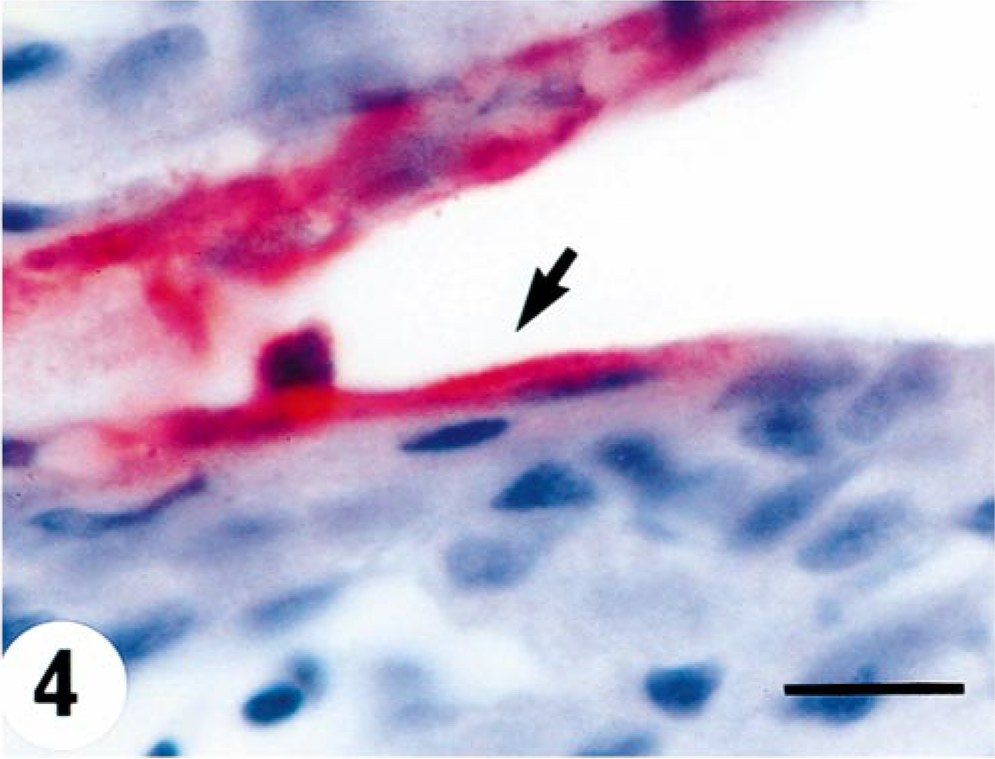
Virus was first detected by immunohistochemistry in the spleen of one fish at 7 days p.i. (Table 1). At days 15 and 20 p.i., one fish exhibited a positive reaction in heart and spleen and also in kidney at day 20 (Fig. 3). Viral antigen was identified in single cells in the white pulp of the spleen without any pathology. At day 10 p.i., there was positive virus staining in the endothelium and myocardium of the heart of two fish (Fig. 4). The pathologic changes vary from single cells being found positive for VHSV in kidney with either no degenerative change or punctate necrotic foci (Fig. 5). All fish were negative at day 25 p.i., and antigen was detected in the spleen of one fish at day 45 p.i with no associated pathologic change.
Identification of VHSV in tissue from turbot infected with turbot isolate 860/94.
Number of VHSV positive samples, visualised by increased shading, as demonstrated below.
1 2 3 4
Immunohistochemistry a total of four fish per sample day
1 2 3 4
Virus cultivation a total of six fish per sample day
OR = odds ratio
Kidney; turbot (Scophthalmus maximus), infected with 860/94 (high-virulence turbot isolate) at 20 days p.i. Single cells with demonstrable antigen are noted in the interstitium. Virus antigen is detected by alkaline phosphatase–coupled antibodies and observed by Fast Red. The slide is counterstained with Mayer's hematoxylin. Bar = 10 µm.
Heart; turbot (Scophthalmus maximus), infected with 860/94 at 10 days p.i. Virus-positive endothelial cells (arrow) lining myocardium of the ventricle. Virus antigen is detected by alkaline phosphatase–coupled antibodies and observed by Fast Red. The slide is counterstained with Mayer's hematoxylin. Bar = 10 µm.
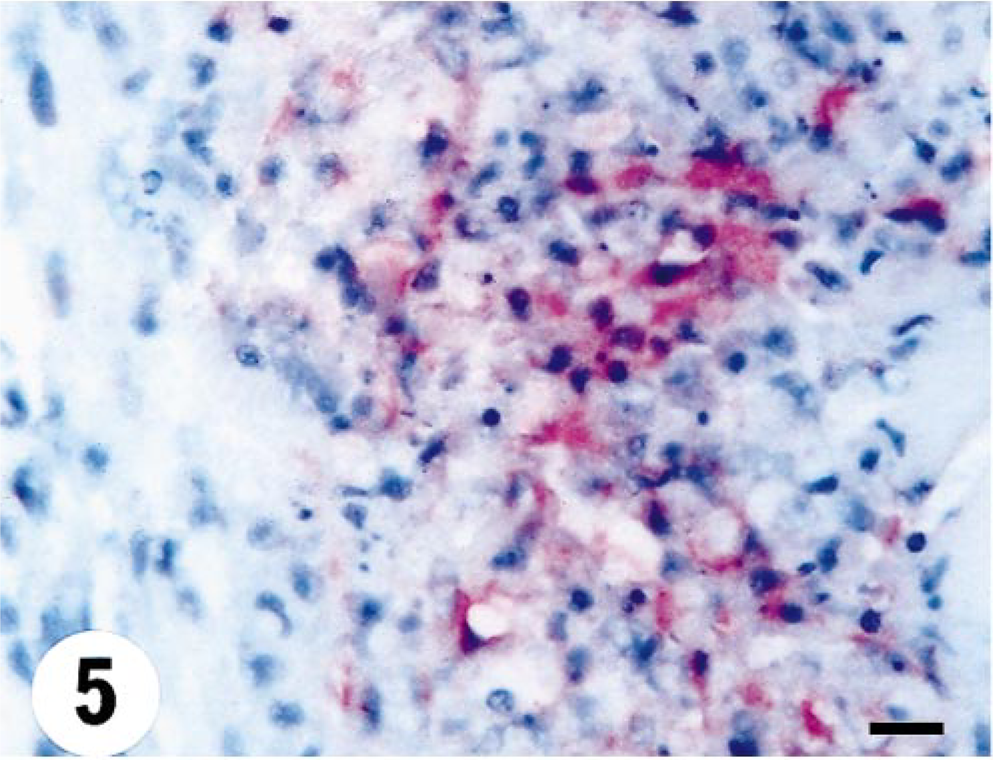
Heart; turbot (Scophthalmus maximus), infected with 860/94 at 15 days p.i. Single cells with demonstrable antigen are noted within the interstitium Focal area with positive staining for VHSV in the myocardium. Note leukocyte infiltration (lymphocytic) and area with degeneration and cell necrosis. Virus antigen is detected by alkaline phosphatase–coupled antibodies and observed by Fast Red. The slide is counterstained with Mayer's hematoxylin. Bar = 10 µm.
