
Abstract
The tail cut bleeding model (CUT) is routinely used in factor VIII-deficient mice to assess pharmacodynamic effects of therapeutic strategies for haemophilia A. Results from this model are highly variable, many modifications to the model are reported and at times the animals’ wellbeing may be compromised by recording survival as an endpoint. We therefore investigated if the ferric chloride carotid occlusion model (COM) used for thrombosis research can be applied to enhance data quality and animal welfare in haemophilia A research. Relative dose effects and relative dose variations were calculated for the CUT and COM. The requisite sample sizes were estimated and the importance of survival rates to assess rebleeds during recovery was evaluated by correlating initial blood loss to mortality. Relative dose effects increased with higher doses in both models. The COM was more sensitive at lower doses than the CUT, had up to 82% less variation across doses and clearly showed superior accuracy. Only 5% of the sample size required for the CUT would be needed to establish non-inferiority between a specific therapeutic dose in haemophilia A mice and healthy wild-type animals. A strong statistically significant correlation was found between initial blood loss and mortality within 24 h. Our findings clearly suggest that the COM is a valid tool for assessing haemophilia A treatment
Haemophilia A is an inherited X-linked clotting disorder caused by a deficiency of plasma factor VIII (FVIII) that affects approximately one out of 5000–10,000 men and is characterized by an abnormal bleeding tendency. 1 Intravenous injections of FVIII concentrates purified from human plasma or produced by recombinant technology (rFVIII) are effective in controlling bleeding. Current research is exploring the development of molecules with increased potencies, modified rFVIII with a prolonged half-life to reduce the frequency of prophylactic intravenous injections and gene transfer therapy to provide an alternative long-term correction of FVIII deficiency.
Several different animal models are used to mimic impaired haemostasis in patients. These include naturally occurring bleeding disorders in dogs, models that use pharmacologically induced conditions in rabbits, rats and mice, and targeted genetic modification in mice. All these models have been helpful in developing treatment strategies against bleeding disorders. In principle, haemorrhage is induced and the inability of the animal to form a stable clot within a given time with and without treatment is assessed and compared with physiologically normal animals. A cuticle bleeding time and a gingival bleeding time have been used in dogs for this purpose.2-4 Large animal models closely resemble haemophilia in humans with dosing of replacement and bypassing therapies more similar than in small animal models to that of humans.5-7 Nonetheless, as genetically modified mice are available they are most frequently used to evaluate safety and efficacy. Maintaining a haemophilic dog colony is labour-intensive and expensive. In addition, the short reproduction cycle of mice renders larger quantities of experimental animals.
Two FVIII knock-out mouse models were developed for research of haemophilia A through disruptive insertion of a neomycin gene into either the exon 16 (E16) or 17 (E17) of the murine FVIII gene. Both models have a functional FVIII activity of less than 1% in their plasma, which makes their plasma comparable to plasma of humans with haemophilia A. 8
Although FVIII activity is virtually absent in these mice, different results of spontaneous bleeding events are reported, which makes it difficult to characterize their phenotype. Lethal spontaneous bleedings in 10.0% of females and 19.8% of males have been reported, 9 but no such events are recorded in other reports.8,10 Amputation of the tail tip is the most common trauma inflicted to assess the impaired coagulation function in mice. However, variation between reports on how much of the tail is clipped, how the blood is collected, or bleeding time and survival rate are recorded is considerable. The effort to overcome the difficulty of producing reproducible bleedings might in part account for the many modifications of the model that have been developed. The tail clip bleeding time is certainly abnormal in haemophilic mice and the bleeding time and volume bled are highly variable. 5 Conflictingly it was reported that tail vein bleeding time is not affected by FVIII deficiency. 11 We therefore speculated that statistically significant results would be unlikely in the case of small effects with conventional sample sizes.
