
Abstract
Anionic peptides (APs) are small antimicrobial peptides present in human and ovine lung. In this study APs were also detected in bovine lung, and production of APs in lungs with acute inflammation induced by various stimuli was determined. The distribution and intensity of APs were determined by immunohistochemistry in lungs of 1) neonatal calves (1-3 days of age) inoculated with Mannheimia (Pasteurella) haemolytica, a known inducer of the bovine β-defensin lingual antimicrobial peptide (LAP) or pyrogen-free saline (PFS), and 2) growing calves (3 months of age) similarly inoculated with M. haemolytica, a lipopolysaccharide (LPS) from M. haemolytica, an LPS-associated protein from M. haemolytica, or PFS. APs were also detected by western blots with the same antibody in lungs of the calves above, as well as in calves inoculated with Pseudomonas aeruginosa, and an adult cow. Anionic peptide (AP) immunoreactivity was detected in bands (approximate weights) in the western blots of lung at 28-30 (strongest signal), 31, 45, and 52-60 kd regardless of inoculum. The adult cow lacked bands at 45 kd, but it had additional bands at 64 (inconsistently) and 35-38 kd. All these band sizes are consistent with those of the western blots of human and ovine lung. The cellular distribution of APs in lung of neonatal and growing cattle was similar to that in lung of human and sheep. In lungs with acute inflammation induced by live bacteria, LPS, or protein, AP distribution and intensity were similar to those in control (PFS-inoculated) lungs and slightly decreased in bronchioles. This work demonstrates that AP is present in lung of cattle and is thereby conserved among two ruminant species and man. Distribution and intensity of AP production are not enhanced by infection or acute inflammation and are decreased in bronchioles, which suggests that AP is not induced like β-defensins such as LAP, but, instead, is produced constitutively.
Because of the close association of respiratory mucosa to environmental substances, innate defense mechanisms exist, at least in part, to protect the host against microbial colonization and infection. These mechanisms include the mucociliary apparatus, surfactant and surfactant proteins, alveolar macrophages, and an intact epithelium. Recently, several classes of antimicrobial proteins and peptides (AMP) produced by respiratory mucosal cells have become increasingly appreciated because of their contribution to innate defense against microbial pathogens.9,13,16,22 These include peroxidase, lactoferrin, lysozyme, bacterial permeability–inducing protein-1, phospholipase A2, secretory leukoprotease inhibitor, defensins, and cathelicidins.9,13,22 Expression of some of these, notably the human cathelicidin LL3712 and human β-defensin-2 and −3 (HBD 2, 3 = peptide; DEFB 2, 3 = gene),19,22 tracheal antimicrobial peptide (TAP)10,20 and lingual antimicrobial peptide (LAP)20–23 of cattle, mouse β-defensin-3 (MBD-3)1 and rat β-defensin-2 (RBD-2),17 is induced or potentially induced by inflammation. Brogden et al.2,4,7 have determined that a class of peptides with anionic properties also has antimicrobial activity and is produced by epithelial cells of lung of sheep and humans. Anionic peptide (AP) production during bacterial pneumonia or in response to inflammatory stimuli has not been fully characterized in any species but has recently been shown to be decreased in alveolar epithelium and bronchoalveolar lavage fluid (BALF) from lungs of humans with cystic fibrosis6 and increased in the cytoplasm of hyperplastic cells during chronic pneumonia of sheep.11
APs are distinct from many AMPs for the following reasons: they are small, they are anionic because of homopolymeric regions of aspartate, they require zinc as a cofactor for bactericidal activity, and they are present in pulmonary surfactant.4,6,7 It has been suggested that APs are products of posttransitional cleavage of a larger peptide, such as a zymogen.7 The identity of the mature protein product from which APs are cleaved is not yet known. Similar to other AMPs, APs are bactericidal against Gram-positive and Gram-negative organisms. Efficacy of AP microbial killing is inversely proportional to the number of aspartate residues in the peptide.4,7 The mechanism of bacterial killing has not been fully elucidated but is thought to involve the bacterial uptake of APs and zinc. AP bactericidal activity is inhibited in the presence of phosphates or ethylenediaminetetraacetic acid, but they have increased antimicrobial activity with increasing concentrations of NaCl.4,18 Recent work indicates the presence of larger AP-containing proteins in the trachea, nasal epithelium, liver, and small intestine of sheep.5 Degenerate oligonucleotides were synthesized to identify precursor genes for APs in sheep genomic DNA; these probes hybridized with larger sequences, further indicating the likelihood that APs are precursor portions of a zymogen.4,5 AP has been detected in BALF from neonatal and adult cattle by enzyme-linked immunosorbent assay (ELISA), and the percent killing of APs in BALF from adult cattle (n = 11) ranged from 23% to 75% (60.2% average) in a dilution susceptibility test.8 But cellular production of AP in bovine lung has not been previously assessed. Bovine respiratory epithelial cells produce at least two β-defensin antimicrobial peptides, TAP and LAP, which are induced by inflammatory mediators.20–23 Of these, LAP mRNA expression is markedly increased during infection with the ruminant respiratory pathogen Mannheimia (Pasteurella) haemolytica.23 Both M. haemolytica LPS and an LPS-associated peptide (LPS-P) also induce inflammation in this model.3
The purpose of this study was to assess the cellular distribution and intensity of AP production in lung during an acute inflammatory response induced either by a live M. haemolytica or by bacterial products (LPS, LPS-P). We assessed the presence of APs in healthy and inflamed lung of neonatal and growing (3 months old) calves, ages at which human infants and other mammalian species are particularly susceptible to microbial infection.
