
Abstract
A total of 100 liver samples from pigs with postweaning multisystemic wasting syndrome (PMWS) were studied. All livers were evaluated microscopically and were staged based on the severity and localization of lesions. Presence of porcine circovirus type 2 (PCV-2) was evaluated using an in situ hybridization technique. Eighty-eight of 100 livers (88%) had a variable degree of lymphohistiocytic hepatitis, with apoptotic bodies, disorganization of hepatic plates, and/or perilobular fibrosis. Twelve pigs did not have microscopic liver lesions. Four stages of hepatic damage were established based on intensity and distribution of the lesions. Viral nucleic acid was detected in 70 of 100 livers (70%). Target cells for PCV-2 infection included Kupffer cells, hepatocytes, and inflammatory infiltrates. According to distribution of PCV-2 nucleic acid, four basic labeling patterns were identified. This study shows that liver damage is a frequent microscopic finding in cases of PMWS and hepatocytes are a target cell for PCV-2 infection and replication. Therefore, PCV-2 should be considered a new hepatitis-inducing viral agent in pigs.
Postweaning multisystemic wasting syndrome (PMWS) is a new porcine disease that affects nursery and early fattening pigs. Accumulated evidence, using in situ hybridization (ISH), immunohistochemistry, virus isolation, or detection by polymerase chain reaction methods has shown the presence of porcine circovirus type 2 (PCV-2) in tissues of pigs with PMWS. 2 5 6 8 Furthermore, experimental inoculation of pigs with PCV-2 or with tissue homogenates from pigs with PMWS produced a mild form of the disease. 1 3 Although macroscopic lesions can suggest the disease, diagnostic confirmation is based on histopathologic examination and detection of virus. 6 Main lesions consist of lymphocyte depletion and histiocytic inflammatory reaction in lymphoid tissues and variable degrees of interstitial pneumonia. Other microscopic lesions, which probably develop in later stages of the disease, are interstitial nephritis and hepatitis. 8 Hepatic lesions have been described as lymphocytic-histiocytic inflammatory infiltration in portal zones, single cell necrosis of hepatocytes, swelling and vacuolation of hepatocyte cytoplasm, and karyomegaly. 6