In contrast to murine haemophilia models, a standardized murine model of vascular thrombosis is available. Ferric chloride (FeCl3) is commonly used as a thrombogenic agent for investigating the mechanisms of thrombosis in transgenic mice. 12 FeCl3 applied topically to the carotid artery of mice has been used to study the effects of plasminogen activator inhibitor-1 deficiency on arterial thrombolysis. 13 FeCl3 in this case proved useful for studying initial thrombus formation as well as the factors involved in clot lysis. 14 A direct relation between FeCl3 concentration and vascular occlusion has been demonstrated in a refined murine model. 15 Direct application of FeCl3 to the adventitial surface of an artery rapidly induces formation of a platelet-rich thrombus that typically progresses to complete vascular occlusion in wild-type animals. 13 But, no thrombus forms with this model in haemophilia A and haemophilia B mice. 16 Thus, we supposed that this model would be useful for assessing the ability of therapeutic interventions to restore haemostasis in haemophilic animals.
We hypothesized that using the FeCl3 carotid occlusion model (COM) in FVIII knock-out mice would facilitate a simple, sensitive and highly standardized model to test potential therapeutic agents or gene therapy for haemophilia A
Materials and Methods
All animal procedures carried out for this study were subject to review and were approved by the Baxter BioSience Institutional Animal Care and Use Committee and were in compliance with the Austrian Ordinance on Animal Experiments.
Animals and husbandry
We used 160 female and male locally bred Exon 17-disrupted haemophilic A mice weighing between 20 and 25 g (6–8 weeks old). As the interest of the study was comparing methods, equal numbers of females and males were used for each model. The colony of fully inbred haemophilic A mice was established with a breeding pair from the original colony (on a 129SV and C57BL/6 mixed background) and crossed on to the C57BL/6J background.
9
Twenty-two female and male C57BL/6J01aHsd (C57BL/6J) mice within the same weight range were purchased from Harlan (Harlan Winkelmann GmbH, Borchen, Germany). C57BL/6J mice delivered by van in filter boxes were acclimatized for seven days before use as control animals. Mice were separated by sex and conventionally housed in groups of 2–3 in polycarbonate Eurostandard type II cages (Tecniplast, Buguggiate, Italy) with aspen wood chips as bedding and aspen wood shavings as nesting material (Abedd, Vienna, Austria). Mice were fed a pelleted maintenance diet (R/M-H, ssniff, Soest, Germany) and received tap water via polycarbonate bottles (Tecniplast, Buguggiate, Italy)
Anaesthesia
Animals were anaesthetized by an intraperitoneal administration of 100 mg/kg ketamine (Ketasol) and 10 mg/kg xylazine (Xylasol, both from Dr E Graeub AG, Bern, Switzerland) per kg body weight (BW) and placed on a heating pad with a rectal probe to ensure a constant body temperature of 38°C.
Tail cut model
Groups of 16 (8 female and 8 male) mice received a single intravenous bolus injection of six different doses of test and of the corresponding dose of reference article in the lateral tail vein. Five minutes after the application of the test or reference item, 2 mm of the tail tip was cut off with a number 10 surgical blade. The tails were immersed in saline (approximately 37°C) and blood was collected over an observation period of 60 min. The amount of blood loss during the observation period, assessed gravimetrically in milligrams, was defined as the outcome measure. Surviving animals were allowed to recover from anaesthesia and returned to their cages. Buprenorphine (0.1 mg/kg BW) (Temgesic, Reckitt Benckiser Healthcare Ltd, Hull, UK) was injected subcutaneously at the end of the observation period for analgesia, and survival was assessed 1, 2 and 24 h after the tail tip cut. Mice were killed with CO2 inhalation at the end of the experiment.
Carotid arteria occlusion model
We studied groups of 6–16 animals (equal numbers of females and males) per dose. The left carotid artery was carefully dissected and then injured by placing a 1 x 1 mm filter paper (Whatman No. 3, Whatman International Ltd, Maidstone, UK) soaked in 15% FeCl3 solution onto the adventitia for 3 min to denude the endothelium. An FeCl3-mediated endothelial lesion initiates the coagulation cascade, which forms an occluding thrombus in wild-type mice. The test and reference article were administered intravenously (lateral tail vein) 15 min before injury to the vessel wall. Carotid blood flow was monitored before and after injury by using an ultrasound flow probe interfaced with a flow meter (Transonic Systems Inc, Ithaca, NY, USA) and a computerized data acquisition program (BDAS W, Hugo Sachs Elektronik - Harvard Apparatus GmbH, March-Hugstetten, Germany). Only animals with a baseline blood flow greater than 1 mL/min were included in the study. The mean carotid arterial blood flow was measured for 30 min after removing the filter paper, and the time to complete occlusion of the vessel was noted. The outcome was measured as time to occlusion in minutes, defined as the time required to permanent total occlusion of the vessel during the observation period of 30 min. An occlusion was defined as a decrease in blood flow to 0 ± 0.2 mL/min. Permanent occlusion was reached when the value 0 ± 0.2 mL/min did not increase again during the observation period. Where the measured values never reached 0 ± 0.2 mL/min or reached 0 ± 0.2 mL/min but increased again during the observation period, the outcome was rated as ‘no occlusion’ (>30 min) and corresponding results were used as censored observations. Mice were killed by cervical dislocation at the end of the observation period while still under ketamine/xylazine anaesthesia.