Materials and Methods
Animals and tissues
Eight healthy male Holstein calves were deprived of colostrum and brought to the Iowa State University Laboratory Animal Resources facility within 24 hours of birth. Four animals each were inoculated with 5 ml of live M. haemolytica (1.55 × 109 CFU) or pyrogen-free saline (PFS) in the primary bronchus of the right cranial lung lobe by fiberoptic bronchoscopy and euthanatized 6 hours after inoculation. Colostrum-deprived, neonatal calves did not survive up to 24 hours after inoculation with M. haemolytica and were, therefore, euthanatized 6 hours after inoculation. In addition, eight male Holstein calves, 3 months of age, were similarly inoculated in the primary bronchus of the right cranial lung lobe by fiberoptic bronchoscopy with M. haemolytica, M. haemolytica LPS, M. haemolytica LPS-P, or PFS (two calves per inoculum) and euthanatized 24 hours after inoculation.
From each animal of each age group (neonatal and 3 months old), sections of lung were taken from each inoculated lung lobe and fixed in 10% neutral-buffered formalin for immunohistochemical detection of APs. Lung tissue collected 6 hours after inoculation from neonatal calves inoculated intrabronchially with either PFS or M. haemolytica, as well as lung tissue collected 4 hours after inoculation with Pseudomonas aeruginosa (included to compare two different Gram-negative bacteria), killed P. aeruginosa, or P. aeruginosa LPS, and lung tissue collected from one healthy, uninoculated adult cow (1 cm3/sample, two samples per animal) were frozen immediately in liquid nitrogen and stored in a −70 C freezer for western blot analysis.
Antianionic peptide monoclonal antibody (IG9-IC2)
Antianionic peptide monoclonal IgG was produced by Brogden et al.4,7 in CF1 mice against the synthetic peptide H–DDDDDDD–OH conjugated to keyhole limpet hemocyanin. This peptide was synthesized on the basis of the sequence of APs isolated from sheep BAL fluid.4,7
Western blot analysis
Lung tissue was homogenized in chilled 0.14 M NaCl and then boiled for 5 minutes after addition of reducing sample buffer (ImmunoPure® Reducing Lane Marker Buffer, Pierce, Rockford, IL). Twenty microliters of a 1:10 dilution (lung sample/1× lane marker buffer) was loaded per lane. Broad-range sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) molecular weight standards (BioRad®, Hercules, CA) were run in two lanes per gel. SDS-PAGE was performed with 4% stacking and 12% resolving precast mini-gels (BioRad®) run at 100 V for 90 minutes. Proteins were transferred to polyvinylidene difluoride (PVDF) membranes (Immobilon-P, Millipore Corporation, Bedford, MA) by electroblotting (30 V for 16 hours). Primary antibody (mouse anti-AP IgG) was applied at a concentration of 5 µg/ml (1:200 dilution) to the PVDF membranes. Secondary antibody was goat anti-mouse IgG conjugated with alkaline phosphatase (Kirkegaard & Perry Laboratories, Gaithersburg, MD) used at 0.5 µg/ml (1:2000). The blots were incubated with peroxidase chromagen (Kirkegaard & Perry Laboratories) for 5 minutes. The western blots were graded by the presence or absence of immunoreactive bands; molecular weights of bands were estimated by broad-range SDS-PAGE molecular weight standards (BioRad®). Lung tissue from a healthy adult cow was included in the event that lung tissue of neonatal cattle lacked anionic peptide immunoreactivity (AP-IR).
