
Abstract
Gliomas are brain tumors originated from glial cells. The most frequent form of glioma is the glioblastoma (GB). This lethal tumor is frequently originated from genetic alterations in epidermal growth factor receptor (EGFR) and PI3K pathways. Recent results suggest that signaling pathways, other than primary founder mutations, play a central role in GB progression. Some of these signals are depleted by GB cells from healthy neurons via specialized filopodia known as tumor microtubes (TMs). Here, we discuss the contribution of TMs to vampirize wingless/WNT ligand from neurons. In consequence, wingless/WNT pathway is upregulated in GB to promote tumor progression, and the reduction of these signals in neurons causes the reduction of synapse number and neurodegeneration. These processes contribute to neurological defects and premature death.
Gliomas are the most frequent tumors of the central nervous system originated from neoplastic glial cells. The most common type of glioma is the glioblastoma (GB); this aggressive and infiltrative brain tumor displays a glial network of cellular protrusions known as tumor microtubes (TMs). GB patients have a poor prognosis associated with the formation of TMs that contribute to radiotherapy resistance, cell proliferation, and neurodegeneration. 1
These tumors are characterized by a high heterogeneity, as they constitute different entities of the disease; GB affects patients of different ages and develops from different genetic alterations, even within the same tumor. 2 However, different studies reveal altered mutations or metabolic pathways that are maintained in most GB clinical cases such as the epidermal growth factor receptor (EGFR) gene and phosphatidyllinositol-3-kinase (PI3K). 3 The most common mutations in primary GBs involve EGFR amplification and overexpression. EGFR is one of the first pro-oncogenic factors identified in GB patients, as it serves as a central regulator role of the neural stem cell. Mutations cause constituent kinase activity that induces proliferation, cell migration, and blockage of apoptosis. The other most common mutations in GB affect the PI3K signaling path, loss of lipid phosphatase or PTEN, or the presence of Akt in its constituently active form. Both pathways are related and must be co-altered in Drosophila glial cells to reproduce the transformation of glial cells, the development and progression of a GB, and the associated neurodegeneration. 4
In consequence, a great number of clinical strategies were designed against EGFR or PI3K or members of these signaling pathways. EGFR is sensitive to tyrosine kinase inhibitors that have been proposed as potential treatment against EGFR signal transduction in GB. Besides, monoclonal antibodies that recognize EGFR epitopes have been also proposed against GB, with special consideration given to immunological approaches, the use of EGFR as a docking molecule for conjugates with toxins, T-cells, oncolytic viruses, exosomes, and nanoparticles. But, unfortunately, all attempts to target EGFR in GB patients have been unsuccessful. 5
Besides, pathways linking receptor tyrosine kinases, PI3 kinase, Akt, and mTOR represent therapeutic targets against GB. 6 Small molecules against one or combinations of these kinases have been tested; unluckily, preclinical studies demonstrate low or noncytotoxic effect on GB cells. 6 Current trends point toward combined treatments, including cytotoxic therapies, survival signaling inhibitors, targeted therapies, and radiotherapy. However, further knowledge and novel strategies are a need to tackle GB progression.
We have recently published our latest results on GB progression and associated neurodegeneration. 4 Our results in a Drosophila melanogaster model indicate that TM network in GB triggers a tumor-cell communication involved in the transport of proteins or signals that promote tumor growth, tumor-associated neurodegeneration, and resistance to current treatment. In particular, TMs mediate Wingless (Wg)/WNT signaling imbalance in surrounding healthy neurons in favor of GB cells. The reduction of Wg/WNT signaling in neurons causes neurodegeneration, which is of great relevance for life span in GB progression. Besides, Wg/WNT input promotes GB cells increase and triggers a positive feedback loop that also includes JNK pathway and matrix metalloproteases (MMPs) that contribute to TMs’ expansion and infiltration (Figure 1). 4 Other groups have also described the intimate relation of GB cells with neurons by synaptic contacts that modulate the behavior of GB cells,7,8 supporting the relevance of cellular communication in GB progression.

Glioma-neuron signaling imbalance mediated by TMs. Glial cells are initially transformed into malignant GB on EGFR and PI3K pathways’ constitutive activation that stimulates the actin cytoskeleton remodeling and enables initial expansion of TMs. Glioma cells expand the network of TMs that accumulate Fz1, surround neighboring neurons, and facilitate neuronal-Wg vampirization mediated by glioma Fz1 receptor. Afterward, GB cells establish a positive feedback loop, including TMs, Wg signaling, JNK, and MMPs. TM-mediated neuronal-Wg depletion results in Wg signal extinction in neurons and Wg signaling up-regulation in the GB cells, which activates JNK pathway in GB. Therefore, MMPs are upregulated and facilitate further TM infiltration in the brain, and the GB TM network expands and mediates further Wg depletion to close the loop, resulting in neurodegeneration.