The purpose of this study was to describe the frequency and the stages of hepatic damage observed in pigs naturally infected with PCV-2 and to describe the distribution of viral nucleic acid in the hepatic parenchyma.
A total of 100 liver samples from 6–16-week-old pigs with PMWS from 52 farms in central and northeastern Spain were included in this study. These samples corresponded to all PMWS cases diagnosed in pigs submitted to the Department of Pathology of the Veterinary School of Barcelona (Spain) from May 1997 to June 1998. Sixty pigs were euthanatized and necropsied immediately. The other 40 pigs were represented by formalin-fixed tissues submitted by swine practitioners. The final diagnosis of PMWS was established in all animals by the presence of characteristic microscopic lesions and PCV-2 nucleic acid in tissues. 8 Fixed samples were dehydrated, embedded in paraffin, sectioned at 4 µm, and stained with hematoxylin and eosin (HE). The ISH technique was carried out on liver samples from diseased pigs following previously described protocols. 8 A specific 40-nucleotide probe (5′-CAGTAAATACGACCAGGACTACAATATCCGTGTAACCATG-3′) complementary to nucleotides 1,085–1,124 of open reading frame 2 from PCV-2 was used; 5 this probe only labeled PCV-2-infected cells when tested for specificity on cell cultures infected with PCV-1 and PCV-2.
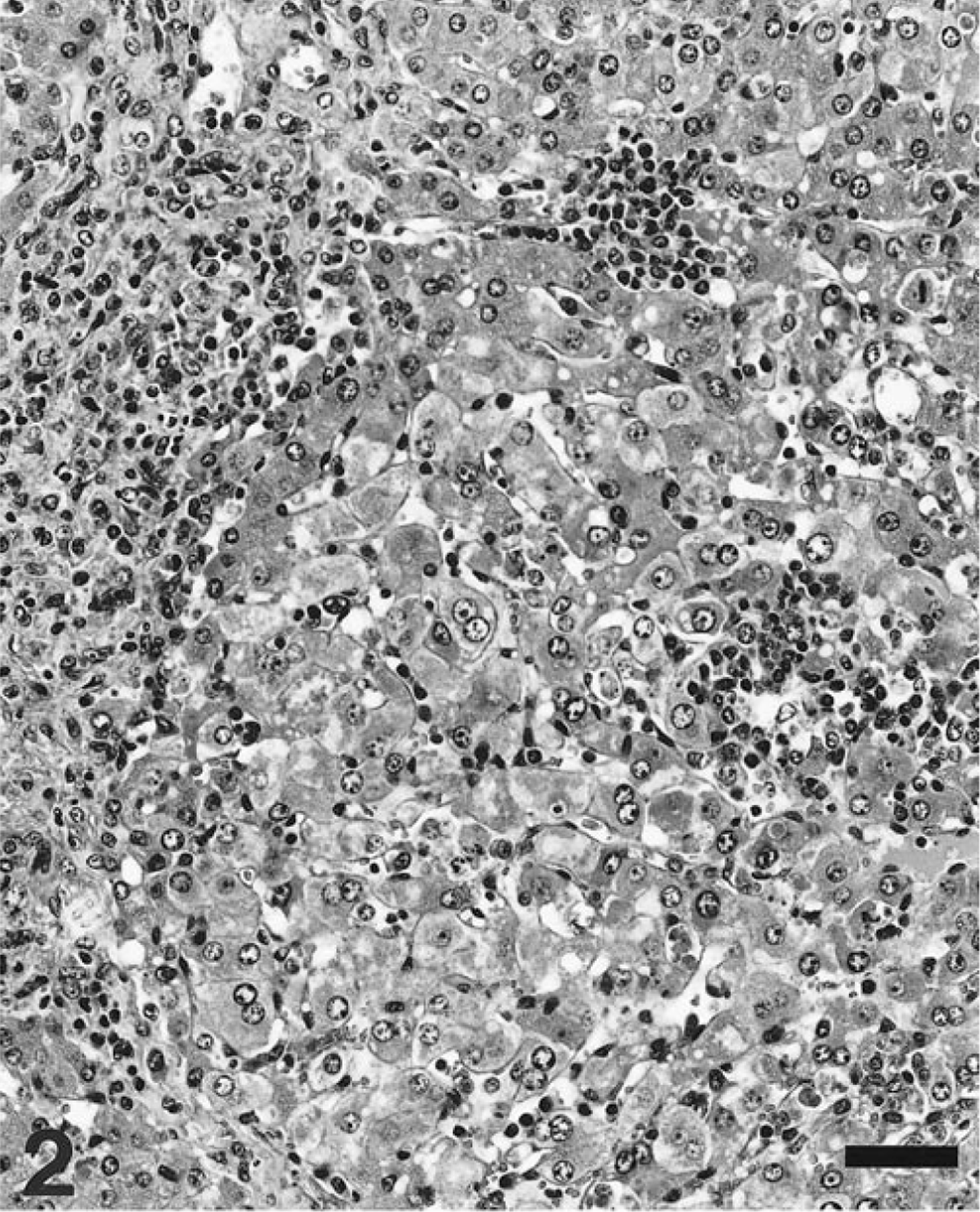
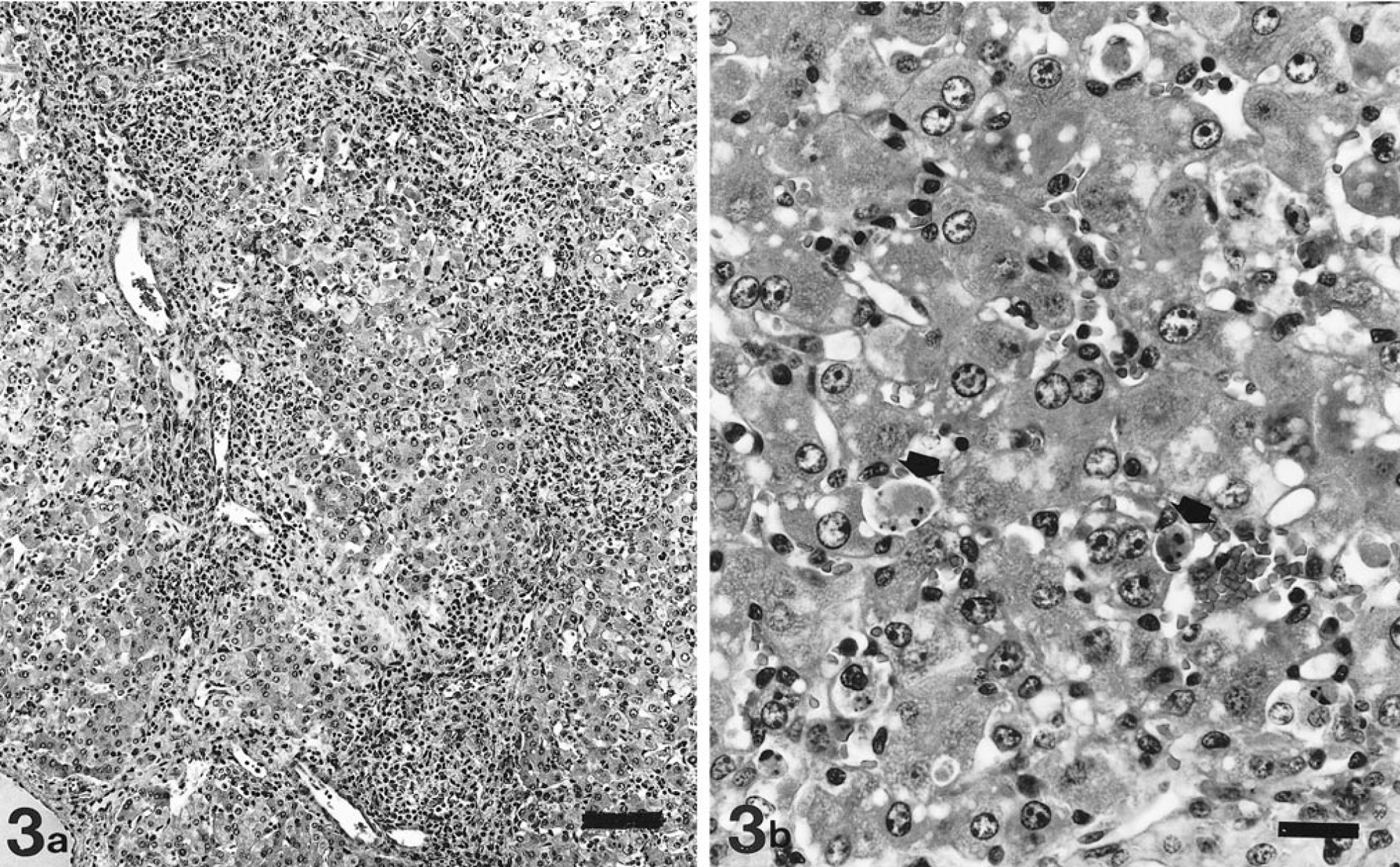
Grossly, 51 livers of the 60 necropsied pigs had no remarkable lesions. Of the other nine livers, seven had an orange-yellow discoloration (icterus), one was apparently reduced in size, and one was increased in size (hepatomegaly). In constrast, microscopically, most (88 of 100 pigs) had a variable degree of lymphohistiocytic inflammatory infiltration of hepatic parenchyma, together with formation of apoptotic bodies (Councilman bodies), disorganization of hepatic plates, and perilobular fibrosis. Twelve pigs did not have microscopic liver lesions. Four stages of hepatic damage (Table 1) were established, based on intensity and distribution of lesions. Livers with slight