Anionic peptide immunohistochemistry
Briefly, slides were deparaffinized, rinsed with ultrapure water, subjected to antigen retrieval (microwaved for 10 minutes in pH 10.0 Tris buffer), and then placed in a −20 C freezer for 15 minutes. Slides were washed in TNTB buffer (0.05 M Tris, 0.85% NaCl, 0.05% Tween, 1% bovine serum albumin [BSA]) and then incubated with mouse anti-AP IgG (1:700) overnight in a humidified chamber at 4 C. Slides were rinsed with TNTB the next morning and incubated for 30 minutes at room temperature with biotinylated goat anti-mouse IgG, 1:400 dilution (Vector, Burlingname, CA). Slides were rinsed in wash buffer (BioGenex, San Ramon, CA), and then alkaline-phosphatase–conjugated streptavidin (BioGenex) was applied for 30 minutes. Slides were rinsed and then exposed to chromagen (Vector Red, Vector) for 18 minutes, rinsed, counter-stained (1/4 strength hematoxylin), and coverslipped. Specificity of antibody affinity with tissue sections was determined by preincubating the primary antibody with varying concentrations of synthetic AP. Three types of negative controls were used, and all lacked immunoreactivity: 1) control sections that were not incubated with primary antibody, 2) the primary antibody was incubated with the synthetic AP (HO–DDDDDDD–H) before tissue application (see also below), and 3) mouse preimmune serum was substituted for primary antibody.
Distribution of AP-IR
The cellular distribution of AP-IR was determined by light microscopy of sections stained by immunohistochemistry, defined, and compared with the AP-IR distribution in human and ovine lung.
Intensity of AP-IR
The intensity of AP-IR was assessed and characterized by light microscopy. Intensity of staining was scored on a subjective scale where 0 = no AP-IR; 1 = <10% of cells (of bronchi, bronchiole, alveolus, vessel) had AP-IR; 2 = 10–30% of cells had AP-IR; 3 = 30–60% of cells had AP-IR; and 4 = 60–90% of cells had AP-IR. Five 20× fields per tissue of lung from at least two sections of lung per animal were scored.
Scoring of inflammation intensity
Inflammation intensity was assessed in a predetermined scoring system at 20× magnification, where 0 = no inflammatory cells, 1 = detectable infiltrates of neutrophils in airways, 2 = >5 neutrophils in >10% and <30% of alveolar and airway lumens, 3 = >5 neutrophils in >30% of alveolar and airway lumens, and 4 = filling of alveolar and airway lumens with exudates. Five 20× fields per tissue of lung from at least two sections of lung per animal were scored.
Anionic peptide specificity
Assessment of antibody specificity
Antibody specificity was previously assessed with a competitive ELISA.6,7,18 Briefly, BSA-DDDDDDD–OH conjugate (50 ng conjugate/well) was used as the adsorbed antigen, and varying graded concentrations (1.0–0.002 mM) of H–DDDDDDD–OH, H–GADDDDD–OH, H–VDDDDK–OH, H–TQDDGGK–OH, H–GGEEK–OH, and H–SGSGSGS–OH were used as competitive antigens in the presence of 1G9-1C2. 1G9-1C2 was specific for peptides with C-terminus Asp residues. The antibody recognized H–GADDDDD–OH nearly as well as H–DDDDDDD–OH. But H–VDDDDK–OH, H–TQDDGGK–OH, H–GGEEK–OH, and H–SGSGSGS–OH were not recognized and did not cross-react.
Identification of antibody epitopes
The epitope-binding site of the antibody was also previously determined.6–8 Thirty peptides (7 residues each) corresponding to a single residue frameshift of the sequence were synthesized simultaneously on a derivatized cellulose sheet (SPOTs, Genosys, Woodlands, TX). Antibody 1G9-1C2 recognized epitopes that contained greater than 2 C-terminal Asp residues.
Immunocytochemistry
The specificity of the monoclonal was previously determined.19 Briefly, control tissue sections were incubated without antibody 1G9-1C2 or with antibody 1G9-1C2 preincubated with 0.5 mM H–DDDDDDD–OH. In both these controls, specific staining of APs could not be detected.
