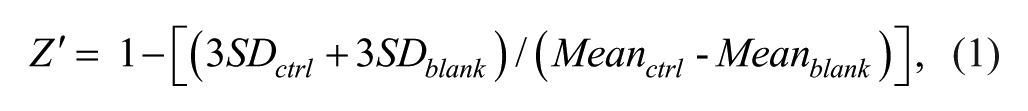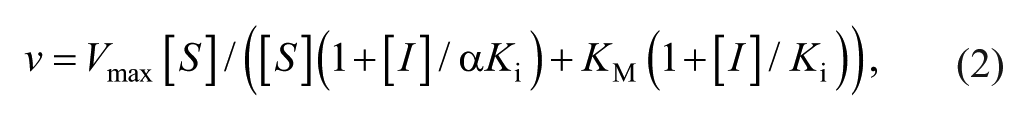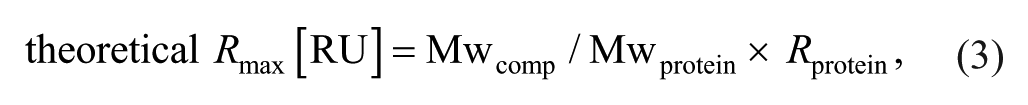Peripheral arterial disease (PAD) is an occlusive disease that can lead to atherosclerosis. The involvement of arginase II (Arg II) in PAD progression has been proposed. However, no promising drugs targeting Arg II have been developed to date for the treatment of PAD. In this study, we established a method for detecting the activity of Arg II via high-throughput label-free RapidFire mass spectrometry using hydrophilic interaction chromatography, which enables the direct measurement of l-ornithine produced by Arg II. This approach facilitated a robust high-concentration screening of fragment compounds and the identification of a fragment that inhibits the activity of Arg II. We further confirmed binding of the fragment to the potential allosteric site of Arg II using a surface plasmon resonance assay. We concluded that the identified fragment is a promising compound that may lead to novel drugs to treat PAD, and our method for detecting the activity of Arg II can be applied to large-scale high-throughput screening to identify other structural types of Arg II inhibitors.
Peripheral arterial disease (PAD) is an important manifestation of atherosclerosis, which is closely linked to cardiovascular events and increased all-cause cardiovascular mortality.1 Approximately 155 million people had PAD worldwide in 2015.2 PAD is accompanied by a narrowing of the peripheral arteries in the legs, and medical interventions have been strongly desired because severe PAD causes walking difficulties due to intermittent claudication. Cilostazol, a selective phosphodiesterase 3 inhibitor, is the only clinically proven drug that improves walking distance in patients with PAD;3 however, in addition to not being a causal treatment for PAD, it should be avoided in patients with cognitive heart failure.4
PAD pathogenesis is closely linked to vascular endothelial dysfunction, followed by impaired vasodilation and vascular smooth muscle cell differentiation/proliferation. Nitric oxide (NO) is a potent antiatherosclerotic vasodilator produced by vascular endothelial cells that mediate vessel relaxation, resulting in protection against cardiovascular events.5 NO synthase (NOS) is the rate-limiting enzyme of NO synthesis from l-arginine, and it exists in three isoforms: endothelial (eNOS), neuronal, and inducible NOS, in which eNOS plays a key role in producing NO in the endothelium. Pro-inflammatory factors, such as 25-hydroxycholesterol,6 oxidized low-density lipoproteins,7,8 tumor necrosis factors,9 and C-reactive proteins,10 decrease the expression and/or activity of eNOS, thereby decreasing NO bioavailability and increasing the risk of subsequent atherosclerotic events.
Arginase (Arg) is a key enzyme that catabolizes l-arginine to urea and l-ornithine (
Fig. 1A
), and it consists of two subtypes: Arg I and Arg II.11 They are encoded by distinct genes, and they share 100% homology regarding enzymatic activities despite sharing approximately 60% similarity in amino acid sequence in total. Arg can directly compete with eNOS through their common substrate l-arginine, thereby reducing NO production. Although Arg I knockout mice die soon after birth due to hyperammonemia,12 Arg II knockout mice exhibit alleviated atheromatous plaque burden, decreased oxidative stress, and restored endothelial diastolic function, all of which are attributable to increased NO production.13 Because Arg II appears to be the predominant isoform in the human endothelium, inhibition of Arg II activity is considered to be a potential therapeutic strategy for PAD.