Test and reference articles
Recombinant human factor VIII (Advate, Baxter SA, Lessines, Belgium) was reconstituted with sterile water for injection (Baxter AG, Vienna, Austria) according to the manufacturer's instructions. The solution contained 175 IU rFVIII/mL after reconstitution and was further diluted with Advate dissolution buffer (Baxter AG, Vienna, Austria) to the concentration of application. The diluted rFVIII solutions were administered at six different doses (5, 10, 25, 50, 100 and 200 IU rFVIII/kg BW) in a volume of 10 mL/kg BW. Advate dissolution buffer was also used as the reference item.
Statistical analyses
The systematic dose effects of the two methods were compared by assessing the dose–response profiles of the models using relative dose effects (RDEs). All analysis were performed in R. 17 The RDE was defined as the outcome of a dose in FVIII knock-out mice relative to the outcome of buffer-treated C57BL/6J mice for a given method and was found through first ranking the responses across all doses, and second calculating the median rank per dose level divided by the median rank of buffer-treated C57BL/6J mice where ties were broken randomly. The dose–response profile for RDEs was visualized with R package drc using a four-parameter logistic function. 18 Time to complete occlusion with the COM between the 100 IU rFVIII/kg and the 200 IU rFVIII/kg BW dose was compared by an exact permutation test. 19
The variability of the two methods was compared using relative dose variation (RDV). The RDV measures the tightness of observations at a given dose in relation to other doses and can be regarded as a scale-independent variance. The RDV for a given method was calculated through the following steps: (1) rank the responses across all active doses, (2) scale the ranks by the number of observations and (3) find the variance of the scaled ranks for each dose. Ties were broken randomly to allow for upper limit censoring in the data, while scaling allowed for different sample sizes in each dose level. To compare RDV between methods, we assumed that the effect of a compound is the same, independent of the measurement method and accordingly the observed RDVs can simply be related to each other. Differences in variability were assessed by relative differences.
We used the ratio of the response at dose 100 IU rFVIII/kg BW in FVIII knock-out mice versus the response of buffer-treated C57BL/6J mice to compare the sample sizes and tested for non-inferiority using Fieller intervals, 20 which are based on the assumption of normal distributed observations. Non-inferiority can be demonstrated when the lower boundary of the one-sided 95% confidence interval (CI) for the ratio is above the prespecified margin of non-inferiority. We further assumed that the variation is equal for buffer-treated C57BL/6J mice and a dose of 100 IU rFVIII/kg BW administered in FVIII knock-out mice. Power for the hypothesis of non-inferiority was defined as the probability that the lower limit of the one-sided 95% CI for the ratio is above the prespecified margin of non-inferiority when both treatments are in fact identical. The sample size for showing non-inferiority was assessed using Monte Carlo simulations based on 1000 simulation runs.
The correlation between blood loss assessed in milligrams and mortality within 24 h (yes/no) was assessed by the point-biserial coefficient of correlation. 21 The point-biserial coefficient of correlation is mathematically equivalent to the Pearson (product moment) coefficient of correlation, that is, if there is one continuously measured variable such as blood loss and a dichotomous variable such as mortality within 24 h (yes/no). A two-sided 95% CI for the point-biserial coefficient of correlation was calculated by the bootstrap percentile approach based on 10,000 bootstrap replications. 22
Results
The RDEs were assessed to compare the dose–response profiles for the models. RDE was defined as the outcome of buffer-treated C57BL/6J mice/outcome of a rFVIII dose in FVIII knock-out mice.