Statistical analysis
Inflammation, intensity, and distribution scores were analyzed by the chi-square test and Fisher's exact test. The significance level used was 0.10.
Results
Western blots of lung from calves and an adult cow
In the western blots of lung homogenates from all neonatal calves (Fig. 1) and an adult cow, five bands were detected regardless of inocula: 28–30 (strongest signal), 31, 45, and 52–60 kd (in calves only); in the western blot of the cow, additional bands were present at 35–38 and 64 kd (not shown). These band sizes are consistent with those seen in lung of sheep and man.5,6 Similar findings were present in lung homogenates of calves similarly inoculated with P. aeruginosa, killed P. aeruginosa, and P. aeruginosa LPS (not shown).
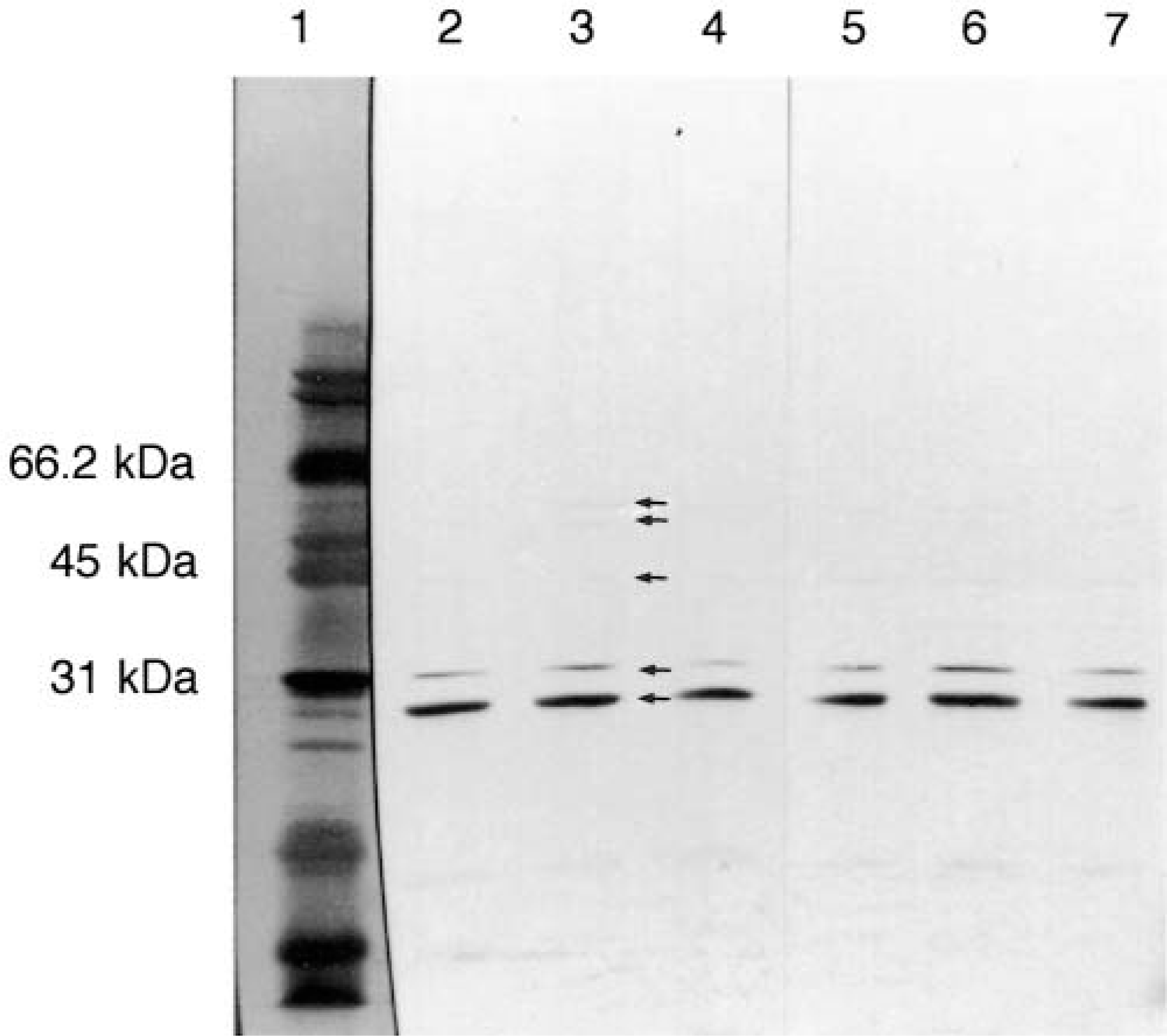
The western blots of homogenized lung tissue from neonatal calves inoculated intrabronchially with Mannheimia haemolytica (lanes 2, 4, 6, 7) or saline (lanes 3 and 5) and then euthanatized 6 hours after inoculation. Proteins were separated on a 12.5% SDS-PAGE by electrophoresis and transferred to PVDF membranes. A monoclonal antibody (IG9-IC2) against AP is immunoreactive to several fragments of different sizes that are also seen in the western blots of sheep and man. The 31.0-kd band is most consistently immunoreactive. Broad-range molecular weight markers are shown in lane 1. Note the similarities in band size despite the different treatments, indicating a lack of induced expression.
