
Abstract
Visualization and manipulation of defined motoneurons have provided significant insights into how motor circuits are assembled in Drosophila. A conventional approach for molecular and cellular analyses of subsets of motoneurons involves the expression of a wide range of UAS transgenes using available GAL4 drivers (eg, eve promoter-fused GAL4). However, a more powerful toolkit could be one that enables a single-cell characterization of interactions between neurites from neurons of interest. Here we show the development of a UAS > LexA > QF expression system to generate randomly selected neurons expressing one of the 2 binary expression systems. As a demonstration, we apply it to visualize dendrite-dendrite interactions by genetically labeling eve+ neurons with distinct fluorescent reporters.
Significance Statement
The study of how motor circuits are assembled in the Drosophila brain is limited by the genetic tools available to target and manipulate individual neurons. We present a genetic toolkit that allows us to generate LexA- and QF-expressing clones randomly in eve+ motoneurons. Here, we demonstrate the utility of this system in visualizing dendrite-dendrite interactions between eve+ clones at high resolution. We anticipate that this system will become an indispensable tool to study interactions between distinct clones in unique cell types.
Introduction
Revealing the circuit principles underlying specific behavior is one of the primary goals of neuroscience research. Particularly in Drosophila, there is a vivid interest in dissecting the motor circuit.1,2 We have previously focused on eve-positive (eve+) neurons, aCC and RP2, which are segmentally repeated in the ventral nerve cord (VNC).3-5 Both motoneurons innervate body wall muscles (Figure 1a, dots). These neurons also extend their dendrites to overlapping fields across segments in the VNC (Figure 1a), 6 suggesting dendrite-dendrite interactions between the neurons. However, little is known about how contacts between dendrites from neighboring neurons influence the formation of dendritic fields. A toolkit with which one can study interactions between eve+ neurons would be of particular interest. Here, we describe a new system genetically manipulating these neurons by 2 binary expression systems (ie, LexA/LexAop and QF/QUAS).7,8

The> LexA > QF mosaic expression system: (a) Gross anatomy of eve+ neurons in the ventral nerve cord (VNC) at the 2 instar larva stage. Dots indicate the VNC exit points of eve+ neurons. (b) Schematic of the >LexA > QF system. Cre induces an excision event between LoxP sites. Before Cre action, only LexA is expressed. Cre-mediated recombination switches expression to QF. (c) S2 cells transiently transfected with pUAS > LexA > QF. Co-transfected with pUAS-Cre, these cells switch to QF expression. We use the actin 5c ubiquitous promoter to drive GAL4, which in turn activates the expression of genes under the control of UAS. We also co-transfect reporter plasmids, that is, LexAop-GFP (green) and QUAS-RFP (red). There are 2 cells shown in yellow*; the red and green cells contact each other. Scale bars: 10 µm.
Materials and Methods
Generation of transgenic flies
The pUCIDT > LexA > QF plasmid was generated through synthesis and molecular cloning by Integrated DNA Technologies, Inc. To generate pUAS > LexA > QF, we PCR-amplified the >LexA > QF cassette and cloned it into the backbone of pACUH (Addgene # 58374) via the BglII and XbaI sites. To generate QUAS-myr::mCherry, we PCR-amplified the myristoylation sequence from pJFRC19-13XLexAop2-IVS-myr::GFP (Addgene # 26224) and placed it in the pQUAST-mCherry vector via the NotI and BamHI sites. After verifying these sequences, we generated transgenic flies using a transposon-based approach (Rainbow Transgenic Flies, Inc).
Cell culture
Drosophila Schneider S2 cells were cultured at 25°C in an SFX-INSECT cell culture medium (HyClone). An actin 5C-GAL4 driver, pUAS > LexA > QF, and fluorescent reporter plasmids (100 ng of each plasmid per well) were co-transfected by using Effectene (2.5 μl, QIAGEN) into an 8-well Lab-Tek II chambered #1.5 coverglass system (Nalge Nunc International).