Figure 1 visualizes the dose–response relation for the methods investigated. The dose–response profile with the COM indicates a classical sigmoid shape for the dose range investigated, whereas the maximum response is not observed with a dose of 200 IU/kg with the CUT. The RDEs were similar for doses up to 50 IU/kg BW (Figure 1). The dose effect with 100 IU/kg BW with the COM was three times larger than with the CUT model. The effect was reversed when a dose of 200 IU/kg BW was used, with a larger dose effect observed with the CUT (Table 1). The reason for the extremely high effect of the largest dose with the CUT method is not entirely clear. A possible explanation can be found in the Discussion section.
Summary of RDEs per model and dose levels of 5, 10, 25, 50, 100 and 200 IU rFVIII/kg BW
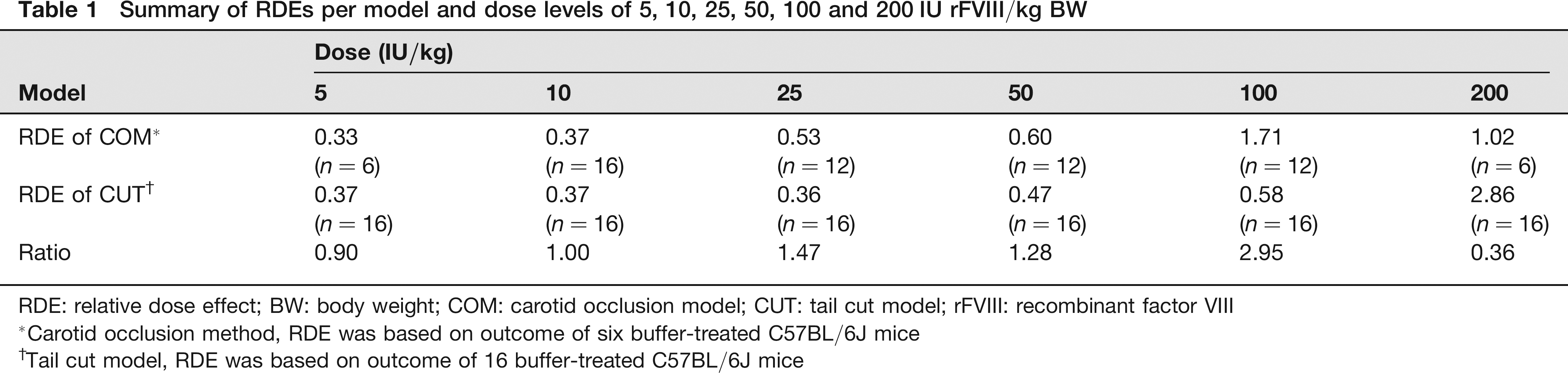
RDE: relative dose effect; BW: body weight; COM: carotid occlusion model; CUT: tail cut model; rFVIII: recombinant factor VIII
Carotid occlusion method, RDE was based on outcome of six buffer-treated C57BL/6J mice
Tail cut model, RDE was based on outcome of 16 buffer-treated C57BL/6J mice
Dose-response profiles for relative dose effects (RDEs) comparison of the tail cut model (CUT) and the carotid occlusion model (COM). The RDE measures the outcome in FVIII knock-out mice relative to the outcome of buffer-treated C57BL/6J