Distribution of AP-IR
Control lung (PFS-inoculated animals)
Lung tissue from control (PFS-inoculated), neonatal, and growing (3-month-old) calves lacked inflammatory cells or other lesions. The distribution of AP-IR was similar between neonatal and growing calves, and it was similar to that of human and ovine lung stained for AP-IR. In bronchi, intense staining in the nuclei of multifocal bronchial epithelial cells was present in all age groups. AP-IR was also present in the apical cytoplasm of most cells in the pseudostratified epithelium of bronchial mucosa. In most areas of bronchial epithelium, there was intense staining (nuclear, cytoplasmic, or both) of multifocal, individual bronchial epithelial cells, whereas adjacent cells completely lacked staining. Nuclei of multifocal cells of the bronchial submucosal glands were immunoreactive for AP, but submucosal gland cells lacked cytoplasmic AP-IR.
In bronchioles of neonatal and 3-month-old calves, AP-IR was present in nuclei of multifocal cells and in the apical cytoplasm of multifocal epithelial cells (Fig. 2). AP-IR was present in the nucleus and was only sporadically present in the cytoplasm of alveolar epithelial cells. In all age groups, nuclear AP-IR of arterial and bronchial smooth muscle cells was present, and cytoplasmic AP-IR was rare. AP-IR was distributed randomly in the smooth muscle cells, and it did not have a consistent or discernable pattern of staining within the layers of the bronchi or in the vascular wall.
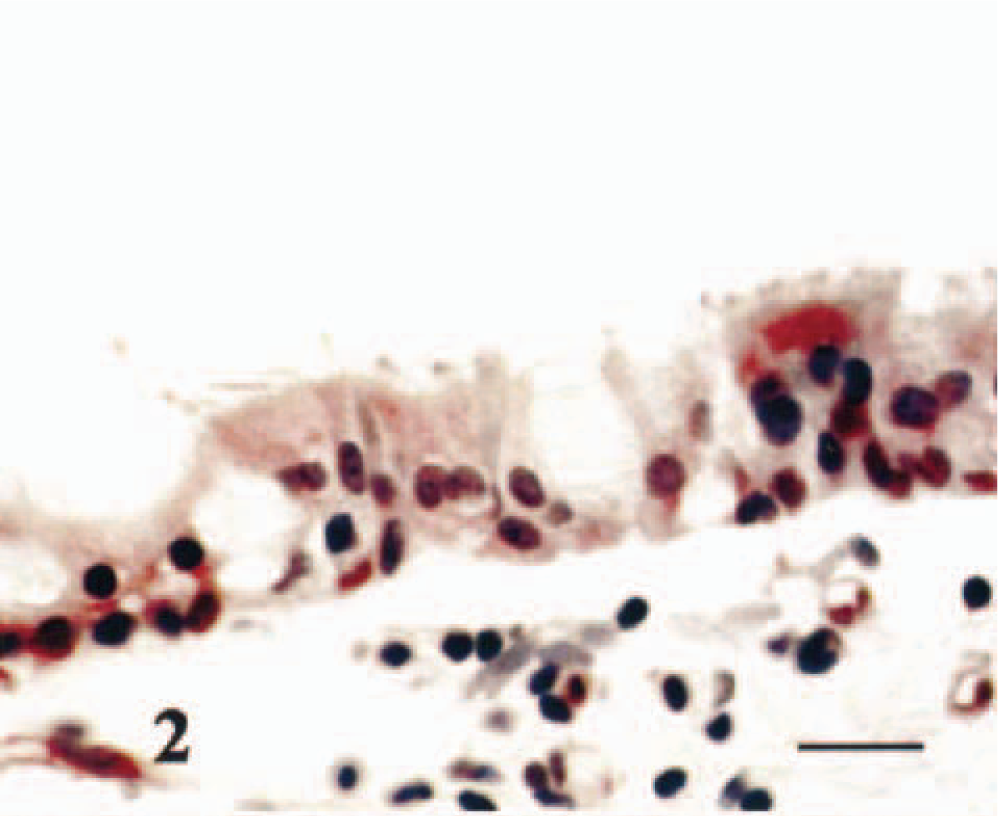
Control lung lacking inflammation from calf inoculated with PFS 6 hours after inoculation. Multifocal bronchiolar epithelial cells have relatively increased cytoplasmic and/or nuclear immunoreactivity for AP. Hematoxylin counterstain. Bar = 30 μm.