multifocal lymphohistiocytic infiltrates in the portal tracts (47 of 100 livers examined) were classified as stage I (Fig. 1). In stage II (26 of 100 livers), intermediate to intense lymphohistiocytic inflammation of the portal tracts was often accompanied by clusters of mononuclear inflammatory cells in the liver parenchyma (Fig. 2). Prominent, enlarged Kupffer cells were found lining the sinusoidal walls. Infrequently, apoptotic bodies were observed within the parenchymal inflammatory foci. The distinctive lesions of stage III were multifocal clusters of mononuclear inflammatory infiltrates within the hepatic parenchyma, a variable hepatic plate disorganization (Fig. 3a), and frequent presence of apoptotic bodies (Fig. 3b). Concilman bodies were present in almost all hepatic lobules. Also, a severe generalized lymphohistiocytic inflammation in the portal tracts was invariably associated with this stage of liver damage. In about half of these cases, enlarged hepatocytes with karyomegaly were prominent. Also, some enlarged hepatocyte nuclei had chromatin condensation along the nuclear membrane. Stage III lesions were detected in 10 of 100 livers. The most severe liver damage (stage IV), observed in 5 of the examined livers, was a generalized perilobular fibrosis, with disorganization of liver plates and massive loss of hepatocytes (Fig. 4). The remaining stroma of the lobules was diffusely infiltrated by mononuclear cells. The seven livers showing orange-yellow discoloration were classified in lesional stage II (two livers), stage III (four livers), and stage IV (one liver). The livers that were smaller and larger than normal were classified histologically as stage IV.
Results of PCV-2 in situ hybridization (ISH) in relationship to histopathologic changes in 100 pigs with PMWS.

Amount of PCV-2 nucleic acid: - = absence; + = slight; + + = moderate; + + + = intense.

Liver, stage I; pig. Slight presence of mononuclear inflammatory infiltration in the perilobular areas (arrows). HE. Bar = 120 µm.
Liver stage II; pig. Mononuclear inflammatory infiltration of the perilobular areas and a cluster of inflammation of the periacinar area. HE. Bar = 60 µm.
Liver stage III; pig.