mice. An RDE of 0.5 indicates that 50% of the effect with normal C57BL/6J was achieved with a given dose whereas an RDE of 1 with a specific dose indicates an identical effect to that of normal C57BL/6J mice. Sample sizes with the COM ranged from six to 16 animals per dose level investigated whereas with the CUT 16 animals per dose were studied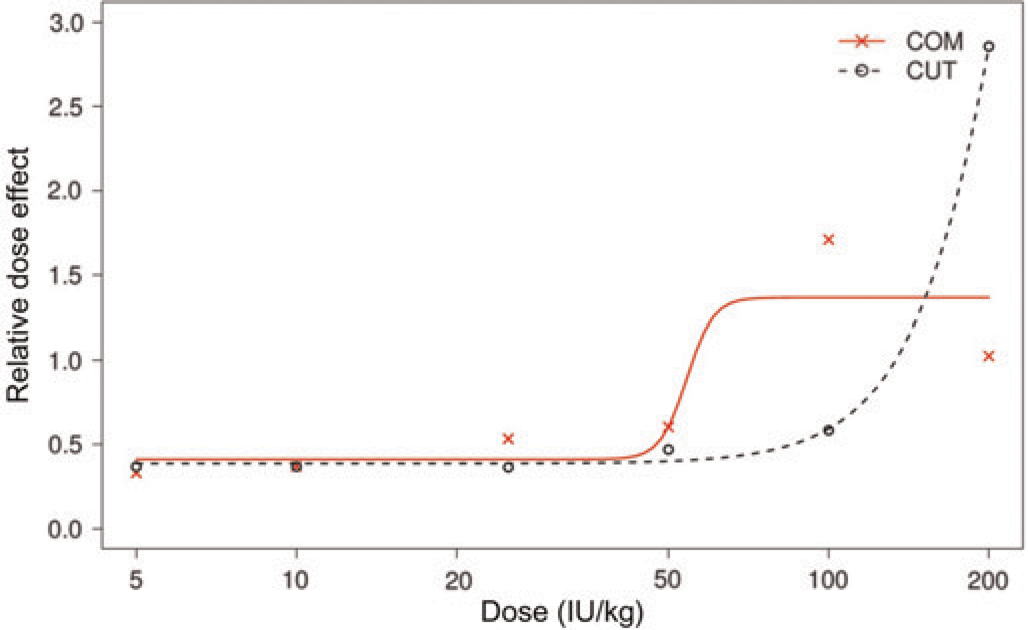
A larger effect was also found with the COM with the 100 IU/kg BW dose than with the 200 IU/kg BW dose (Table 1 and Figure 1), which was not statistically significant at the 5% level (two-sided
RDV was used to compare the variability between the methods. The reduction in variability for the COM over the CUT ranged from 63% to 82% (Table 2) depending on the dose given. As had been expected, in both models the lowest variation was seen with the lowest and not effective dose on the one side and with the highest (effective) dose on the other side. Variations were higher with doses in between the non-effective and effective doses (Figure 2). Variations ran somewhat in parallel for both models with the COM uniformly yielding significantly smaller variations.
Summary of RDV per model and dose levels of 5, 10, 25, 50, 100 and 200 IU rFVIII/kg BW
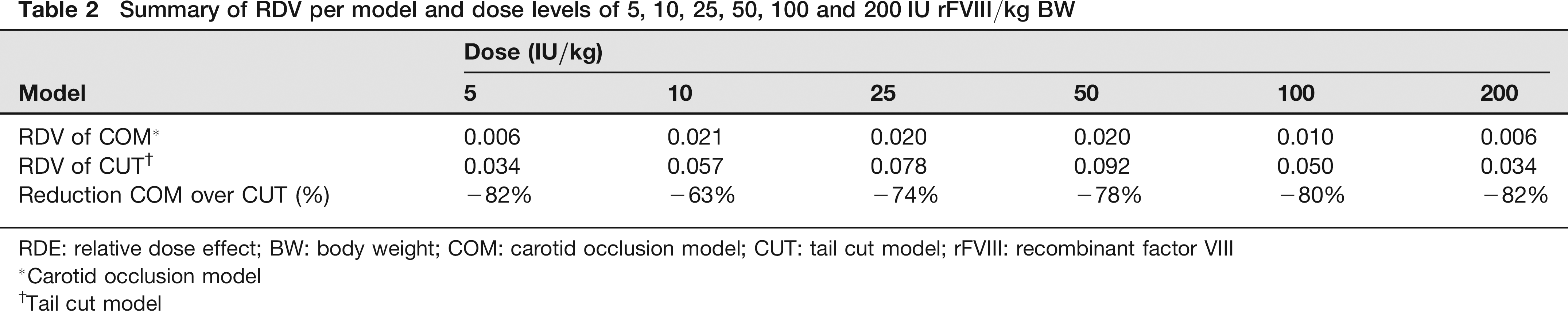
RDE: relative dose effect; BW: body weight; COM: carotid occlusion model; CUT: tail cut model; rFVIII: recombinant factor VIII
