
Abstract
Five camelid mucocutaneous fibropapillomas with histologic features similar to equine sarcoids were diagnosed. They were characterized by a dermal fibroblastic proliferation and overlying, often ulcerated hyperplastic epidermis with thin rete pegs extending down into the dermis. Two of the tumors came from llamas and three from alpacas. Four of the animals were 6-year-old females. The fifth was a 6-year-old castrated male. The fibropapillomas were located on the nose, lip, and cheeks. One of the llama tumors waxed and waned before surgery and recurred and spread after surgery. None of the other tumors recurred. All five tumors were positive for papillomavirus (PV) DNA by polymerase chain reaction testing. Nucleotide sequence analysis of the PCR product from one of the llama fibropapillomas confirmed a unique PV. This report provides the microscopic and clinical features of fibropapillomas in camelids as well as evidence for a PV etiology.
Benign sessile fibroblastic proliferations with overlying epithelial hyperplasia, hyperkeratosis, and long, thin rete ridges in which the fibrous proliferation predominates and the epithelial proliferation may be slight have been referred to as fibropapillomas. 19,20 Others refer to these lesions as fibromas; 9 however because the term fibroma does not indicate the epithelial proliferation characteristic of these lesions, fibropapilloma is the term used in this article. These lesions are usually attributed to papillomavirus (PV) infection and have been best studied in cattle and horses. In horses, fibropapillomas have been referred to as sarcoids. A PV association has been documented for fibropapillomas in cattle, 2 equids, 1,3 , 11–14 , 18 sheep, 5 cervids, 9 a pronghorn antelope, 16 and most recently in cats. 15
Some camelids (llamas) are popular pets and others (alpacas) are raised for their wool. Consequently, biopsies from these animals are seen with increasing frequency at diagnostic laboratories. This report documents the occurrence of mucocutaneous fibropapillomas in this genus and the presence of PV in these lesions.