Fly genetics and husbandry
Flies were maintained at room temperature on standard cornmeal-agar food. Full genotypes of stock used in Figures 1a and 2b: (

Fluorescent reporter expression in >LexA > QF larvae: (a) Example crossing schemes for the generation of LexA and QF neurons using UAS > LexA > QF. (b) Confocal projection of the VNC with Cre-mediated expression of LexA (green) or QF (red) in eve+ clones. Note that we see a variety of FP expression levels. Asterisks indicate eve+ neurons that express neither LexA nor QF. Insets show higher magnification of boxed regions. (c) Quantification of GFP+ and RFP+ clones in eve+ neurons. The pie chart shows the percentage of different clones in the same third instar larva. This experiment was replicated 5 times independently. Scale bars: 10 µm.
Immunohistochemistry
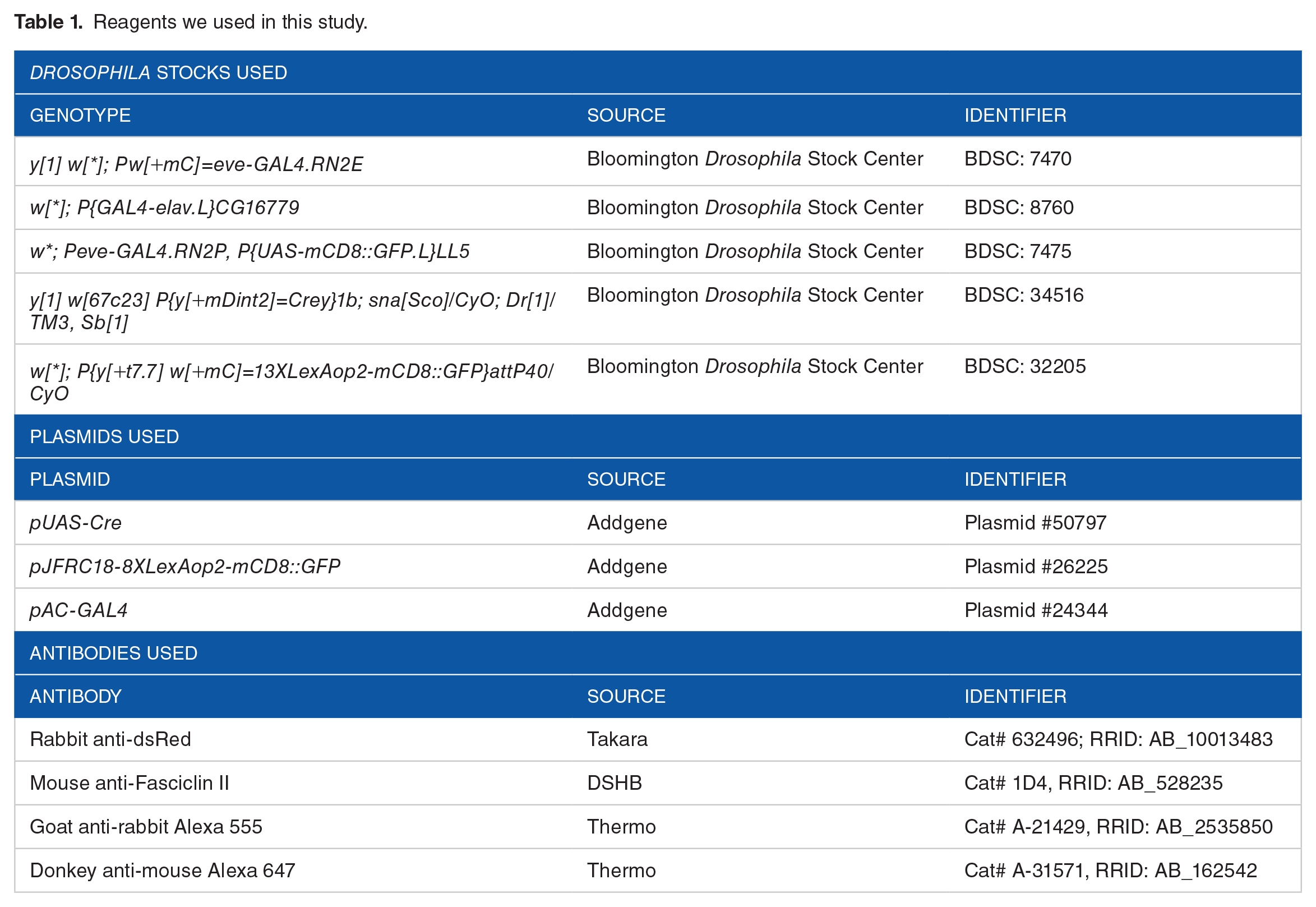
Fly larvae were dissected in PBS and fixed with 4% paraformaldehyde in PBS for 30 minutes at room temperature. Brains were washed in TBS (1XPBS + 0.1% Triton X-100), blocked for at least 1 hour in blocking solution (1XPBS + 0.1% Triton X-100 + 1% bovine serum albumin), and incubated in primary antibodies at 4°C overnight. On day 2, brains were washed 3 times in TBS and then incubated in secondary antibodies in TBS for 2 hours at room temperature. After extensive wash in TBS, brains were mounted in 50% glycerol for slide preparation. See also Table 1 for antibodies used.