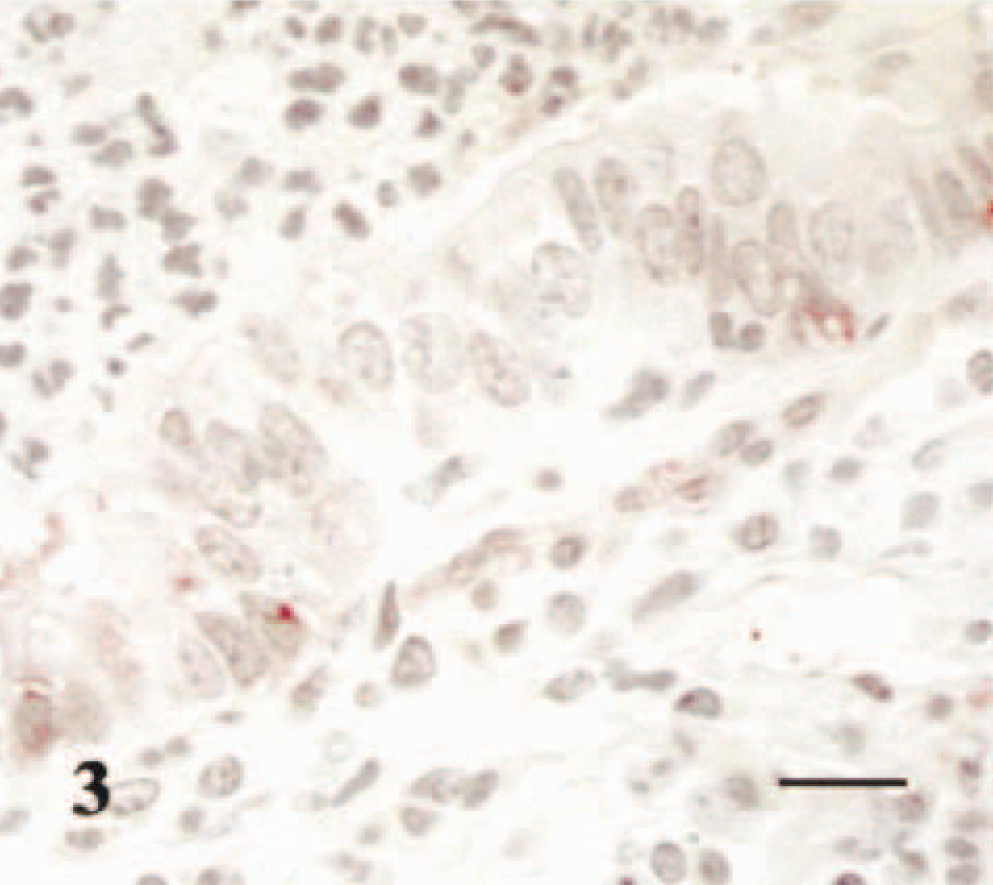
Lung from calf infected with M. haemolytica 6 hours after inoculation. The bronchiolar lumen contains numerous neutrophils, and in the epithelium there is minimal/mild immunoreactivity for AP (red) in multifocal nuclei and, to a lesser degree, the cytoplasm of bronchiolar epithelial cells. Neutrophils and exudate lack immunoreactivity. Hematoxylin counterstain. Bar = 25 μm.