Five mucocutaneous fibropapillomas from two llamas and three alpacas were collected from biopsy specimens submitted to Marshfield Laboratories, Marshfield, WI and Phoenix Central Laboratories, Everett, WA from April 1998–August 2001. The prevalence of these lesions cannot be determined because the number of camelid lesions for that time period is not known. Two, haired skin biopsy specimens diagnosed with nonspecific proliferative dermatoses, one from a llama and one from an alpaca, served as controls.
Tissue sections were processed routinely, paraffin embedded, sectioned at 5 μm, and stained with hematoxylin and eosin.
Polymerase chain reaction (PCR) testing was conducted on the five formalin-fixed, paraffin-embedded tumors and on two camelid control tissues as previously described. 8,15,17 PV DNA was detected by consensus PCR of a highly conserved region of the E1 gene using the primer pair PV5 (5′-GGBCCTCCAAAYASWGGVAAD-3′) and PV3 (5′-ATGTDTCAAADTABYTCCAK-3′). 15 The presence of amplifiable DNA in all llama and alpaca specimens was determined using the primer pair ALD5 (5′-GCCTGTTCTGAACTCTAGTTG-3′) and ALD3 (5′-CTGCTACTGGCATTAAAATTAGG-3′) to camelid cytochrome b. The expected product was 140 bp (base pair).
The submitting clinicians were contacted by telephone to find out if there was any other treatment and if there was tumor recurrence.
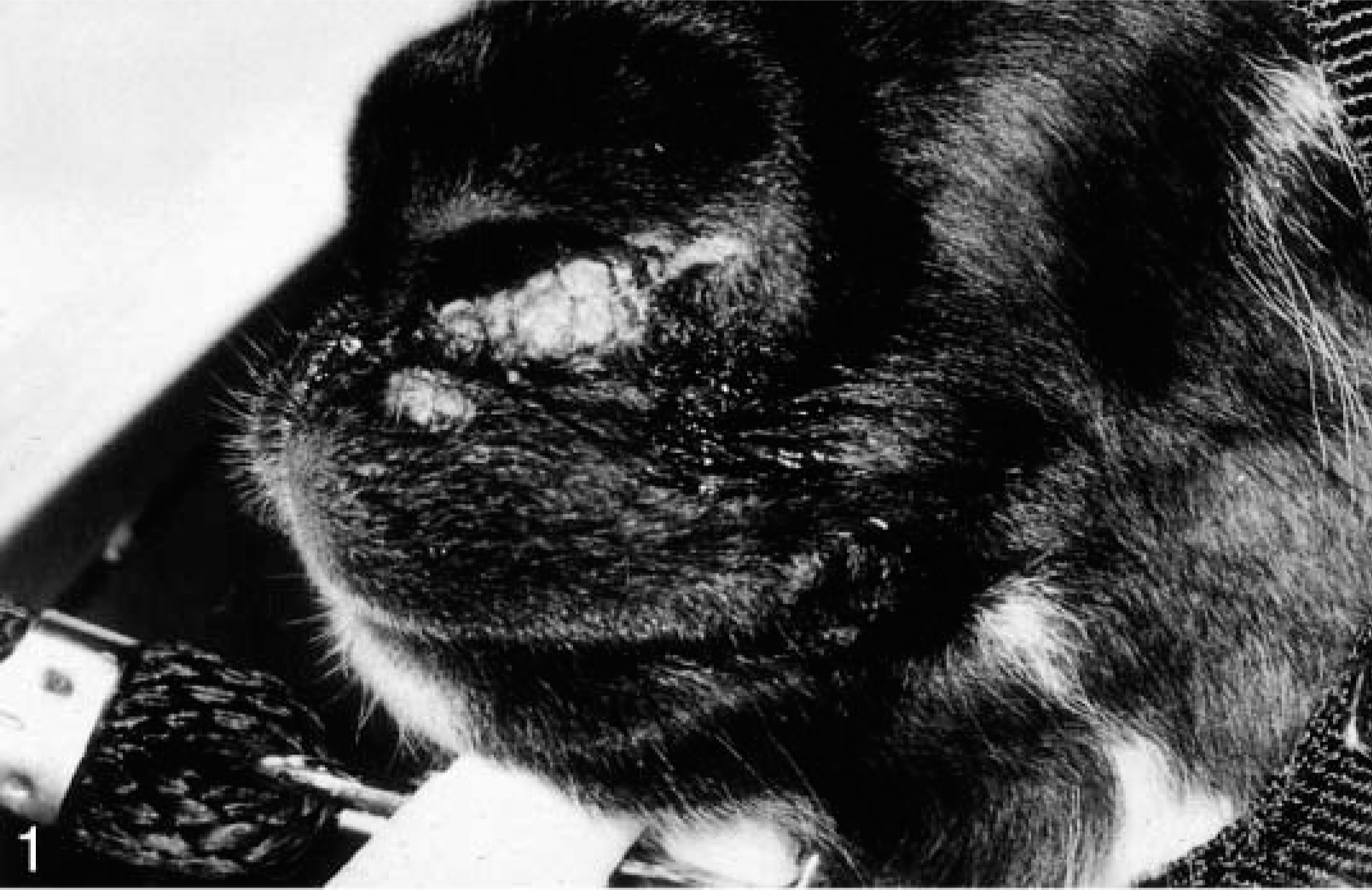
The clinical information and distribution of the lesions are given in Table 1. All the animals were 6 years old. There were four females and one castrated male. All lesions were located on the nose, lip, and cheeks. Grossly, the lesions were gray nodular hyperkeratotic masses (Figs. 1, 2) around the nostrils, on the lips, or on the cheek mucosa.
Signalment, lesion distribution, and follow-up information on camelids with mucocutaneous fibropapillomas.
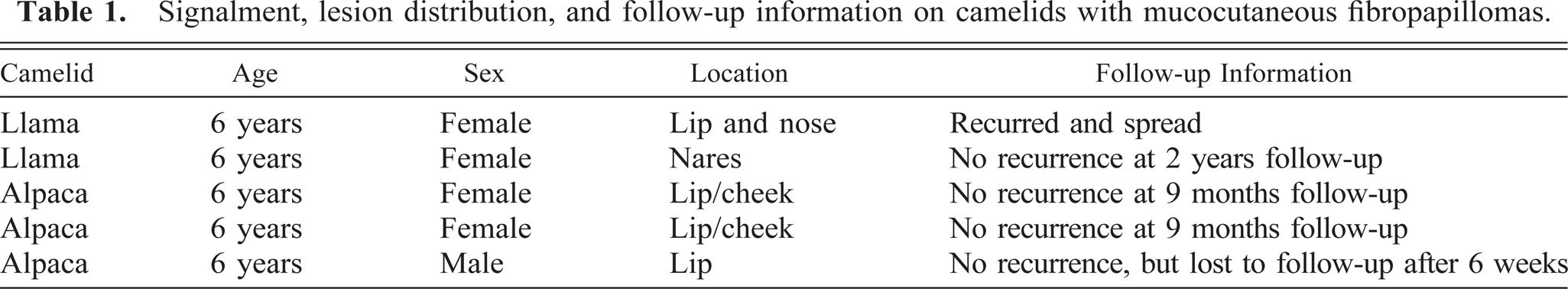
Muzzle; llama. A multinodular hyperkeratotic fibropapilloma of the left naris.
Right side of the face; llama. A similar multinodular hyperkeratotic fibropapilloma involving the right oral commissure of the same llama depicted in Fig. 1.