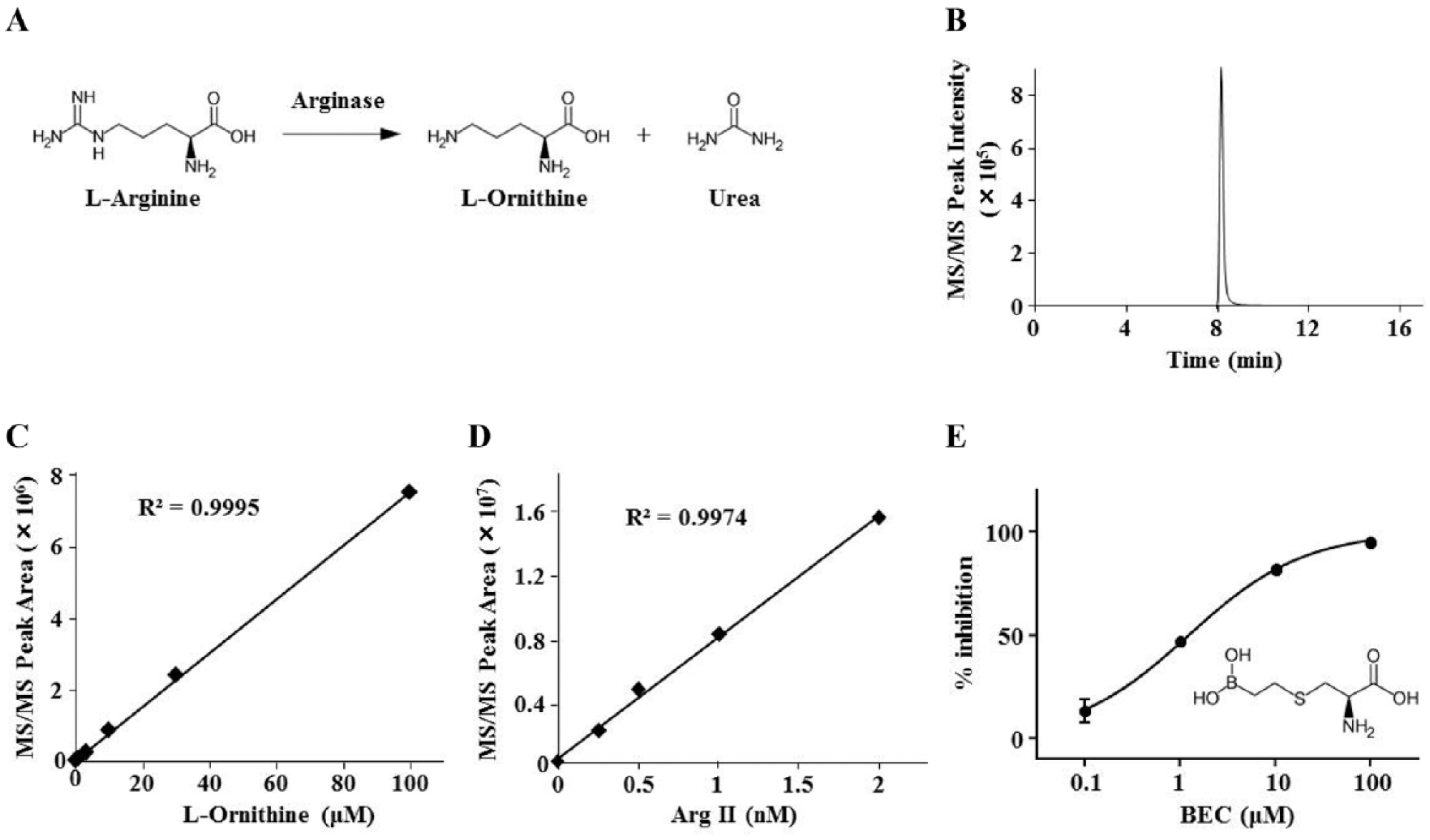
Detection of recombinant human Arg II activity via LC/MS-MS. (A) The enzyme reaction scheme of Arg. (B) A representative LC/MS-MS chromatogram of purified 100 µM l-ornithine eluted via a linear gradient of acetonitrile (from 90% to 40%) for 10 min. (C,D) Quantitative analysis of the analyte peak area for MS/MS signals and confirmation of the linearity and dynamic range for concentration-dependent purified standard l-ornithine (0, 1, 3, 10, 30, and 100 µM) (C) and Arg II enzyme reactions (0, 0.25, 0.5, 1, and 2 nM) for 30 min under the condition of 6.3 mM (1× KM) l-arginine (D). (E) Concentration-dependent inhibition of Arg II activity by BEC (0.1, 1, 10, and 100 µM) under the assay conditions of 1 nM Arg II and 6.3 mM (1× KM) l-arginine and the structure of BEC.