Inflamed lung (M. haemolytica, LPS, and LPS-P–inoculated animals)
No inflammatory infiltrates were present in the tissues of control (PFS-inoculated) animals (Tables 1, 2). Inflammatory lesions consistent with M. haemolytica pneumonia were present in animals that received M. haemolytica and M. haemolytica LPS. Only minimal infiltrates of neutrophils were present in animals inoculated with M. haemoltyica LPS-P.
Inoculation with M. haemolytica, LPS, or LPS-P did not alter AP-IR distribution in bronchi of neonatal and growing (3-month-old) calves. In bronchioles of inoculated neonatal and growing calves, the distribution of nuclear and cytoplasmic staining was decreased (fewer cells stained, Fig. 3) when compared with bronchioles containing inflammatory cells (Tables 1, 2), but the decrease was not statistically significant. Lungs inoculated with M. haemolytica and LPS had the highest degree of inflammatory exudate, yet fibrinous exudate and neutrophils in the lumen lacked AP-IR. Nuclei of multifocal lymphocytes within the lamina propria and adventitia of bronchi were immunoreactive for AP, but neutrophils within the bronchial lumina lacked AP-IR. AP-IR was not reduced in alveoli that contained inflammatory exudate.
Intensity of inflammation and distribution and intensity of AP-IR in lungs of growing (3 months old) cattle 24 hours after intrabronchial inoculation of PFS, M. haemolytica LPS-P, M. haemolytica LPS, and M. (Pasteurella) haemolytica.
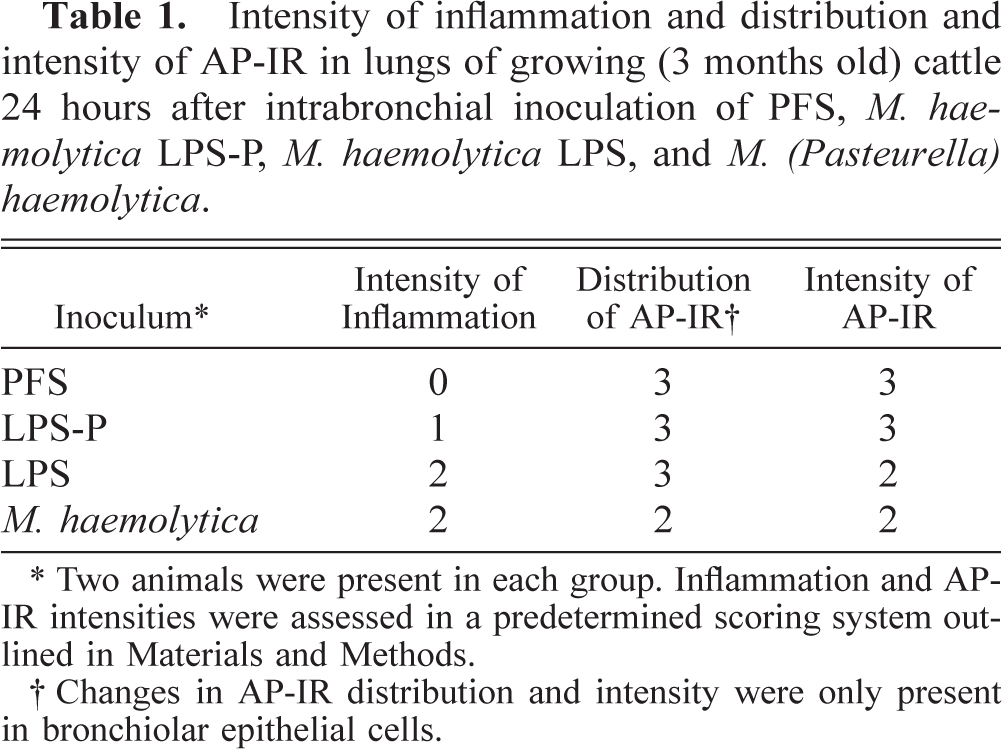
∗ Two animals were present in each group. Inflammation and APIR intensities were assessed in a predetermined scoring system outlined in Materials and Methods.
† Changes in AP-IR distribution and intensity were only present in bronchiolar epithelial cells.
Intensity of inflammation and distribution and intensity of AP-IR in lungs of neonatal calves inoculated with either PFS or M. haemolytica.

∗ Four animals were present in each group. Inflammation and APIR intensities were assessed in a predetermined scoring system described in Materials and Methods.