A common and relevant cellular feature of GBs is the formation of membrane protrusions (TMs) that mediate cell-to-cell contact. These TMs, first described in human GB cells 1 and later in a Drosophila model of GB, 4 arise as key players in gliomagenesis and the interactions with surrounding tissues. TMs are sensitive to downregulation of the JNK pathway, MMPs, or Gap43 knockdown (Figure 2), and antagonists of TM formation emerge as promising candidates against GB.
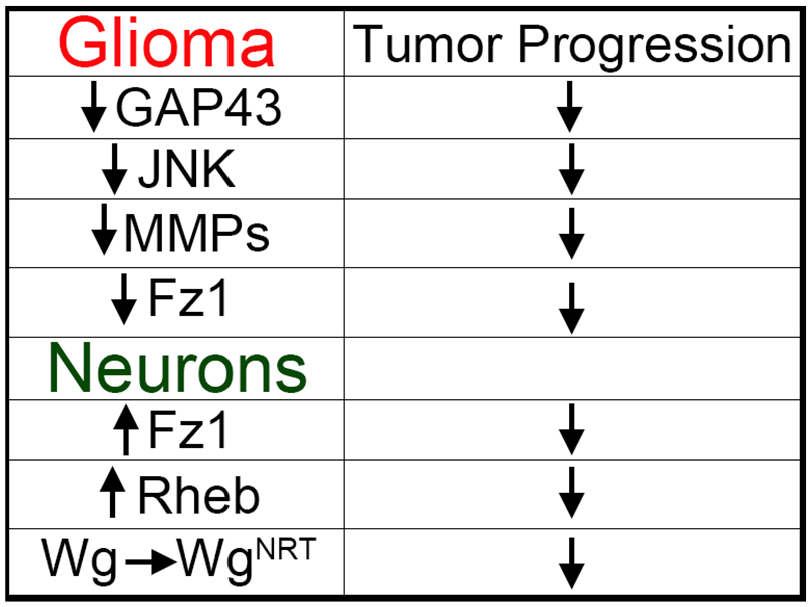
Nonfounder mutations involved in GB progression. Manipulation of signaling pathways involved in the regulation of TMs in GB cells such as downregulation of Gap43, JNK, or MMPs is sufficient to prevent tumor progression. Moreover, impairment of Wg vampirization by downregulation of Fz1 in GB cells or substitution of endogenous Wg for a nonsecretable Wg in the neurons also prevents tumor progression. In addition, modifications in neurons such as Fz1 overexpression that facilitate the competition for Wg between GB and neurons or the overexpression of Rheb also prevent tumor progression nonautonomously.
On the contrary, the bidirectional relation of GB cells and neurons establishes a novel dimension for tumor control. Preliminary results suggest that the physiological status of neurons can affect the progression and aggressiveness of GB. Our experiments show that the overexpression of Wg/WNT receptor Frizzled1 (Fz1) in neurons stimulates Wg signaling in neurons. This single modification in healthy tissue facilitates the competition for Wg between GB and neurons. In consequence, the restoration of Wg signaling equilibrium prevents synapse loss and neurodegeneration, and reduces GB progression (Figure 2). 4 Moreover, recent results from our group have established that activation of PI3K pathway in neurons by means of Rheb overexpression prevents mitochondrial defects and synapse lose, and prevents the reduction in life span caused by GB (Figure 2). 9
Altogether, these data indicate a central role for the filopodia-like membrane protrusions (TMs in tumoral cells) in the regulation and maintenance of the signaling equilibrium between the neurons and glial cells. However, if a GB arises, the transformed glial cells expand and take advantage of the TM network to vampirize signaling molecules from the surrounding neurons to sustain tumor progression (Figure 1), highlighting once again the critical role of the tumor microenvironment in cancer.