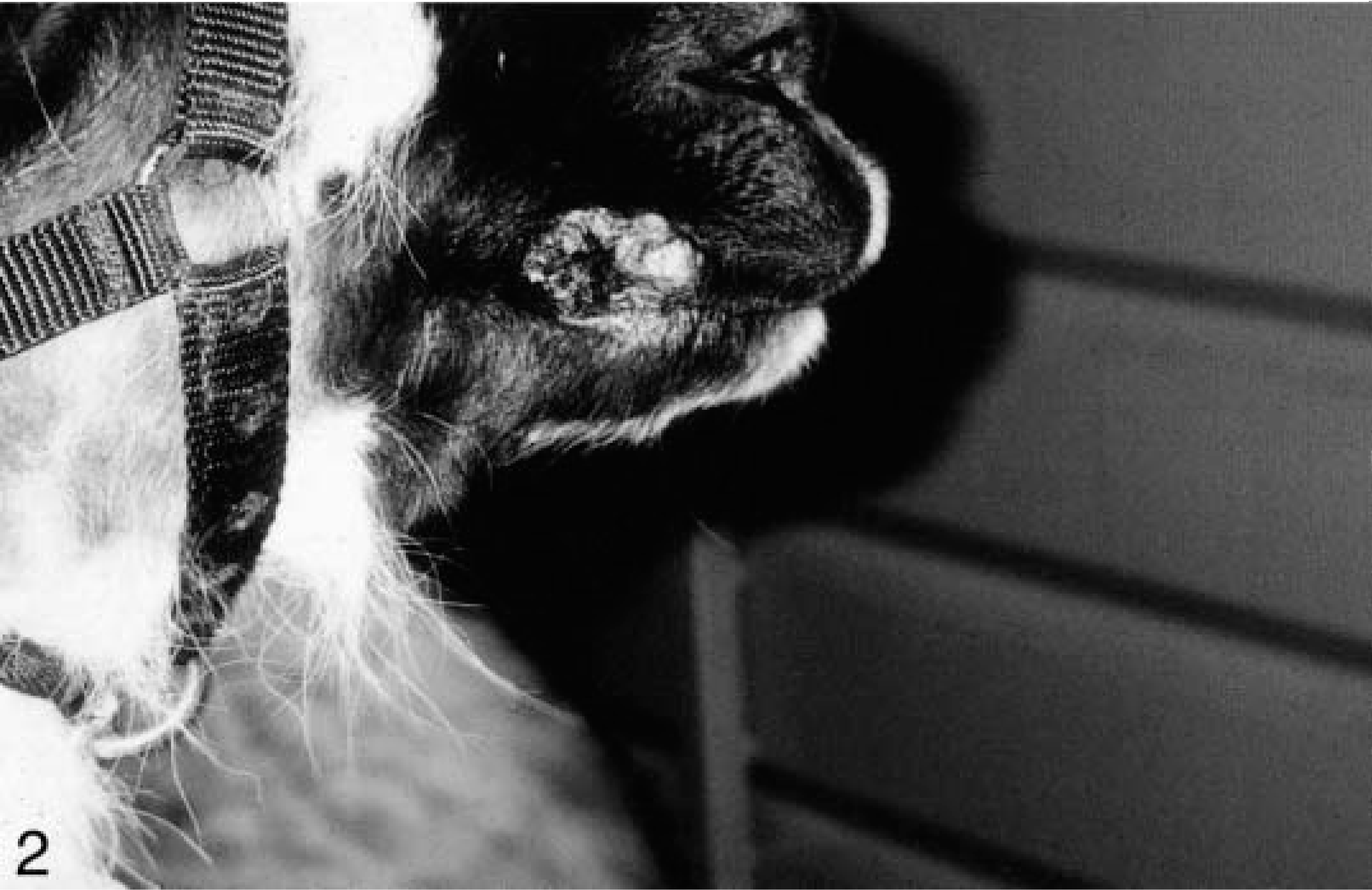
Histologically, the cutaneous tumors were characterized by spindloid to stellate fibroblastic cells in a moderate amount of collagenous matrix that infiltrated and expanded the dermis and subcutis, surrounded and widely separated adnexa, and abutted overlying hyperplastic epithelium which had long, thin rete ridges (Figs. 3, 4). Neoplastic cells were arranged haphazardly and in vague streams and had oval to fusiform nuclei, finely stippled chromatin, small nucleoli, a small amount of eosinophilic cytoplasm, and indistinct cell margins (Fig. 5). The mitotic index was less than one per ten high-power fields. There was mild anisokaryosis and moderate orthokeratotic hyperkeratosis. Two of the skin tumors were ulcerated. The oral mass was a similar neoplasm that expanded the mucosa and submucosa.