Group 2—herring (4p168) isolate
Positive reaction for VHSV was detected in three of four fish by immunohistochemistry at day 7 p.i. in heart, kidney, spleen, and pancreas (Table 2). In the early stages of the infection (up to 7 days), virus was observed in single cells in the spleen with no concomitant pathologic changes (Fig. 6). The myocardium had dense viral staining diffusely through the cytoplasm, frequently with insignificant pathologic changes (Fig. 7). In kidney, focal necrosis in the interstitium was observed (Fig. 8), and virus was detected in the endothelial cells (Fig. 9). At day 10 p.i., the tissue of all sampled fish were positive by immunohistochemistry. Three fish were positive both in heart and kidney and had detectable antigen in the white pulp.
Identification of VHSV in tissue from turbot infected with herring isolate 4p168.
Numher of VHSV positive samples, visualised by increased shading, as demonstrated below.
1 2 3 4
Immunohistochemistry a total of four fish per sample day
1 2 3 4 5 6
Virus cultivation a total of six fish per sample day
OR = odds ratio
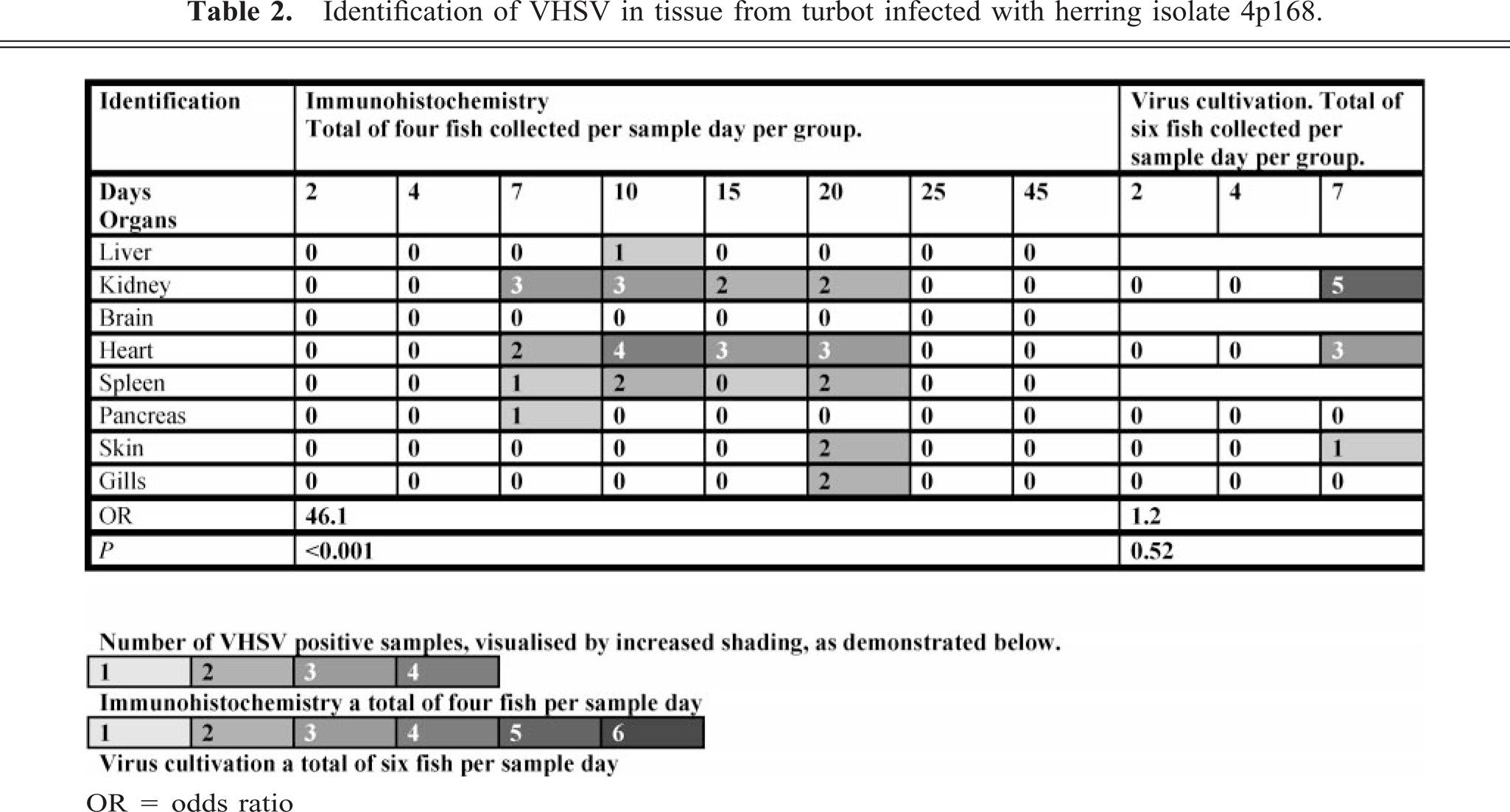
Spleen; turbot (Scophthalmus maximus), infected with 4p168 (high-virulence herring isolate) at 7 days p.i. Positive cells in the white pulp of the spleen, also associated with ellipsoids (arrows), without evidence of inflammatory or degenerative changes. Virus antigen is detected by alkaline phosphatase–coupled antibodies and observed by Fast Red. The slide is counterstained with Mayer's hematoxylin. Bar = 10 µm.