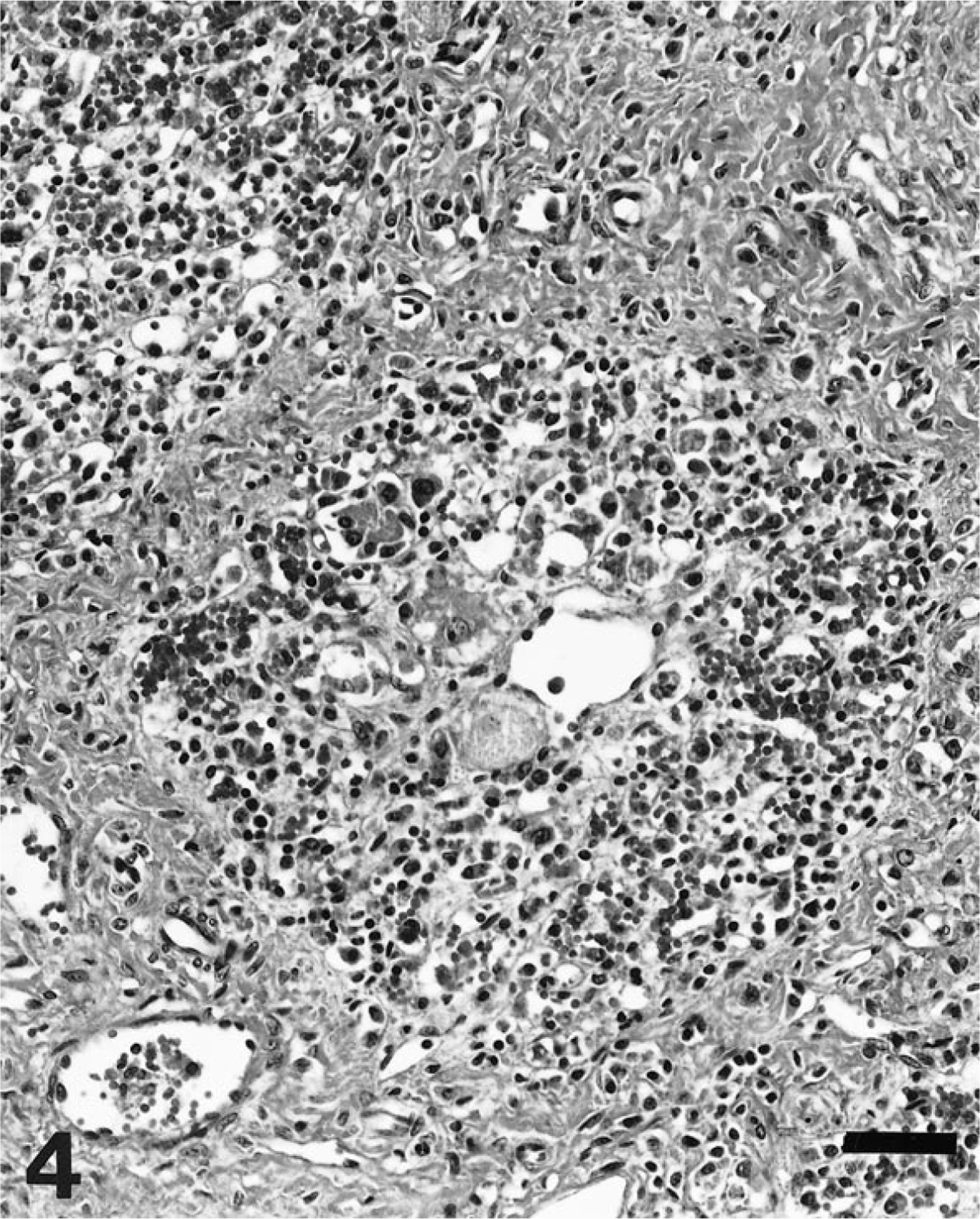
Liver; stage IV; pig. Marked perilobular fibrosis, with disorganization of liver plates, and massive loss of hepatocytes. The stroma of the lobules is diffusely infiltrated by mononuclear cells. HE. Bar = 60 µm.
Results of PCV-2 nucleic acid detection and its correlation to the stage of liver damage are summarized in Table 1. Viral nucleic acid was detected in 70 of 100 livers (70%); of those, 63 had some degree of hepatic damage. Target cells for PCV-2 infection included Kupffer cells, hepatocytes, and inflammatory infiltrates. Labeling in Kupffer and mononuclear inflammatory cells was mainly located in the cytoplasm and, to a lesser extent, in the nuclei. In contrast, PCV-2 nucleic acid in hepatocytes was found most frequently in the nucleus and sporadically in the cytoplasm (Fig. 5). Occasionally, viral nucleic acid was found in circulating mononuclear leukocytes within the hepatic sinusoids. Fine projections in the sinusoidal wall lining, which probably corresponded to endothelial sinusoidal cells, were occasionally labeled. Neither biliary ducts nor portal venous or arterial structures were stained. According to distribution of PCV-2 nucleic acid, four basic labeling patterns were described: Kupffer cell, periportal, mixed (Kupffer cell and periportal), and diffuse. Frequency of each pattern is given in Table 1. The pattern of Kupffer cell staining corresponded to livers that were histologically normal or in stages I and II. In this pattern, a variable number of Kupffer cells throughout the whole lobule stained positively in cytoplasm and/or nuclei (Fig. 6). The periportal pattern was characterized by intense labeling of the inflammatory infiltrates in the portal tract and in the adjacent hepatic parenchyma. Occasionally, nuclei of hepatocytes in the vicinity of inflammatory infiltrates were intensely stained. Pure periportal pattern was found in only one liver, in stage I. A mixed pattern (Fig. 7) including both Kupffer cell and periportal staining was detected in two livers in stages I and II. Extension of PCV-2 infection to hepatocytes and inflammatory and Kupffer cells led to a diffuse distribution of viral nucleic acid in the hepatic parenchyma (Fig. 8). The diffuse pattern of labeling was associated (in decreasing frequency) with stages IV, III, and II.