† Changes in AP-IR distribution and intensity were only present in bronchiolar epithelial cells.
Intensity of AP-IR in control and inflamed lung
AP-IR intensity was not changed in bronchi and alveoli but decreased slightly in bronchioles of both neonatal and growing animals with evidence of inflammatory infiltrates when compared with control (PFS-inoculated) animals (Tables 1, 2; Fig. 3). This decrease, however, was also not statistically significant. The degree of AP-IR was similar between animals in each group.
Discussion
In this study, AP-IR was widespread in the lungs of neonatal and growing (3-month-old) calves in both control and acutely inflamed tissue. Also, the distribution and the western blots of AP-IR in lung tissue of cattle were similar to those of previously described lung in human and sheep.5,6 But the concentration of antibody required to detect AP-IR by immunohistochemistry in the sheep lung is the lowest of the three species. This may be because of the greater avidity of the antibody to ovine AP, the availability of AP in ovine tissue sections, or the amount of AP in ovine tissues. The antibody may have higher avidity for ovine AP because the exact structure of the possible precursor molecules in cattle, sheep, and humans is currently unknown (see discussion below). Respiratory epithelial cells of all three species produce APs, indicating that APs are present in a location where antimicrobial activity would be most effective against microbial colonization.
AP-IR distribution and intensity were not increased, and, in contrast, appeared decreased in the nuclei and cytoplasm of bronchiolar epithelial cells of neonatal and growing calves inoculated with M. haemolytica and growing calves inoculated with M. haemolytica LPS. M. haemolytica LPS-P incited only mild inflammatory infiltrates and also lacked changes in AP-IR distribution and intensity as compared with controls. The inability of AP-IR to increase in this study indicates that APs are not induced by live bacteria, bacterial LPS, or the acute inflammatory response. This is in sharp contrast to bovine LAP, a β-defensin, which is markedly increased in the same species (bovine) and breed (Holstein) with the same inoculum (M. haemolytica) in the same laboratory.23 This suggests that APs are constitutively produced and rapidly expended during acute pulmonary inflammation. AP is present in high concentrations in BAL fluid of humans,4 sheep,5 and cattle8 and may be an important antimicrobial factor during the initial stages of microbial colonization and infection. The rapid availability of AP may allow time for other inducible antimicrobial peptides, such as HBD-2 in man, LAP and TAP in the bovine, to be transcribed and translated. Although some cationic antimicrobial peptides may inhibit/dampen the inflammatory response,14,15 it is possible that AP, with its anionic charge and small size, may not affect this process and may therefore allow development of the inflammatory response during the earliest stages of microbial colonization and infection. Some recent work suggests that AP expression is dependent on the state of epithelial cell differentiation. During the more chronic (repair) stages of bacterial pneumonia, epithelial cells undergoing hyperplasia have increased AP in comparison with well-differentiated, mature cells.11 An increase of AP in hyperplastic cells during chronic pneumonia may be due to increased basal expression, increased cytoplasmic storage, or a lack or release/secretion of APs.11
The western blots suggest that APs (as intact zymogens) were present in the neonatal and growing animals as well as in adult animals. Several types of proteins, including nucleosomal proteins, contain polyaspartate regions that may be detected by the IG9-IC2 monoclonal antibody. For example, nonhistone acidic proteins that bind to chromatin, known as the high-mobility group (HMG) proteins, are present in the nucleus. These proteins have long stretches of acidic amino acid sequences, with some stretches of up to 40 aspartic acid and glutamic acid residues.24 HMG and other types of proteins may release APs as a normal part of cellular metabolism or maturity; but the exact protein or proteins that release forms of AP that have antimicrobial activity are not known, nor are the cleavage processes that liberate active APs. Additional work with these proteins is necessary to determine more precisely which protein(s) releases APs with antimicrobial activity. Despite this, the study demonstrates that cattle lung epithelia express APs in a similar distribution to sheep and man.5,6 The work demonstrates that a subset of and not all the respiratory epithelial cells express APs and that expression is not increased by inflammatory stimuli such as live bacteria, LPS, or an LPS-P. Therefore, there are precise regions of AP production within the lung, and the production is likely constitutive.
Footnotes
Acknowledgements
This work was supported in part by a USDA/CSREES/NRI/CGP grant 99-35204-7681, the J. G. Salsbury Endowed Funds, and the Healthy Livestock Initiative (HLI). We thank Keith DeJong for his assistance with the western blots.