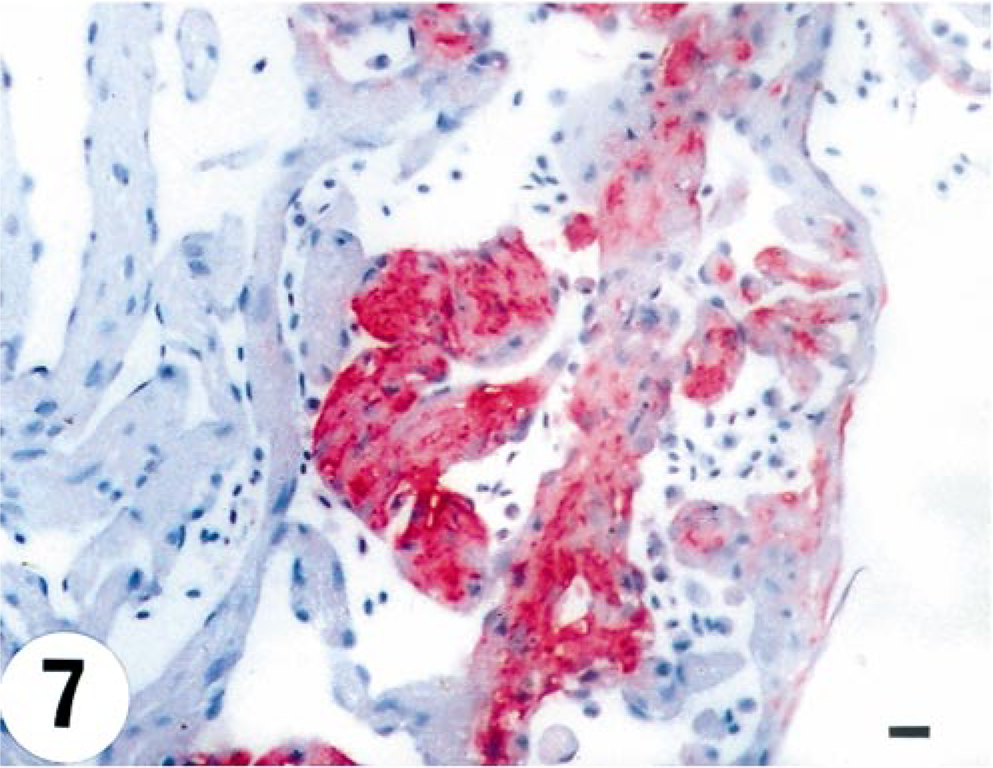
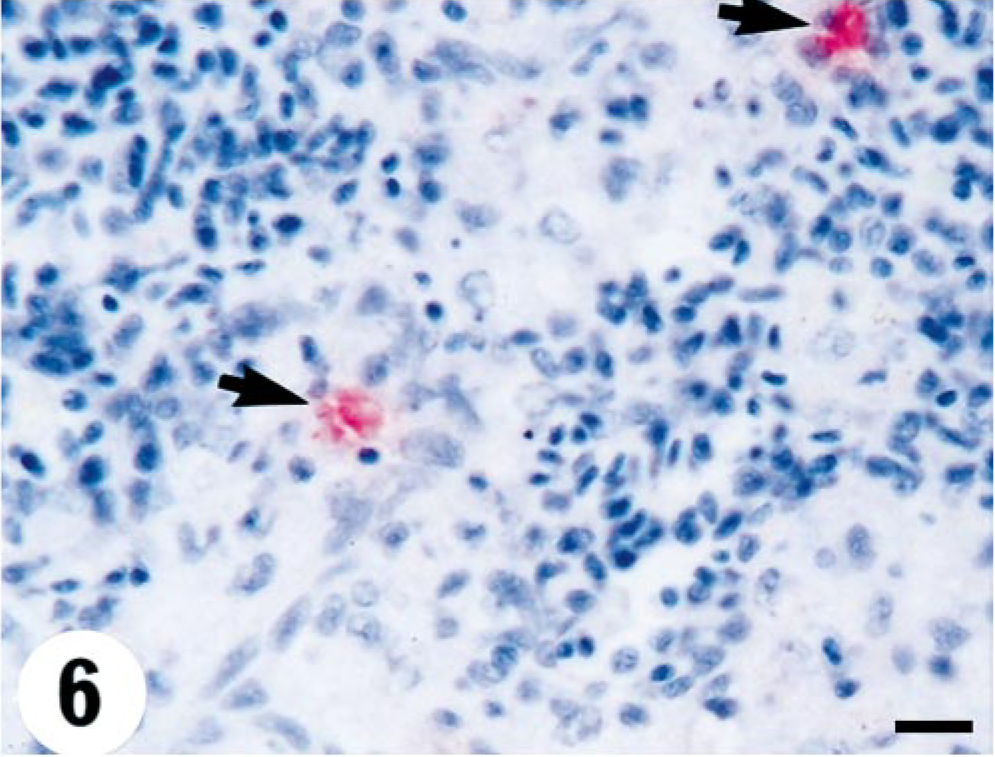
Heart; turbot (Scophthalmus maximus), infected with 4p168 at 7 days p.i. Intense staining for VHSV in the myocardium of the ventricle. Virus antigen is detected by alkaline phosphatase–coupled antibodies and observed by Fast Red. The slide is counterstained with Mayer's hematoxylin. Bar = 10 µm.