Carotid occlusion model
Tail cut model
Relative dose variation (RDV) comparison of the tail cut model (CUT) and the carotid occlusion model (COM). The RDV measures the tightness of observations at a given dose in relation to other doses. It is found as the variance of the adjusted ranks of a given dose. The adjusted ranking is determined based on the pooled sample and adjusted for the total number of observations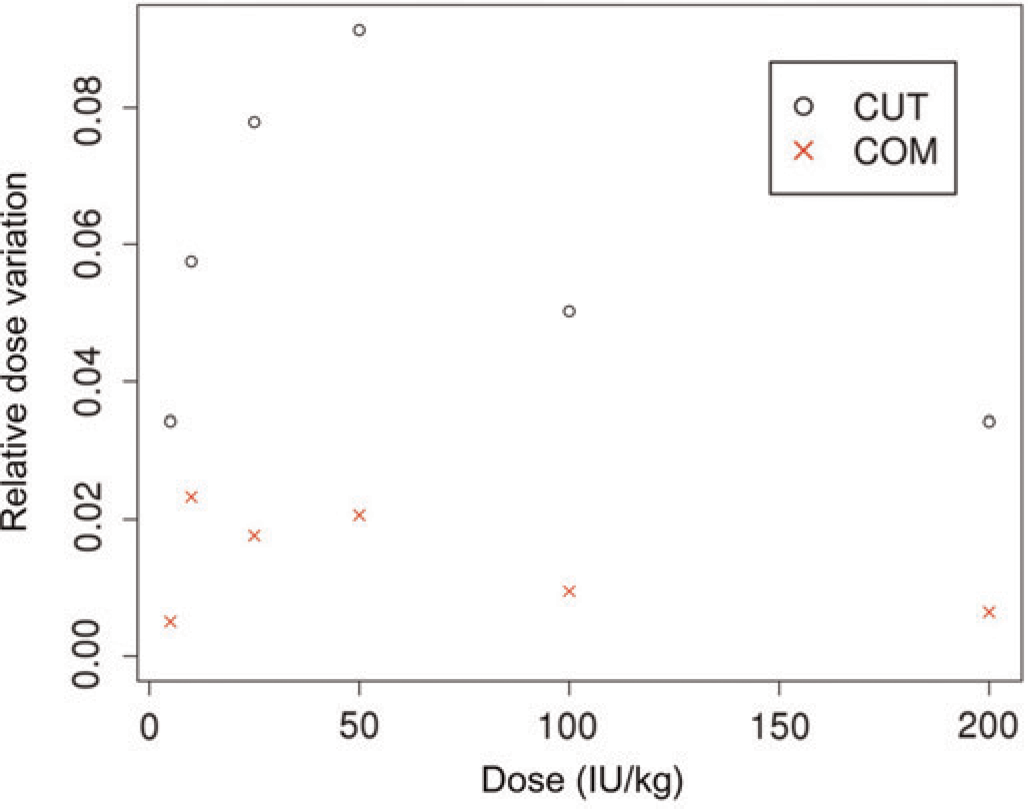
To determine the requisite sample size for the two approaches, we focused on showing non-inferiority of the effect between a dose of 100 IU rFVIII/kg BW in FVIII knock-out mice versus buffer-treated wild-type mice. Sample size estimation was based on observed coefficient of variations of buffer-treated C57BL/6J mice. Due to the large coefficient of variation (>100%) for the CUT method sample sizes were expected to be large and consequently the margins for non-inferiority varied between 0.7 and 0.9 while the type I error was set to 5% and the target power was chosen as 80%. The estimated sample size per group (Table 3) was enormous for the CUT as had been expected because the estimated coefficient of variation was 156%. The COM with a coefficient of variation of 33% required only about 5% of the sample size to establish non-inferiority across all margins.