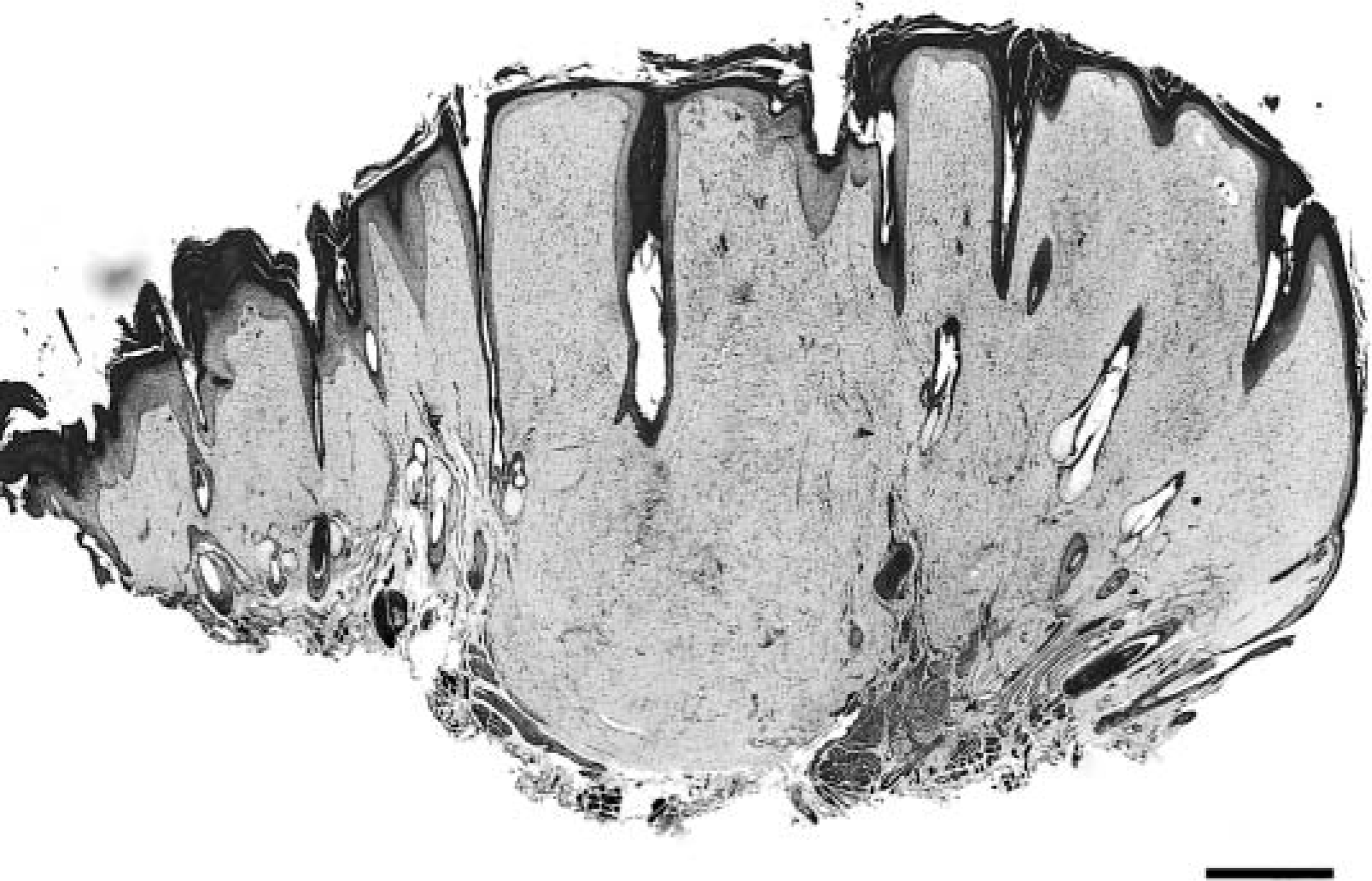
Haired skin; llama. A fibropapilloma expanding the dermis and surrounding and separating adnexa with epidermal hyperplasia and hyperkeratosis. Bar = 0.1 cm.