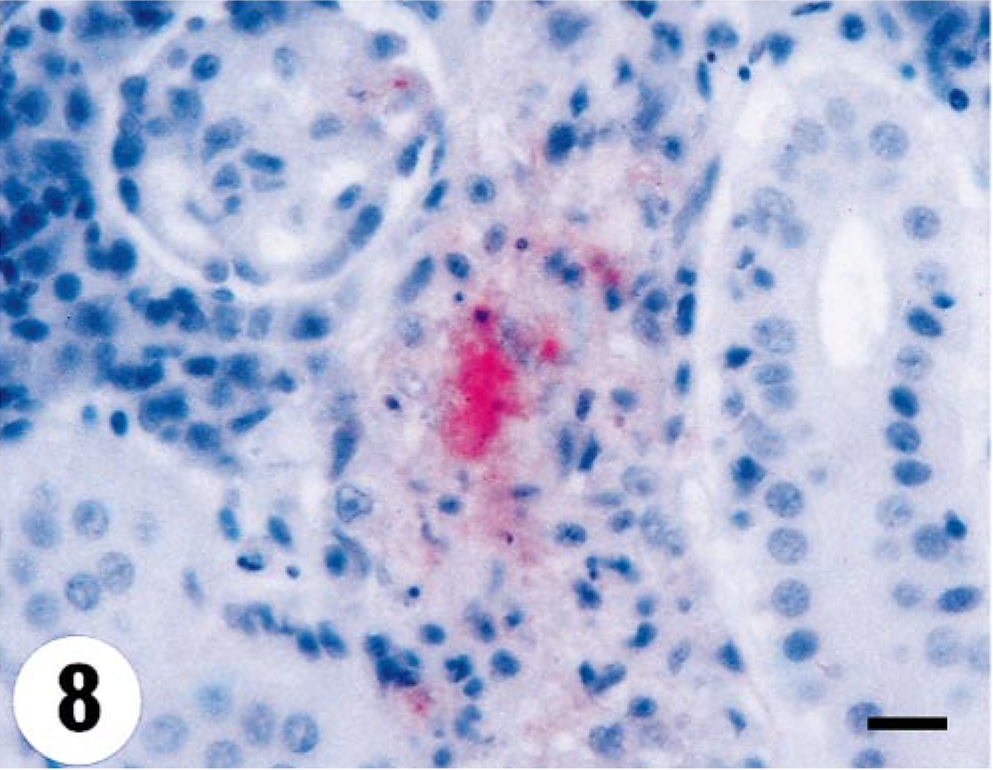
Kidney; turbot (Scophthalmus maximus), infected with 4p168 at 7 days p.i. Focal necrosis with associated VHSV cytoplasmic staining in the interstitium. Virus antigen is detected by alkaline phosphatase–coupled antibodies and observed by Fast Red. The slide is counterstained with Mayer's hematoxylin. Bar = 10 µm.
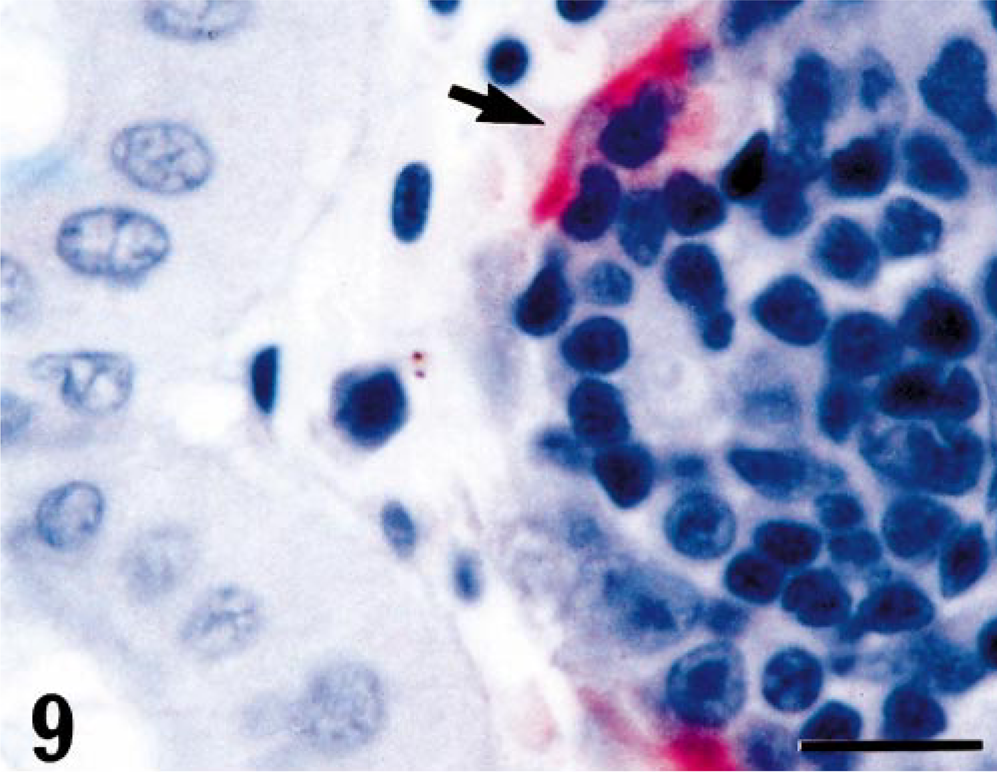
Kidney; turbot (Scophthalmus maximus), infected with 4p168 at 7 days p.i. Strong positive staining for VHSV in the endothelial cells lining the portal sinusoids (arrow). Virus antigen is detected by alkaline phosphatase–coupled antibodies and observed by Fast Red. The slide is counterstained with Mayer's hematoxylin. Bar = 10 µm.