Reagents we used in this study.
Confocal imaging
Confocal images of neurons were captured using an inverted fluorescence microscope (Ti-E, Nikon) with a 10× or 100× objective (Nikon). The microscope was attached to the Dragonfly Spinning disk confocal unit (CR-DFLY-501, Andor). Images were recorded with an EM-CCD camera (iXon, Andor). All images in figures are maximum Z-projections of confocal images (FIJI/ImageJ, NIH).
Results and Discussion
It has been demonstrated that eve-GAL4 can drive the expression of UAS transgenes in every aCC and RP2 neuron. 9 Shown by GFP expression via eve-GAL4, GFP+ dendrites from eve+ neurons extensively overlap during larval stages (Figure 1a). Unless these neurons are labeled with different FPs in a mosaic manner, the use of this GAL4 driver is challenging to investigate dendrite-dendrite interactions between these neurons. We thus design a new mosaic expression system (Figure 1b). This system exploits mitotic recombination by a Cre enzyme to generate 2 distinct clones under the control of GAL4 drivers. A similar design has already been adopted in several techniques to generate mosaic neurons, including UAS-dBrainbow. 10 Whereas the Brainbow techniques can label a subpopulation of neurons with fluorescent reporters, they cannot be used to examine mosaic phenotypes in the context of genetic knockdown or over-expression. Thus, we sought to apply 2 transcriptional activators (ie, LexA and QF) to the mosaic system instead of fluorescent reporters. In this system, Cre-mediated recombination between 2 loxP sites leads to the excision of the LexA sequence, which subsequently induces QF expression (Figure 1b, bottom). By contrast, no recombination gives rise to LexA expression (Figure 1b, top).
To assess whether the >LexA > QF cassette functions in Drosophila S2 cells, we created the pUAS > LexA > QF expression construct and drove the expression by using a ubiquitous actin 5C-GAL4 driver construct. We also employed 2 reporter plasmids for LexA and QF transcriptional activators, such as LexAop-GFP and QUAS-RFP, respectively. In the absence of Cre, we observed only GFP+ cells (Figure 1c). This observation is consistent with the notion that the cassette retains the LexA sequence without Cre expression. For comparison, when we co-transfected >LexA > QF with Cre, we observed RFP expression from some cells (Figure 1c). Note that other cells also expressed GFP due to a lack of recombination (the Cre plasmid would not be introduced into these cells because of co-transfection failure; Figure 1c). Altogether, these experiments demonstrate that pUAS > LexA > QF enables the expression of these transcriptional activators in different groups of cells.