Sample size per group required to establish non-inferiority between a dose of 100 IU rFVIII/kg BW and controls using varying margins of non-inferiority
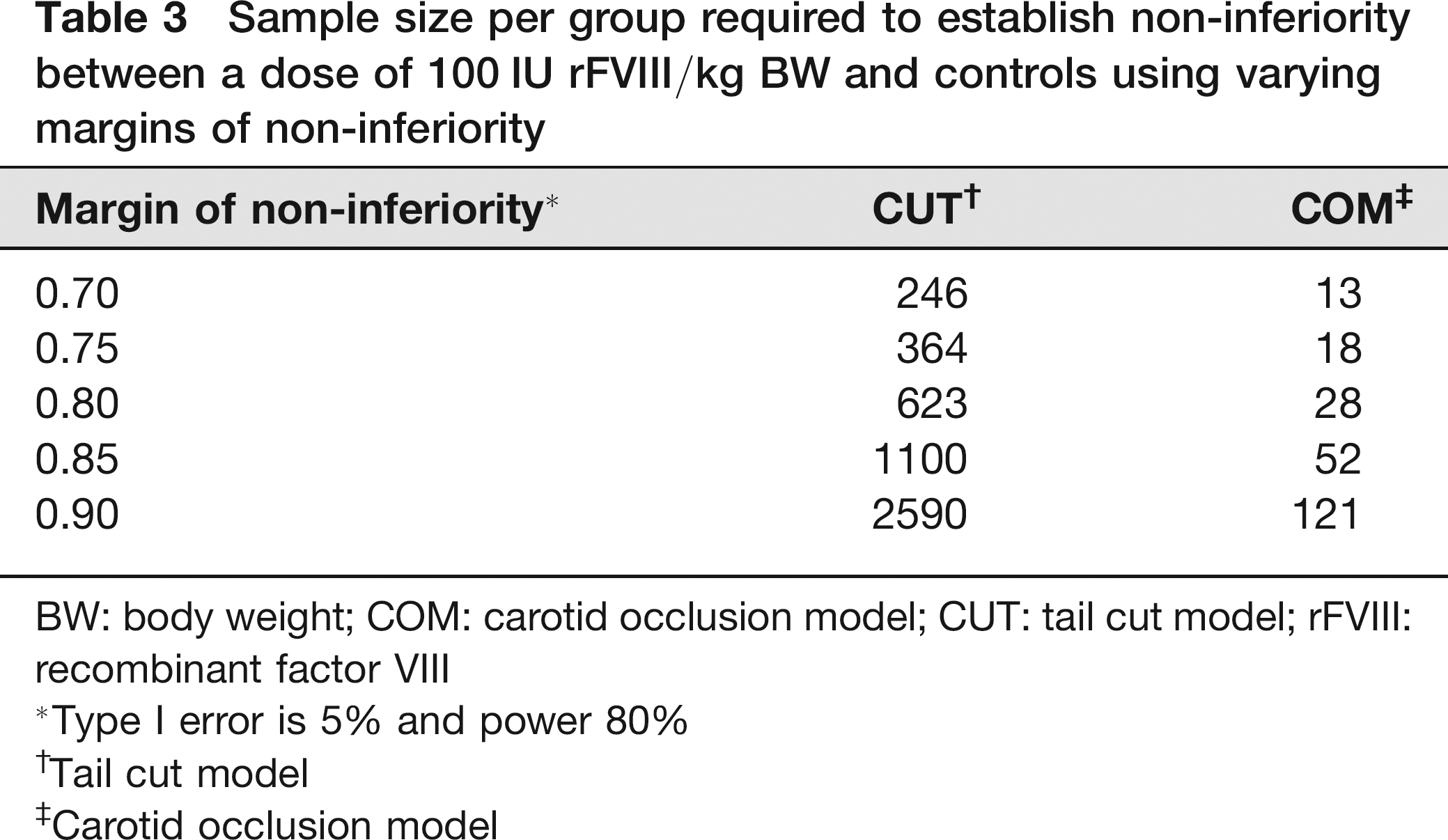
BW: body weight; COM: carotid occlusion model; CUT: tail cut model; rFVIII: recombinant factor VIII
Type I error is 5% and power 80%
Tail cut model
Carotid occlusion model
Finally, to evaluate the importance of mortality for the CUT, the correlation between blood loss and mortality was assessed using data from 128 animals (including 16 buffer-treated FVIII knock-out mice additional to the sample sizes given in Table 1). The point-biserial coefficient of correlation between blood loss assessed in milligrams and mortality within 24 h (yes/no) was 0.88 (95% CI: 0.82–0.93), indicating that mortality increases as blood loss increases. This result was statistically significant (at the 5% level) because the two-sided 95% CI for the coefficient of correlation did not contain the value 0.