At day 15 p.i., two fish were positive both in heart and kidney, whereas a third fish was positive in the heart only. At day 20, two fish were positive both in heart, kidney, spleen, gills, and skin and one in heart only. At both sampling intervals, extensive pathologic changes were observed in the kidney. Leukocytes and hematopoietic elements of kidney and spleen were positive and with histomorphologic signs of degeneration and necrosis, typically observed as indistinct cell borders and initial fragmentation of the nucleus (Fig. 8). The tubules of the kidney were negative for virus and had no pathologic change.
The viral staining in spleen was located in white pulp and ellipsoids. The pathologic changes developed from single-cell necrosis to multifocal necrosis. In heart, virus was present in myocardial and endothelial cells, and focal necrosis of these cells was observed. Although the ventricle was strongly positive for virus, the bulbus arteriosus was consistently negative. The liver exhibited multifocal virus in hepatocytes with focal degeneration and necrosis. Virus was detected sporadically in the endothelia of the central venous sinus of primary lamellae (Fig. 10). In skin, positive viral staining was observed in the cytoplasm of collagen fibers in dermis, without any pathologic changes being observed. All fish were negative at days 25 and 45 p.i.
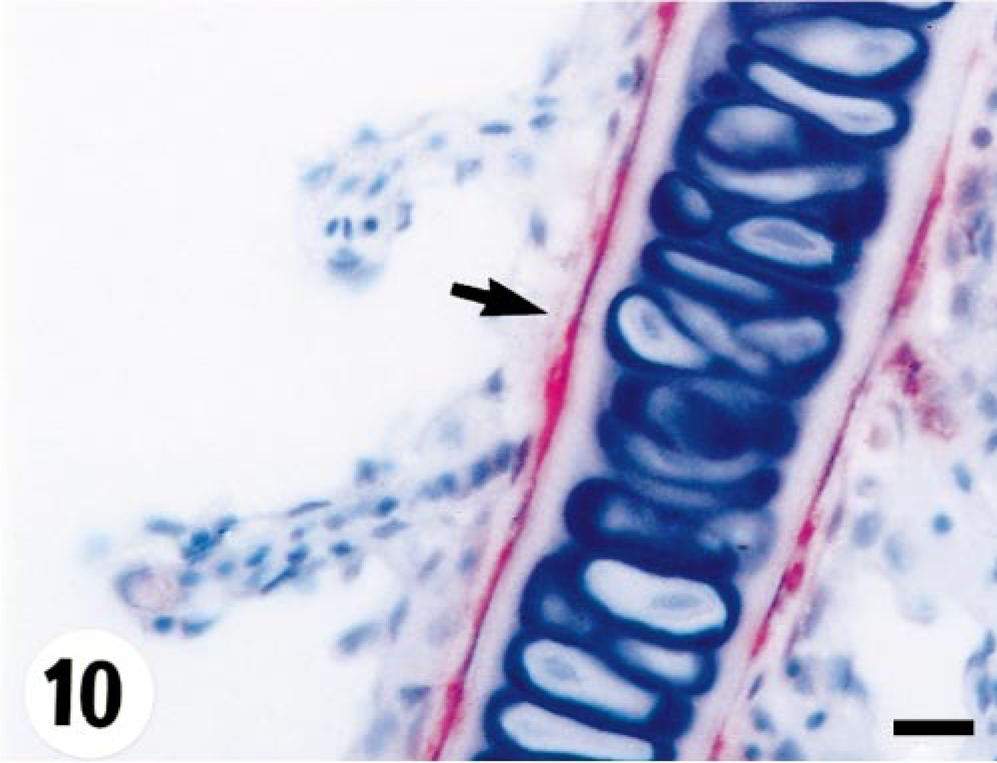
Gills; turbot (Scophthalmus maximus), infected with 4p168 at 20 days p.i. Strong staining of endothelial cells (arrow) in the central venous sinus of the primary lamella. Virus antigen is detected by alkaline phosphatase–coupled antibodies and observed by Fast Red. The slide is counterstained with Mayer's hematoxylin. Bar = 10 µm.
Group 3—herring (1p3) isolate
One fish was found positive by immunohistochemistry in the heart on day 10 p.i. in this group (Table 3). This focus in the myocardium featured severe inflammation (Fig. 11).
Identification of VHSV in tissue from turbot infected with herring isolate 1p3.