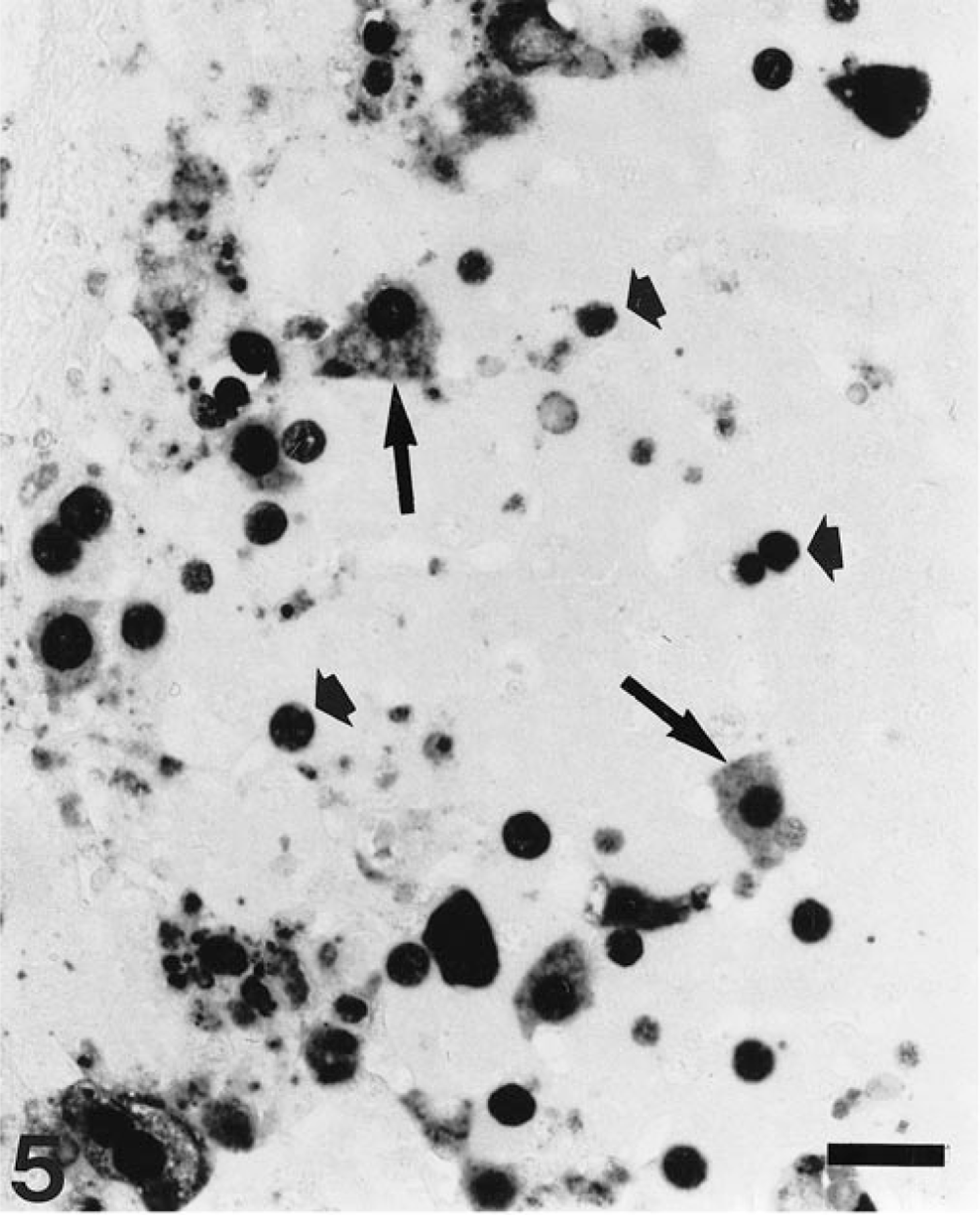
Liver; pig. Presence of PCV-2 nucleic acid mainly in the nucleus of hepatocytes (short arrows) and sporadically in the cytoplasm (long arrows). ISH, fast green counterstain. Bar = 30 µm.