Discussion
The CUT has long been a standard murine model for assessing efficacy of therapeutic agents for the treatment of haemostasis deficiencies while the FeCl3 COM is frequently used for
We also looked at the variability of the two methods to evaluate them further. Several published reports,23,24 as well as data from our own experiments have shown high standard deviations for blood loss in the CUT. The tail in rodents is crucial for thermoregulation and many factors can influence blood flow in the tip of the tail and therefore blood loss after injury. The peripheral vasomotor tone is very sensitive to changes in ambient temperature. The blood flow within the tail at normal room temperature is low but increases exponentially if ambient temperature rises. 25 Being a tool of thermoregulation the tail is well vascularized with a high density of arteriovenous anastomoses, particularly towards the tip. 26 Also, others and we have observed prolonged bleeding in wild-type mice, which is likely related to alternating periods of constriction and dilation of the central tail artery. 11 These facts plus anatomical variation between mice, like length and diameter of tail, may explain the differences between individuals. For example, in a recent report 27 vertebrae were counted in the tail of C57BL/6J mice at six weeks of age where mature vertebrae with end plates were present in 2 mm distal tail lengths at 21 days of age. 27 Other investigators found 28 at 10 weeks and up to 31 at 25 weeks of age in the same strain.28,29 Influences related to the depth of anaesthesia during the blood collection period may also contribute to the difficulties of standardizing the model.
Comparison of variation between the models shows the clear advantage of the COM. A reduction in variation of about 80% at effective dose levels led to our contention that fewer animals would be sufficient to prove treatment effects. We found that the COM requires only about 5% of the animals required by the CUT to establish non-inferiority between a dose of 100 IU/kg rFVIII in haemophilia mice and wild-type mice. This magnitude of fewer mice needed for experiments easily overcomes the drawback of microsurgery intervention that is necessary for the COM. Although there was a learning curve for the surgical procedure during model establishment, all mice survived vessel preparation and reached the necessary baseline blood flow at the beginning of the experiment in this study (3 mice died during the 30 min observation period and were replaced).
Apart from the initial blood loss, rates of survival of haemophilic mice at different time points after the tail cut are commonly taken into account because an initial platelet plug is possible in haemophilic mice but clot stabilization by a fibrin meshwork is impaired resulting in a possibility of delayed rebleeds. These rebleeds are assumed to be the cause of death through excessive blood loss after primary haemostasis at the site of injury. Certainly this is plausible but we believe that this measurement is heavily influenced by the initial blood loss and not so much by rebleeds. Although treatment of haemophilic mice with various preclinical therapeutic candidates may lead to less blood loss, a substantial amount of blood is lost, depending on the length of cut tail, even in wild-type mice (data available upon request), which severely affects the animal's wellbeing after recovery from anaesthesia. Pain induced by the amputation might be difficult to detect, especially if behaviour is influenced by blood loss. However, abnormal changes in behaviour, depending on strain and age, were observed 60 min after biopsy in a study to evaluate tail biopsy methods. 27 The strong correlation found between the initial blood loss and survival after 24 h indicated that the relevance of this endpoint is questionable in this context. Waiving this endpoint should be considered if the recovery period is not optimized by fluid substitution, analgesics and precautions to keep body temperature at physiological levels. Especially, if rebleeds are not actually observed and recorded during this period. The COM offers an easy possibility to judge clot stability. The restart of blood flow in the carotid artery after cessation due to the initial occlusive clot, which is automatically recorded and available for analysis in the COM, can be considered as a ‘rebleed’ event.
Comparison of the COM and CUT showed that both models could be used to evaluate potential therapeutic agents for haemophilia A in a FVIII knock-out mouse model. In addition, the COM proved to be the more sensible method for showing treatment effects at clinically relevant doses. However, the biggest advantage is the ability to better standardize the COM and therefore circumvent the high variability associated with the CUT. This allows for an enormous reduction in animals necessary to statistically verify a phenotypic correction by substitution of FVIII in haemophilic mice. The inability to check for survival with the COM is not viewed as a disadvantage because this is influenced by the initial blood loss, which makes this endpoint redundant unless rebleeds are actually observed and recorded. The return of blood flow in the COM might be a more objective way to assess clot instability. Overall our results clearly demonstrate that the COM is a powerful and valid tool for