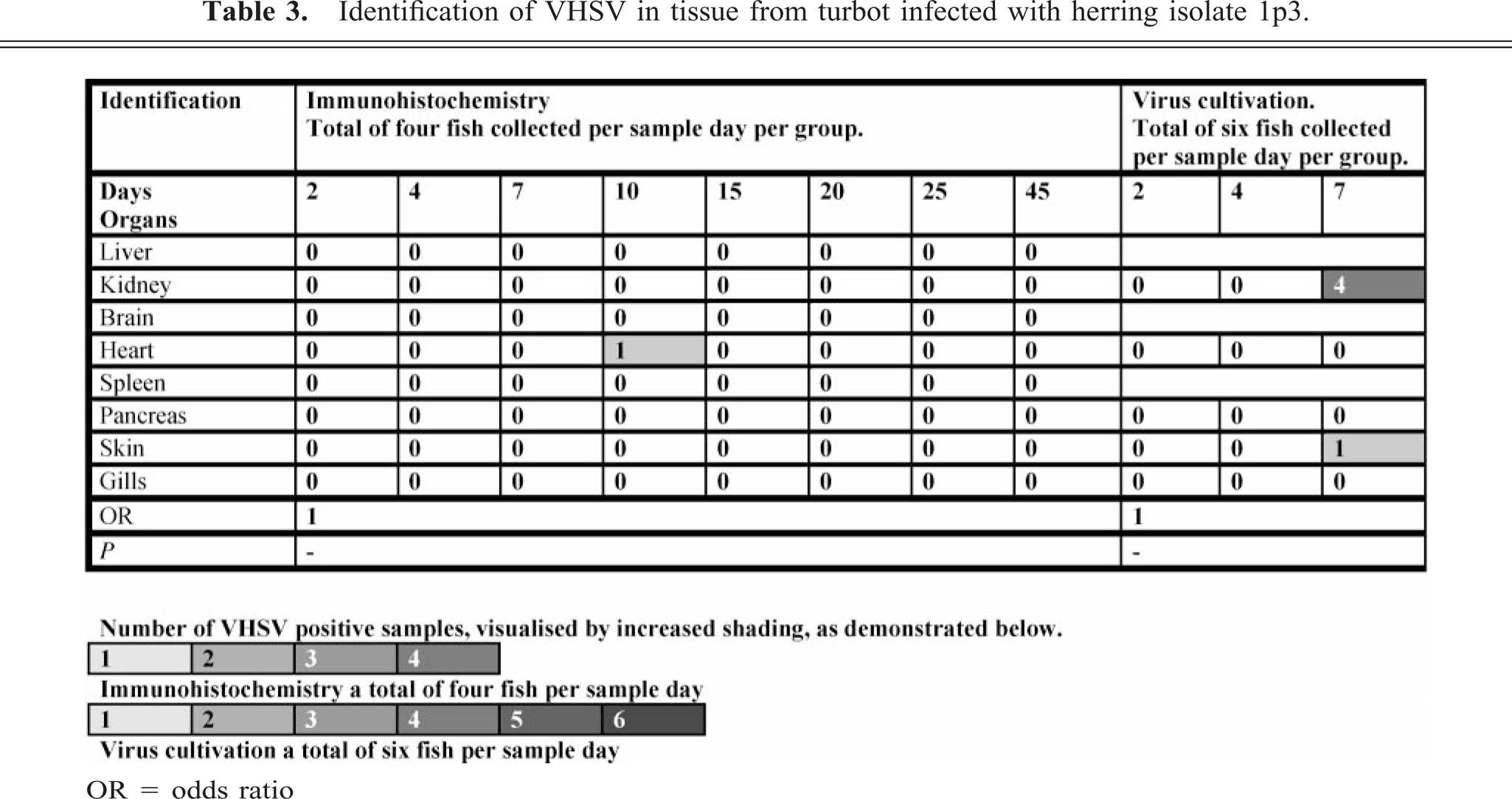
Number of VHSV positive samples, visualised by increased shading, as demonstrated below.
1 2 3 4
Immunohistochemistry a total of four fish per sample day
1 2 3 4 5 6
Virus cultivation a total six fish per sample day
OR = odds ratio
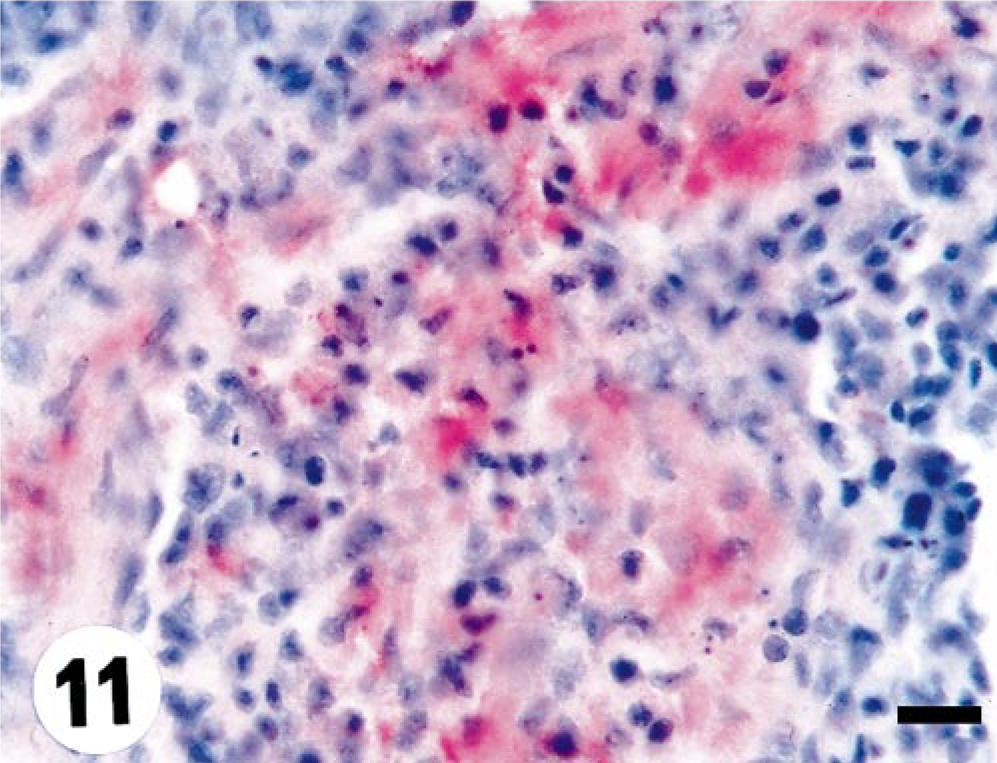
Heart; turbot (Scophthalmus maximus), infected with 1p3 (low-virulence herring isolate) at 10 days p.i. Myocardial fibers with intense cytoplasmic staining (arrow). Note strong infiltration of inflammatory cells. Virus antigen is detected by alkaline phosphatase–coupled antibodies and observed by Fast Red. The slide is counterstained with Mayer's hematoxylin. Bar = 10 µm.