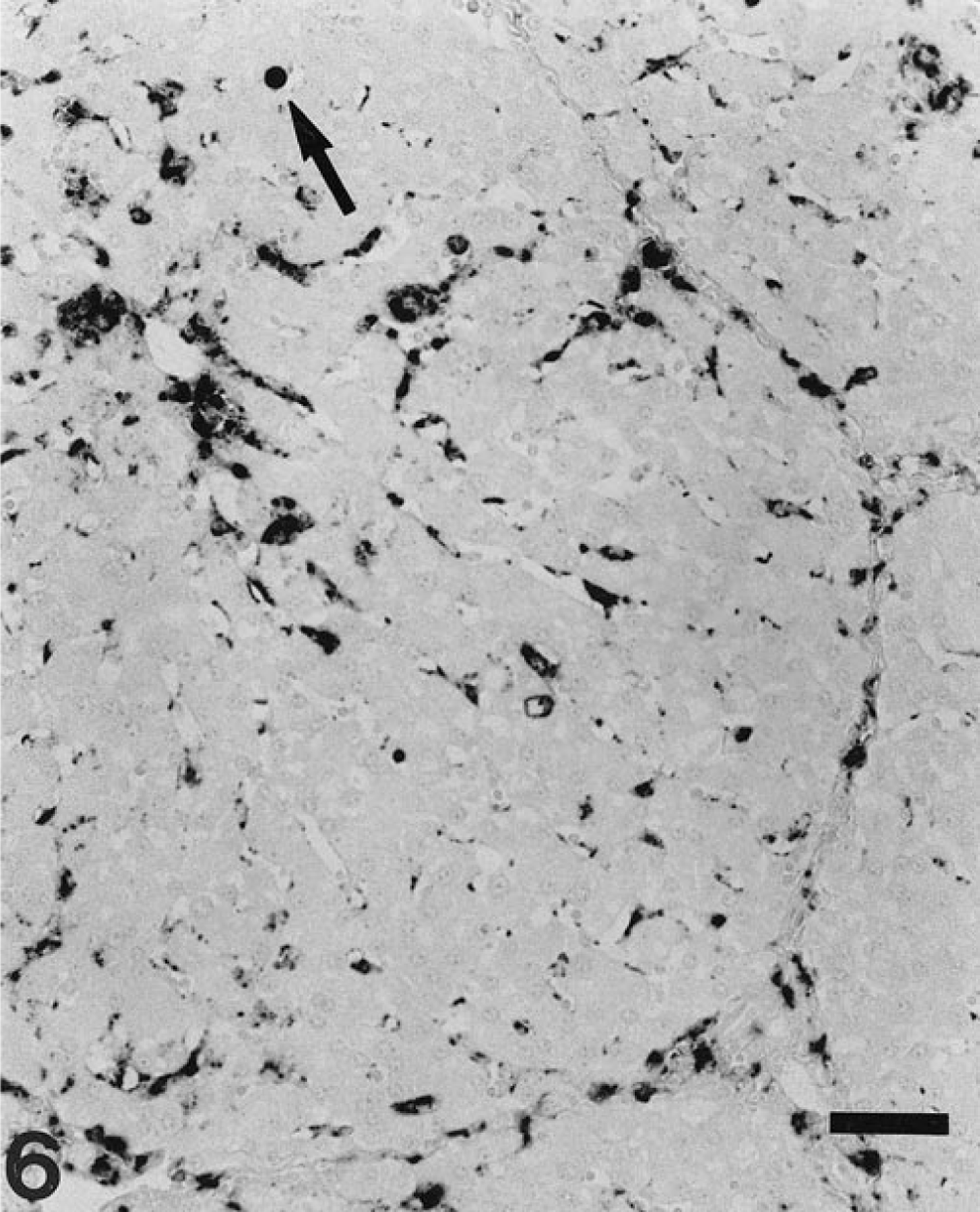
Liver; pig. Kupffer cell pattern of PCV-2 genome distribution. Kupffer cells throughout the lobule appear stained. Occasionally, nuclei of hepatocytes are stained (arrow). ISH, fast green counterstain. Bar = 60 µm.
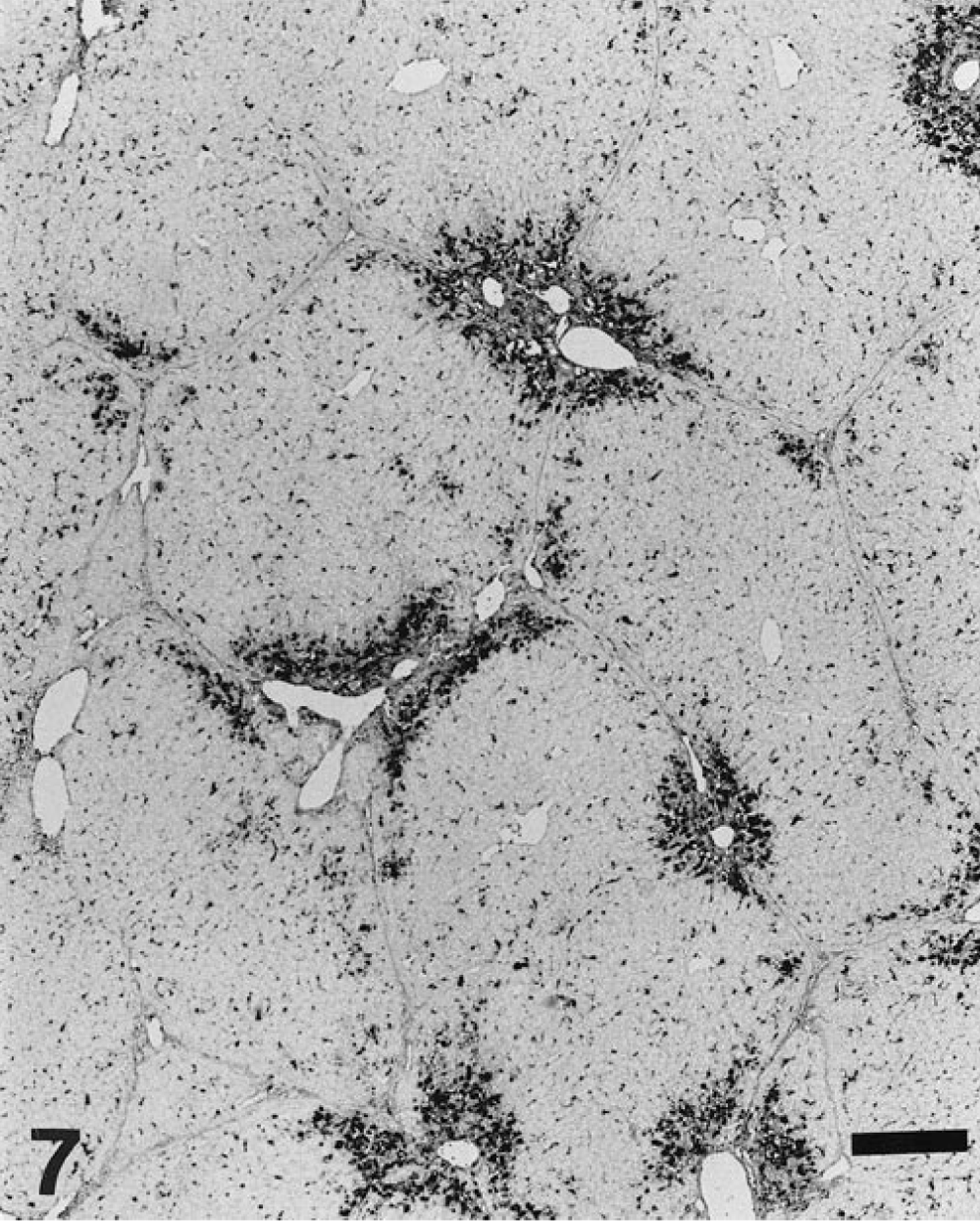
Liver; pig. Mixed pattern of viral distribution includes both Kupffer cell and periportal staining. The