To test whether this expression system functions in Drosophila in vivo, we generated flies harboring UAS > LexA > QF, fluorescent reporters, and eve-GAL4 (see also the crossing scheme in Figure 2a). Using the flies in the absence of Cre recombinase, 100% of eve+ neurons expressed only LexA as assessed by fluorescent reporters (ie, LexAop-GFP and QUAS-RFP) (Figure 2b, left). When Cre was expressed, recombination induced RFP expression (Figure 2b, right). As described in the crossing scheme (Figure 2a), Cre expression was under the control of the hsp70 promoter. 11 It has been reported that loxP sites can be recombined by hs-Cre with ~100% efficiency in cells at room temperature. 11 However, we empirically identified an optimal temperature (such as 16°C) at which Cre-mediated recombination happens in a mosaic pattern (Figure 2b, right). Among animals examined, the selection between these activators appears to be stochastic and independent in each neuron. To quantify the choice of these activators, we measured the number of GFP+ and RFP+ neurons. We found that both reporters were expressed at similar relative frequencies (Figure 2c).
We also noticed that a couple of neurons expressed neither GFP nor RFP in the same animals (Figure 2b,
Nonetheless, as seen in Figure 2b, this system labels individual neurons whose dendritic branches can be visualized in great detail. Analogous results were also observed using a serotonergic neuron-specific Trh-GAL4 (KB and DK, unpublished data). We thus believe that this expression system will be broadly applicable in its current form.
The >LexA > QF system provides a facile platform for studying the interaction between genetically manipulated neurons. A major advantage of this system over previous techniques such as dBrainbow is that we could examine phenotypes elicited by the expression of transgenes in given clones. For example, with the expression of RNAi transgenes (the crossing scheme is outlined in Figure S2a), we would investigate molecular mechanisms of how dendrites are established and patterned via dendrite-dendrite contacts. In such an experiment, one might choose room temperature (25°C) to enhance RNAi efficacy. Another way of recombination can circumvent the low-temperature requirement. In this case, one would use a different Cre variant such as a ligand-inducible DD-Cre (Cre is fused to the
The >LexA > QF system could also get along with a variety of fluorescent reporters. In conjunction with the GFP reconstitution across synaptic partners (GRASP) system (see also the crossing scheme in Figure S2b), it would allow us to characterize the sites of dendrite-dendrite interactions. The recent advent of the split GFP system furthermore enables labeling of proteins endogenously expressed in subsets of neurons. 15 The super-folder GFP sequence is split between 10th and 11th β-strands (GFP1-10 and GFP11). These fragments do not individually fluoresce, but upon associating each other, they can reconstitute a GFP signal. Only the GFP11 fragment is inserted into a specific genomic locus via a custom-design genome editing strategy or a genomic integration into an available MiMIC site. By introducing the expression of the large GFP1-10 fragment into eve+ clones, any clone expressing both fragments can be fluorescently labeled (see the crossing scheme in Figure S2c). This way, we could investigate how endogenous proteins dynamically localize during dendrite-dendrite interactions between eve+ neurons.
Supplemental Material
sj-docx-1-exn-10.1177_26331055211069939 – Supplemental material for A Genetic Toolkit for Simultaneous Generation of LexA- and QF-Expressing Clones in Selected Cell Types in Drosophila
Supplemental material, sj-docx-1-exn-10.1177_26331055211069939 for A Genetic Toolkit for Simultaneous Generation of LexA- and QF-Expressing Clones in Selected Cell Types in Drosophila by Kota Banzai, Ping Shen and Daichi Kamiyama in Neuroscience Insights
Footnotes
Acknowledgements
We thank the Bloomington Drosophila Stock Center for fly lines. For comments on the manuscript, we thank all members of the Kamiyama lab; we particularly thank MA. Inal for critical reading and editing of this manuscript.
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This was supported by an NIH grant (NINDS R01NS107558).
Declaration of conflicting interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
Kota Banzai: Conceptualization, Methodology, Investigation, Formal analysis, Visualization, Writing—review and editing. Ping Shen: Conceptualization, Writing—review and editing. Daichi Kamiyama: Conceptualization, Supervision, Writing—original draft, Writing—review and editing, Funding acquisition.
Data Availability
The pUAS > LexA > QF plasmid and sequence have been deposited in Addgene. The UAS > LexA > QF transgenic line is available from the Bloomington Drosophila Stock Center.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.