Viral distribution
The immunohistochemical data showed significant difference in the OR between the fish infected with turbot isolate 860/94 and herring isolate 4p168, compared with herring isolate 1p3. The chance of getting a virus-positive specimen is 46.1 and 10.4 times higher for fish infected with the highly virulent isolates 4p168 and 860/94, respectively, compared with the low-virulence isolate 1p3. In contrast, the analysis of the virus cultivation data sampled from days 2 to 7 p.i. did not show significant difference in the OR between the fish infected with turbot isolate 860/94 and herring isolate 4p168, compared with herring isolate 1p3. The data showed that the chance of getting a virus-positive sample is only 1.2 and 1.6 times higher for fish infected with the high-virulence isolates 4p168 and 860/94, respectively, compared with the low-virulence 1p3.
At day 25 p.i., the VHSV titer in kidney and heart is higher for the turbot isolate 860/94 and herring isolate 4p168, compared with herring isolate 1p3. For all three groups, the viral titer is higher in heart compared with kidney (Fig. 1).
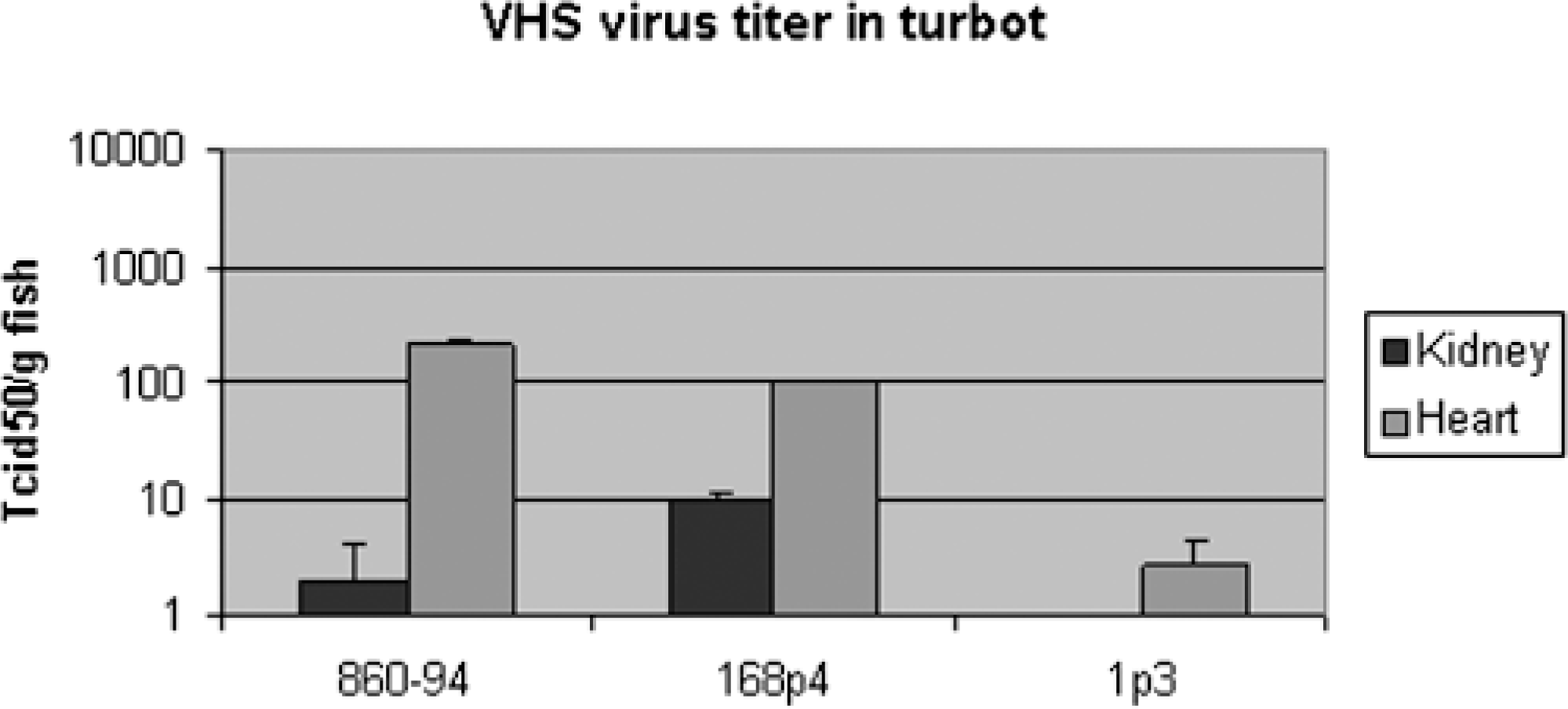
Mean VHSV titer (+ SE) in kidney and heart from turbot 25 days p.i. (n = 6).
Mortality
The mortality rate in the virus-free control group was 0%. The mortality in the duplicate tanks infected with turbot isolate 860/94 was 40% and 43% (Fig. 2). Both tanks had significantly higher mortality compared with the virus-free control and low-virulent herring isolate 1p3 groups but was not significantly different from the high-virulent herring isolate 4p168 group. The mortality in the duplicate tanks infected with herring isolate 4p168 was 53% and 43%. Both tanks had significantly higher mortality compared with the virus-free control and low-virulent herring isolate 1p3 groups but was not significantly different from the high-virulent turbot isolate 860/94 group. The mortality in the duplicate tanks infected with herring isolate 1p3 was 0% and 7% and was not significantly different in terms of mortality compared with the virus-free control group.
VHSV challenge of turbot (mean weight 10 g). Mortality recorded in two tanks per group.
Discussion
The mortality observed in this study was significantly higher for both the high-virulence herring and turbot VHSV isolates (48% and 41.5%, respectively) compared with the virus-free group. In the group infected with the low-virulence herring isolate, the mortality was very low (3.5%). These data are consistent with studies by King et al., 14 who reported 68.6% mortality with herring isolate 4p168, 66.3% mortality with turbot isolate 860/94, and 16.5% mortality with herring isolate 1p3 by bath challenge.