periportal pattern shows intense labeling of the inflammatory infiltrates located in the portal tract and in the adjacent hepatic parenchyma. ISH, fast green counterstain. Bar = 300 µm.
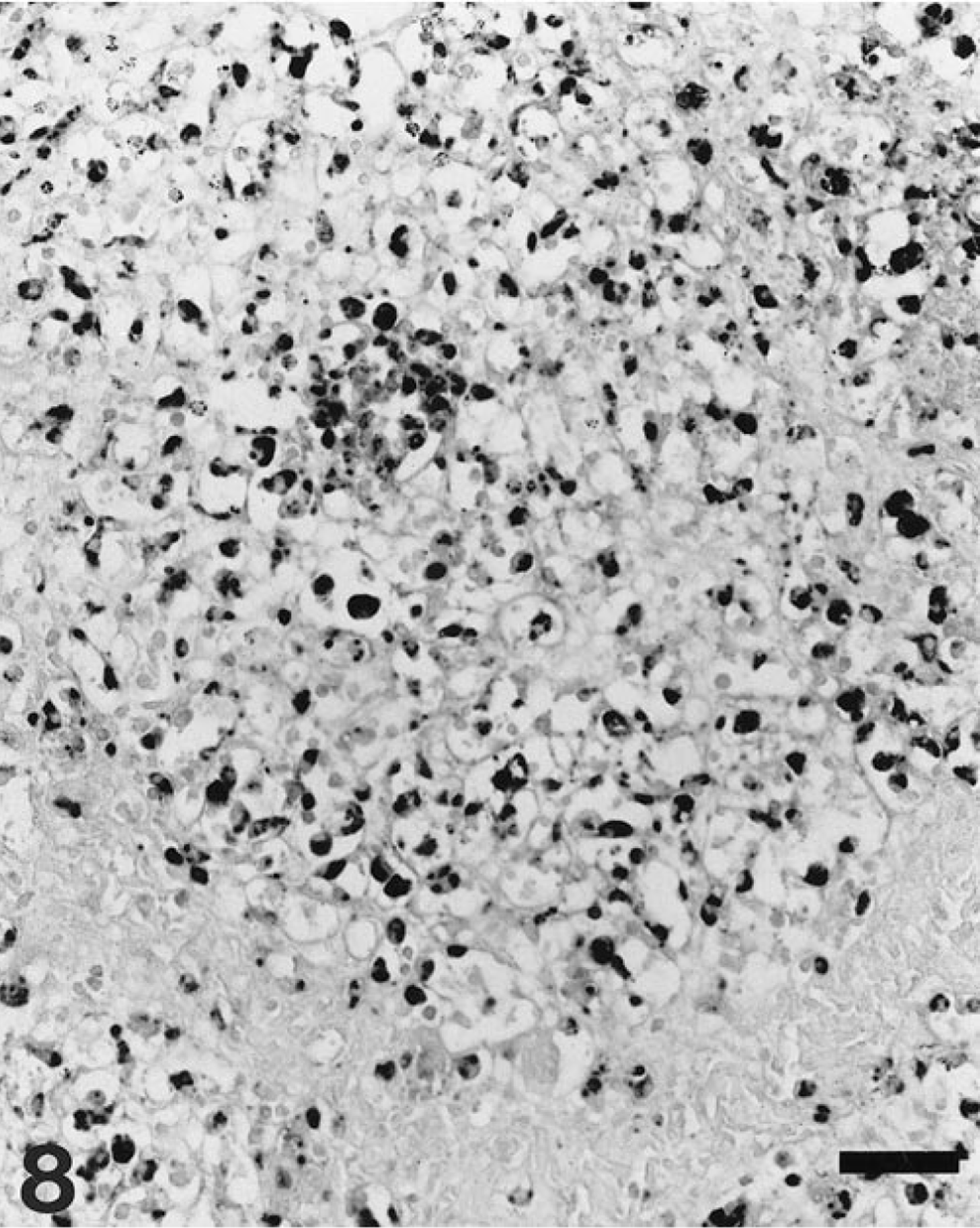
Liver; pig. Diffuse distribution pattern of viral nucleic acid in the hepatic parenchyma, which corresponds to a stage IV lesional pattern. ISH, fast green counterstain. Bar = 50 µm.