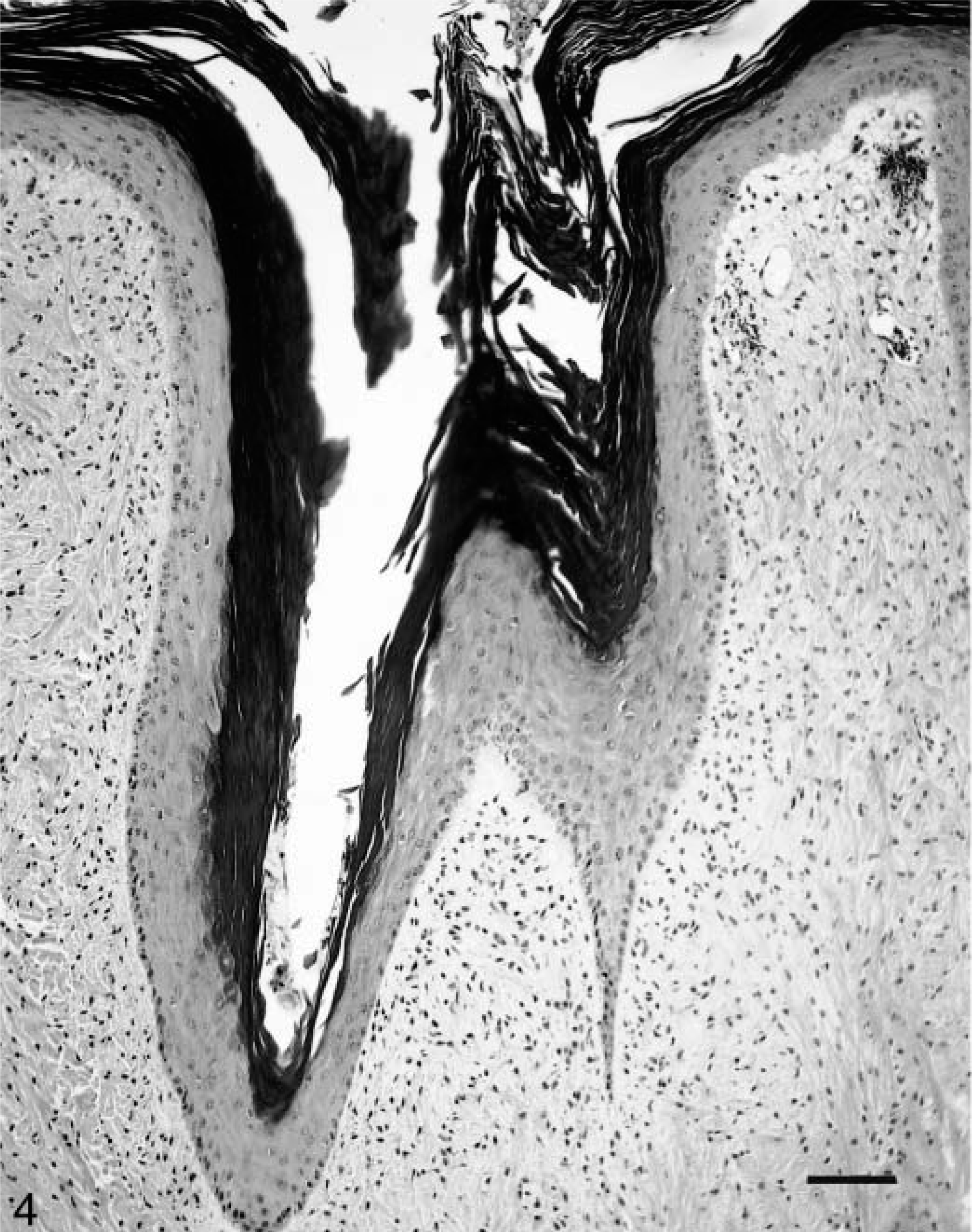
Fibropapilloma; llama. Moderate epithelial hyperplasia and orthokeratotic hyperkeratosis. Bar = 100 μm.
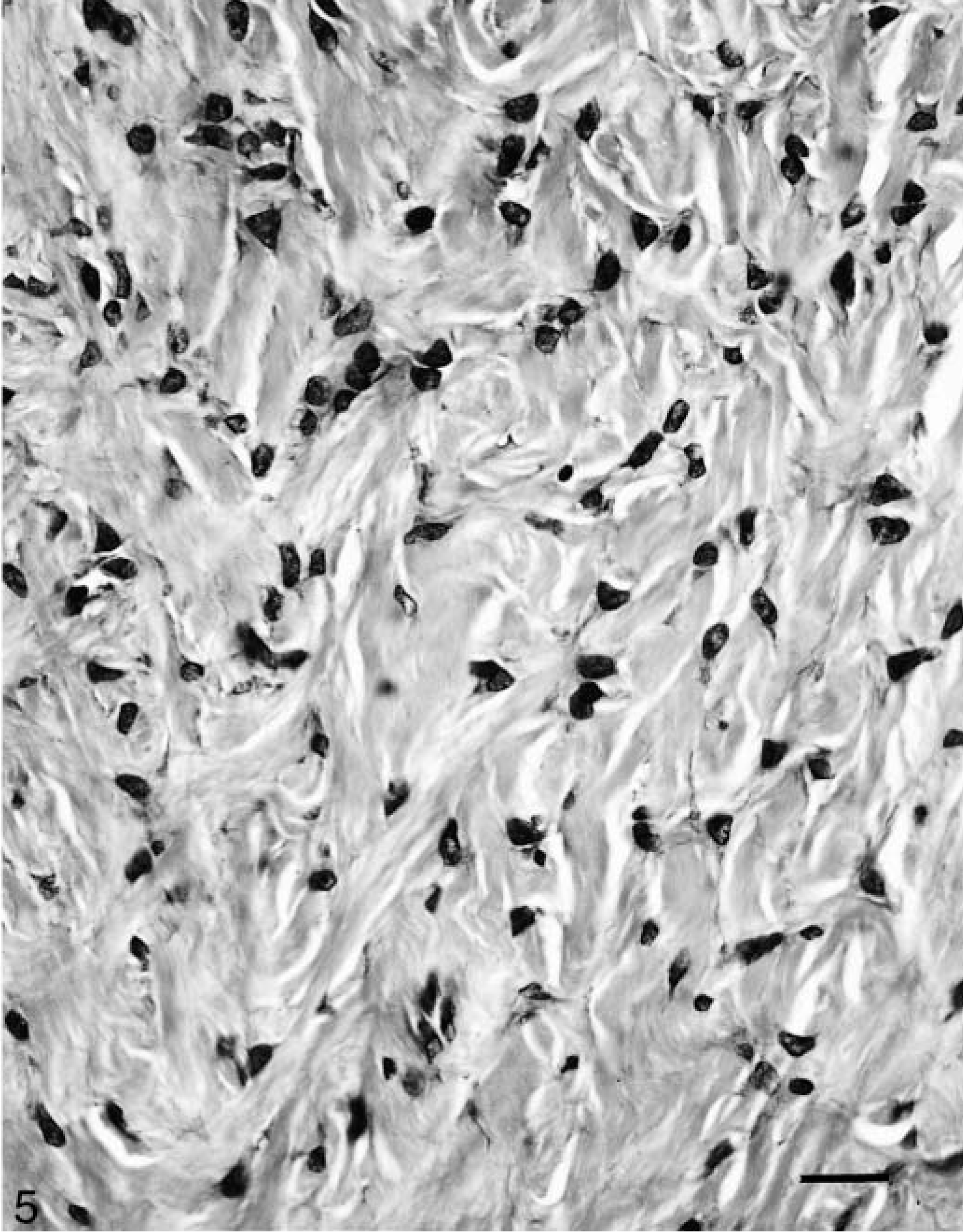
Fibropapilloma; llama. The fibropapilloma is moderately cellular and composed of spindloid to stellate fibroblastic cells in a small to moderate amount of collagneous matrix. Bar = 25 μm.
All five tumors tested had PV DNA detected by consensus PCR. A 176-bp PCR product was obtained from all five camelid fibropapillomas, equine sarcoid tissue, and plasmids containing bovine papillomavirus-1 (BPV1) and canine oral PV (Fig. 6). No PV DNA was detected in the two camelid control skin specimens. Frozen tissue was collected from a recurrence of one of the llama lesions and the 176-bp E1 gene fragment was cloned and sequenced. Nucleotide sequence homologies among the llama PV and other known PVs were determined by BLAST analysis. The 176-bp llama sequence was unique (Fig. 7). It was most closely related to BPV1 with 73% homology. There was 64% homology with canine PV. All other known sequences were more distantly related.
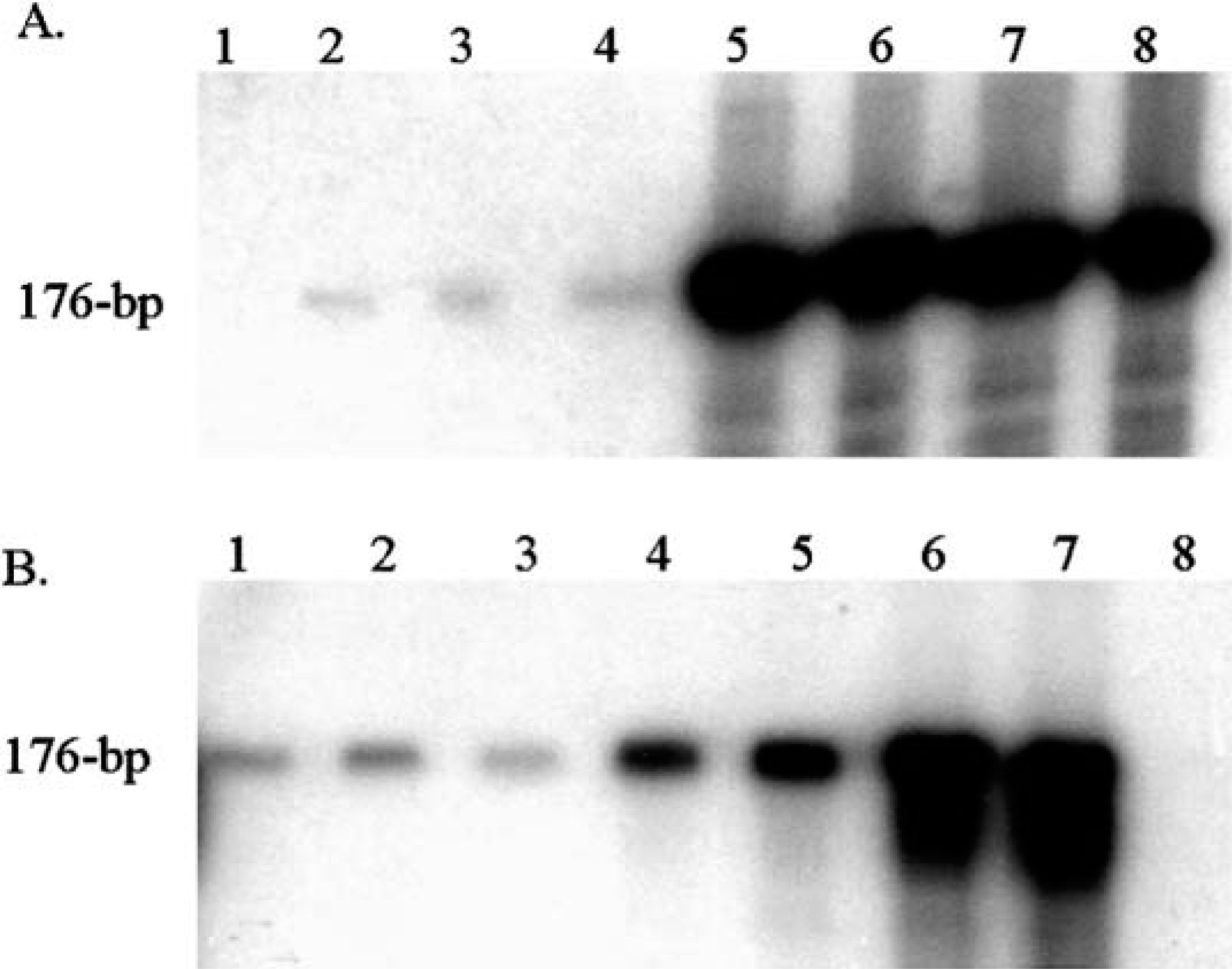