The sequential pathology of marine VHSV in experimentally infected turbot has not been reported previously in detail. Pathologic changes described in turbot infected during farm outbreaks include necrosis of myocardium and hepatocytes, focal hemorrhage of the skeletal musculature and retina, 21 hemorrhage of the intestinal tract, 19 and hemorrhages of skin and liver. 22 The recognized anatomic predilection sites for fish rhabdovirus for piscine rhabdoviruses has been identified as the hematopoietic tissue of the kidney and spleen, as described for spring viremia of carp virus, 1 VHSV 16 and IHNV 30 Hirame rhabdovirus. 9 This study supports these observations and also identifies the heart as a primary organ for localization and replication of the marine VHSV isolates. The pathologic changes observed in fish infected with marine VHSV were most prominent for turbot infected with herring isolate 4p168 and turbot isolate 860/94, concordant with these two isolates also being more virulent. The microscopic lesions were localized in the target organs, heart, kidney, and spleen. Immunohistochemistry revealed strong positive staining for VHSV in the myocardium, with associated degenerative and inflammatory changes. This observation is consistent with previous studies. 21 In the early stage of infection, VHSV antigen was identified in single hematopoietic cells and periportal endothelia. The kidney showed single positive leukocytes in the interstitium. In the later stages the infection progresses to multifocal necrosis of hematopoietic elements. In contrast, splenic involvement was more restricted with primary single cell involvement, with occasional degeneration and necrosis noted in select fish.
Infection with the high-virulence isolates caused a widespread distribution of virus in internal organs resembling that of freshwater VHSV infection in rainbow trout (Oncorhynchus mykiss). 3 The most prominent differences with respect to histopathology between rainbow trout and turbot are the delayed onset of mortality and more restricted distribution of the virus in the internal organs. For the low-virulence herring isolate (1p3), virus was identified in one fish only, but then also with concomitant occurrence of pathologic changes in only one fish that featured myocardial degeneration and necrosis. It has been demonstrated that the endothelial cells of the kidney in rainbow trout are probably the first cells to become infected. 3 The same holds true with a similar predilection site for turbot. It is also noted that in turbot, endothelial cells of the heart and the kidney are VHSV positive at the early stage of infection. In the later stages of infection, virus spreads from the reticuloendothelial to the interstitium with subsequent hematopoietic involvement. This seems to be a typical finding for the high-virulence isolates, but the distribution of virus in internal organs is more limited for the low-virulence isolate as demonstrated by the OR values from the immunohistochemical data. It should be noted, however, that no significant difference in results from virus cultivation was found between the three isolates. Methods sensitivity is potentially a limiting factor, and use of more sensitive methods like quantitative polymerase chain reaction would possibly detect and quantify virus at levels below what can be detected by cell culture. Based on these findings it seems that experimental infection with the low-virulence herring isolate resulted in a subclinical infection without major pathologic changes occurring in the examined internal organs. Despite a comprehensive sampling list, not all fish tissues were evaluated (spinal cord, lateral line, etc). The importance of early VHSV replication in macrophages of rainbow trout has been reported 4 , 6 and macrophages are considered the prime replication sites. However, these studies were mainly based on in vitro 2 data, and in vivo 4 studies indicate that endothelial cells are the prime sites of infection and replication. Furthermore, both kidney and heart are target organs as demonstrated by the immunohistochemical data. This is consistent with the virus isolation data from heart and kidney at day 25 p.i., indicating a higher virus titer in turbot infected with the high-virulence isolates 860/94 and 4p168. The port of entry for the virus has not been finally determined, but it is conceivable that different routes are used. These include the gills, skin, or intestine, although the general opinion advocates the gills to be the prime portal of entry when virus is transmitted in water. 28 Neukirch and Glass 20 isolated virus from gill samples during the initial phase of a water-borne experimental infection. More recently, the epidermis and epidermal tissues were proposed as possible sites for entry and early virus replication. 29 Virus cultivation data from days 2 to 7 in this study do not give any information as to which of the examined organs are most important as a point of entry for marine VHSV. However, there was no indication for earlier replication of virus in gill, skin, or intestine, compared with what are believed to be the target organs, heart and kidney. The data show that virus is detected in skin for both the high-virulence isolates 860/94 and 4p168 at day 7 p.i. It is difficult to verify if these findings indicate viral infection in the skin or only viral attachment to the skin. However, viral replication in epidermis is unlikely because this would have been detected in the immunohistochemistry skin samples. The conclusion is drawn with some caution because a limited area of the skin was examined by immunohistochemistry.
The VHSV isolates 4p168 and 860/94 are of the genotype 3 with a geographic origin of the North Sea and the Eastern Atlantic, 25 whereas the isolate 1p3 is of the genotype 1b, which is primarily a Baltic marine isolate and resembles the freshwater isolates. The separation of genotypes was based on sequence data of the nucleocapsid (N) gene. At amino acid level, differences at position 83 and at position 99 defined geographic isolates. 25 The implications of these findings with regard to the virulence of these different strains is not known because the N gene encodes for the nucleoprotein or the inner protein of the virus. It is therefore unlikely that this protein is directly involved in the pathogenicity of the virus; however, further studies are needed to elucidate a possible correlation between genetic differences and virulence of the marine VHSV isolates.
The main conclusion to be drawn from this study is that virulent marine VHSV isolates induce pathologic changes in turbot comparable with those seen in rainbow trout infected with freshwater VHSV isolates. The viral distribution is also similar, with the initial identification of VHSV in the endothelial cells of the kidney followed by a spread to the target organs heart, kidney, and spleen.
Footnotes
Acknowledgements
We thank the Aquarium Services Group at FRS Marine Laboratory, Aberdeen, for their assistance in aquarium sampling. This study was supported financially by the European Commission, project no. FAIR CT-96-1594.