The classical view has been that cells use membrane protrusions to probe the environment for cues, and for migration and motility. 10 The role of filopodia in migration is well established in many physiological processes like wound healing, angiogenesis, chemotaxis, embryonic development, and adhesion. Cell motility requires cells to adapt to, interact with, and often modify their surrounding extracellular matrix (ECM). Cells use transmembrane and growth factor receptors that cluster and recruit cytoplasmic adapters and signaling molecules to enable the coordinated regulation of plasma membrane dynamics through the remodeling of the actin cytoskeleton that allow cells to restructure their microenvironment. 11
In addition, filopodia have been extensively studied about their role in motility and migration in cancer cells. Increased filopodia formation has been shown to promote migration, 12 and their abundance is a characteristic of invasive carcinoma cells. Interestingly, filopodia are involved in the activation of the WNT signaling pathway during the progression of colorectal cancer. 13
The formation of filopodial actin filaments is controlled by the actin nucleating proteins formins and the actin-related proteins complex Arp2/3. 14 Formins are a group of Rho GTPase effectors involved in adhesion, migration, cytokinesis, and cell polarity. 15 In melanoma cells, overexpression of the Formin Dia2 induces filopodia, and loss of Dia2 inhibits filopodia formation. 16 Arp2/3 induces filopodia formation by promoting the branching of the actin filaments, 14 and ENA/VASP family of proteins regulates actin filament elongation by recruiting actin nucleating factors such as Arp2/3 and profilin. 16 Rif, another Rho family GTPase, is also a potent stimulator of membrane protrusions and its ability to induce filopodia has been shown to be dependent on Dia2.14,16 Moreover, proangiogenic signals, such as vascular endothelial growth factor (VEGF), also trigger filopodia formation in the tip cells during angiogenic sprouting. 16 Mena, a member of ENA/VASP family, is upregulated in the invasive and metastatic populations of breast cancer cells and its isoform Mena (INV) sensitizes cells to EGF and increases the matrix degradation in tumor cells. 16
Most data about the role of filopodia in cancer and the mechanisms and molecules involved in filopodia biology focused on their role in motility, migration, and infiltration. Consistently, the recent studies discussed above showed that TM contributes to invasion, proliferation, brain colonization by GB cells. 1 Moreover, TMs form a multicellular network that connects GB cells over long distances, a feature that likely provides resistance against radiotherapy, chemotherapy, and surgery. 1 However, glial cytonemes/TMs are also involved in intercellular communication that alter Wg/WNT signaling between glia and neurons in Drosophila. These novel data evidence additional roles for these membrane structures in communication with the tumor microenvironment and engaging the microenvironment in sustaining and promoting tumor progression. In line with this, a recent study showed that cytoneme-mediated signaling between an epithelial EGFR-driven tumor and its adjacent stromal tissue is essential for tumor progression, and that genetic ablation of cytonemes is sufficient to prevent tumor progression in the Drosophila wing epithelium. 17 It would be very interesting to revisit previous studies that showed a key role for membrane filopodia in tumor progression and metastasis and analyze a possible role of these membrane structures in communication and signaling, particularly in long-distance intertissue communication.
The examples commented above are evidences of intertissue communication mediated by membrane filopodia/TMs in a tumor model. However, tumoral cells do not create novel structures, and GB cells take advantage of the filopodia already present in normal cells and enhance their activity. Cytonemes were identified as signaling-specialized filopodia in Drosophila and vertebrates, and they are involved in several developmental systems such as during the formation of signaling gradients in Drosophila imaginal disks, during development of the chicken embryos, the zebrafish neural plate, or in the patterned differentiation of specialized cells such as the stripe formation on zebrafish or the patterning of bristles of the Drosophila adult thorax. 18
Focusing on the neural tissue, filopodia were first identified for their roles in cell migration and chemoattractant sensory mechanisms in the neuronal growth cones. 18 Filopodia are also important for establishing spines, synapses, and for the development of dendritic branches and the shaft synapses on them. In addition, in the Drosophila optic lobe lamina, the growth cone filopodia of the photoreceptors R1–R6 continuously interact with neighboring growth cones during development. 18
Nevertheless, there are also physiological examples of intertissue communication mediated by cytonemes. Astrocytes extend thin membrane projections around synapses that mediate the communication with neurons. 18 Besides, almost immediately after central nervous system (CNS) injury, microglia extend filopodia toward the site of damage and undergo a dramatic morphologic and phenotypic transformation during regeneration. 19 Moreover, cytonemes containing the Fz1 receptor from the Drosophila air sac primordium (ASP) cells contact the Wg-producing epithelial cells in the wing disk for Wg signaling activation. Cytonemes from myoblasts carrying the Notch ligand, Delta (Dl), also contact the ASP to activate Notch signaling. In addition, ASP cells extend thickveins (Tkv)-containing cytonemes toward wing disk decapentaplegic (Dpp)-producing cells, while also directing fibroblast growth factor receptor (FGFR)-containing cytonemes toward FGF expressing wing disk cells. 18 Furthermore, epithelial filopodia-like protrusions in the chicken embryo are involved in ectodermal-dermomyotomal WNT signaling regulation via transport of Frizzled-7, providing functional long-range intercellular signaling during somite development. 20
It is now evident that cytonemes/TMs have many interesting roles other than motility and migration, including long-range intertissue communication to coordinate many cells in multicellular organisms during development and homeostasis, although they can provide many advantages, such as overcoming physiological barriers or controlling the surrounding healthy cells in the tumor microenvironment, for tumoral cells.
In conclusion, cytonemes/TMs emerge as potential targets for cancer treatment and bring a novel perspective for cancer biology, and it is important to decipher the specific mechanisms underlying cytoneme formation and regulation. Moreover, tumors like the GB have the ability to control and adapt the surrounding microenvironment, in this case the neurons, and upregulate signaling pathways via cytonemes at expenses of the surrounding healthy neurons. Therefore, healthy tissue contributes to tumor progression. We propose that modifications of healthy tissue can exert a positive effect for neurons and can modify tumor progression. Future studies will determine whether these strategies, in combination with other treatments, are effective against GB progression and contribute to restore the damage caused by this fatal tumor.
Footnotes
Funding:
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
Marta Portela and Sergio Casas-Tintó are contributed equally.