Autoradiograph of P-32–labeled PCR products separated on a 6% denaturing polyacrylamide gel. The expected PV E1 gene product is 176 bp. The gel in A was exposed for 18 hours; B was exposed for 6 hours. The samples in the gel lanes of A are 1, water; 2–4, three dilutions of DNA from an alpaca fibropapilloma; and 5–8, four dilutions of DNA from a llama fibropapilloma. The controls in B are 1–3, equine sarcoid DNA; 4 and 5, canine PV DNA; 6 and 7, BPV1 DNA; and 8, water.
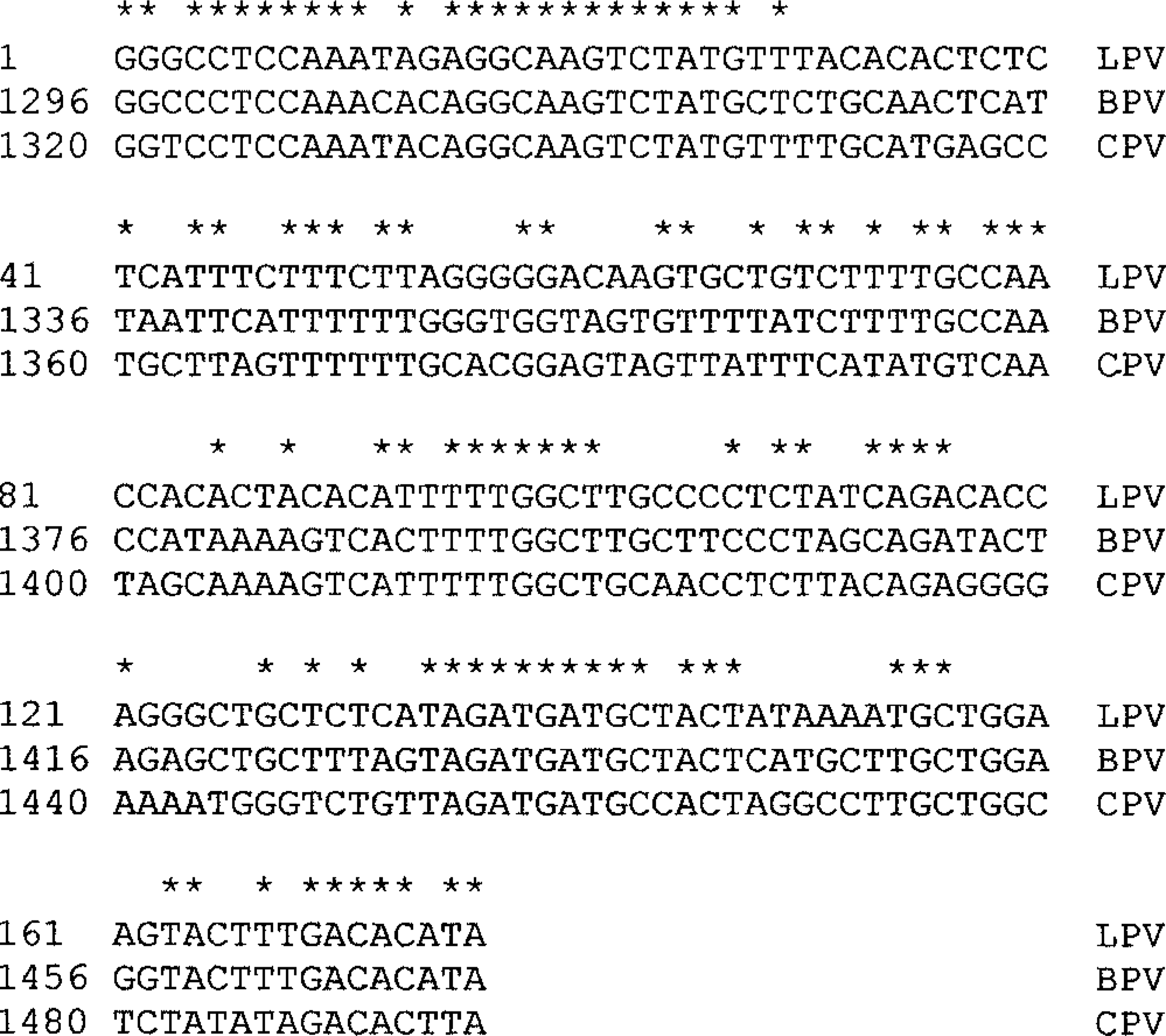
Nucleotide sequence alignment of PV E1 gene fragment found in a llama fibropapilloma (LPV) compared with BPV1 and canine PV. Conserved bases are indicated with an asterisk.
Follow-up information was obtained from 6 weeks to 2 years after surgery. One llama had recurrence and spread of the tumors. None of the other lesions recurred.
Although the numbers in this study are small, these camelid fibropapillomas have many similarities with viral-associated fibropapillomas (sarcoids) in other species. Histologically, the camelid tumors and fibropapillomas of other species are essentially identical, characterized by a fibroblastic proliferation with overlying epithelial hyperplasia and rete ridges. Fibropapillomas are usually seen in young animals. Affected cattle are usually less than 2 years old. 19 Most cats with fibropapillomas are less than 5 years old 15 and, in the horse, these tumors are usually initially reported