In this study, liver damage was a frequent microscopic finding in cases of PMWS. The nature of the lesions was used to classify liver damage into four stages, representing increasing intensity and extension of inflammatory and cytologic changes. Most pigs had slight to moderate inflammatory lesions (stages I and II), which probably remained subclinical during the course of the disease. The most advanced liver damage (stage IV) was sporadically found in pigs with PMWS and was characterized by massive loss of hepatocytes and perilobular fibrosis. The severity of these histopathologic changes suggests that liver failure may occur terminally in PMWS. These findings also demonstrate that the hepatocyte is a target cell for PCV-2 infection and replication, and therefore PCV-2 should be considered a new hepatitis-inducing viral agent in pigs.
The stages of the lesion described in this study probably fit the temporal sequence of pathologic events that occur in the liver of PMWS-affected pigs. Initial liver damage probably starts with a widespread infection of Kupffer cells by PCV-2. Susceptibility of Kupffer cells and of other cells of the monocyte–macrophage lineage to PCV-2 infection has been previously reported. 2 8 Inflammatory infiltration by mononuclear cells in periportal areas (stage I) was found in livers with Kupffer-cell pattern of PCV-2 distribution but also in livers negative for PCV-2 by ISH. Therefore, no relationship between presence of periportal infiltrates and PCV-2 Kupffer cell infection was established.
Periportal and mixed patterns of PCV-2 distribution are probably transient forms and were observed at a very low frequency. Few PCV-2–infected hepatocytes appeared in livers classified as stage II, but the generalization of PCV-2 infection to these cells is probably the hallmark of liver-induced damage by the virus, characterized by stage III lesions. The extension of viral infection from contiguous cells (e.g., endothelial, Kupffer, and inflammatory cells) may be the mechanism by which hepatocytes become infected; however, other factors such as the cell cycle phase may be relevant. This suggestion is supported by the observation that in the few livers with periportal infiltrates, infected hepatocytes were mainly found in the vicinity of periportal areas. However, it is also known that mitosis is needed for PCV-1 replication, 9 and mitotic activity in liver is largely confined to periportal hepatocytes. 7 Infection of hepatocytes by PCV-2 is qualitatively different from the infection of cells of the monocyte–macrophage lineage in that viral nucleic acid is mainly located in the nucleus of the hepatocyte, whereas the cytoplasm is rarely labeled.
Extensive infection of hepatocytes led to disruption of hepatic plates and to changes in the morphology of these cells, including megalocytosis with karyomegaly and apoptotic alterations. These changes have been observed in other pigs with PMWS. 4 Livers showing these morphologic changes in hepatocytes had a diffuse pattern of PCV-2 infection, suggesting that PCV-2 may be the cause of them. Histopathologic evidence implies that PCV-2 and apoptosis are probably associated with the death of hepatocytes. Extensive loss of hepatocytes, diffuse inflammatory infiltration, and perilobular fibrosis occurred as a final stage of liver damage. Macroscopically, these livers appeared atrophic and there was no histologic evidence of hepatic regeneration. Proliferation of biliary ducts was not an obvious feature, even in stages III and IV.
Severe liver damage seems to be a late event in the pathogenesis of PMWS. In this study, only 15 of 100 pigs developed stage III or IV lesions. Possibly, the other pigs did not survive long enough to develop severe hepatic damage, but other factors in the development of liver lesions in PMWS-affected pigs cannot be ruled out.
Footnotes
Acknowledgements
We thank B. Perez, S. Usero, and P. Losada for technical assistance. We also thank R. Cory for English correction of the manuscript. This work was partly funded by Project QLRT-PL-199900307 from the Fifth Framework Programme 1998–2002 of the European Commission and project 2-FEDER-1997-1341 from the I+D National Plan (Spain).