in animals between 3 and 6 years of age. 6,10 All these camelids were 6 years old. Sarcoids appear most commonly on the head, neck, ventral abdomen, and distal legs of horses 6,7,10 and the head, neck, and digits of cats. 15 All the camelid tumors were on the head. Viral-associated fibropapillomas can recur, spread, and occasionally spontaneously regress. One of the five camelids had recurrence and spread of the tumor. Finally, there is a strong association with PV, suggesting a causal relationship. All five tumors tested had PV DNA.
In cattle, there are at least six types of PVs. BPV types 3, 4, and 6 cause epithelial papillomas, whereas fibropapillomas are attributed to BPV types 1, 2, and 5. BPV was first implicated in the pathogenesis of sarcoids in 1951. 11 Since that time many studies have provided additional evidence that BPVs (BPV1 and BPV2) play a role in the development of sarcoids. 1,3,4 , 12–14 , 18 Recently, expression of the BPV transforming gene (E5) was identified in sarcoids providing the strong evidence for a causal relationship. 3
PV-associated fibropapillomas have now been identified in many other species. 5,9,15,16 Interestingly, the associated PVs are consistently most closely related to BPVs. Most fibropapillomas in horses contain identifiable BPV DNA, either type 1 or type 2. 1,4,12,14,18 The E1 gene of the PV identified in donkeys is very similar to BPV1 and proposed to be a subtype of BPV1. 14 The PVs isolated from white tail and mule deer fibropapillomas have a high degree of relatedness to BPV1 and BPV2. 9 In a recent report on feline cutaneous fibropapillomas, the E1 gene of the isolated PV had 75% homology with BPV1. 15 The PV recovered from the llama tumor had 73% homology with BPV1.
All five camelid fibropapillomas have a very similar clinical presentation suggesting that when a 6-year-old female llama or alpaca presents with a hyperkeratotic gray nodular tumor on the nares, lips, or cheek, fibropapilloma should be high on the differential diagnosis. Additional studies on the transmission of PVs and possible role in oncogenesis are needed to direct clinical management and treatment of these tumors.
Footnotes
Acknowledgements
We thank all the clinicians and owners for their willingness to contribute to this study. Ms L. Zimmermann of Marshfield Laboratories, Marshfield, WI, provided invaluable logistic support. The opinions and assertions contained herein are the private opinions of the authors and are not to be construed as official or as reflecting the opinions of the Department of the Army or Department of Defense.