Well-known Arg inhibitors, including l-arginine analogs, lack isoform selectivity.14 Although Arg activity inhibition is suggested to be beneficial in humans, there are potential limitations of long-term Arg I inhibition, considering its essential role in ammonia detoxification in the liver and the involvement of Arg I-positive macrophages in tissue repair.11 We therefore attempted to screen Arg II-selective druggable compounds using a colorimetric biochemical assay;15 however, we failed to identify such compounds from our original chemical libraries, probably because of the striking similarity of metal clusters and active site configurations between the two isoforms of Arg.14 Therefore, to develop Arg II-selective drugs, it is imperative to discover molecules that target the allosteric sites of Arg II. As smaller molecules can enter the small cavities of certain proteins, the chances of discovering Arg II allosteric inhibitors from fragment libraries are high. In this study, we developed a high-throughput screening system using RapidFire mass spectrometry (RF-MS; Agilent Technologies, Lexington, MA) combined with hydrophilic interaction chromatography (HILIC) to assess the activity of Arg II through the detection of l-ornithine, a product of Arg II. By using this method, we conducted a high-throughput, high-concentration screening. As a result, we identified a noncompetitive fragment inhibitor of Arg II. Direct binding of this fragment to Arg II was further validated using a surface plasmon resonance (SPR) assay. We hope that the inhibitor will be developed as a potential drug for treating PAD, and our method for measuring the activity of Arg II can contribute to the identification of other new types of Arg inhibitors.
Materials and Methods
Cloning, Expression, and Purification of Human Arg II
A full-length human Arg II was cloned from Flexi Clone pFN21AB5147 (Promega Corporation, Madison, WI) into a pET15b vector (Novagen, Billerica, MA) with a His8 tag at the N terminus. Plasmids encoding His8-tagged Arg II were introduced into Escherichia coli strain BL21 (DE3). The cells were grown at 37 °C to an optical density (absorbance at 620 nm) of 0.6 and incubated for 5 h at 37 °C after the addition of 0.5 mM isopropyl-β-thiogalactopyranoside. The cells were then harvested, resuspended in lysis buffer (20 mM Tris-HCl buffer [pH 8.0], 150 mM NaCl, 1% CHAPS, 10% glycerol, 40 mM imidazole, 0.5 mM EDTA, and 0.2 mg/mL lysozyme), and disrupted via sonication. After centrifugation, the supernatant was loaded onto an affinity column containing Ni-NTA Superflow resin (QIAGEN, Venlo, the Netherlands), washed with wash buffer (20 mM Tris-HCl buffer [pH 8.0], 150 mM NaCl, 1% CHAPS, 10% glycerol, and 40 mM imidazole), and eluted with elution buffer (20 mM Tris-HCl buffer [pH 8.0], 150 mM NaCl, 1% CHAPS, 10% glycerol, and 150 mM imidazole). The protein was applied to a HiLoad 26/60 Superdex 200-pg column (GE Healthcare, Buckinghamshire, UK), equipped with an AKTA Explorer 10S system (GE Healthcare) equilibrated at 4 °C with equilibration buffer (20 mmol/L Tris-HCl [pH 8.0], 150 mmol/L NaCl, 0.1% CHAPS, and 5% glycerol). Homogeneity of the purified protein was confirmed via sodium dodecyl sulfate–polyacrylamide gel electrophoresis. The purified protein was incubated for 10 min at 60 °C with 10 mM MnCl2 to activate the enzyme,16 aliquoted, and then stored at −80 °C.
Measurement of Arg II Activity via Liquid Chromatography–Tandem Mass Spectrometry
The activity of Arg II was examined using 96-well V-bottom polypropylene plates with a final reaction volume of 80 µL. The reaction mixture containing 50 mM Tris-HCl (pH 7.5), 130 mM NaCl, 0.5 mM MnCl2, 0.01% Tween 20, 6.3 mM (1× KM) l-arginine, and 2 nM purified Arg II (unless otherwise stated) was incubated for 30 min at room temperature. The KM value was predetermined by colorimetric biochemical assay.15 The reaction was stopped by the addition of 80 µL of acetonitrile containing 10 mM ammonium acetate and 20 µM 13C l-ornithine, which was used as an internal control to monitor and guarantee assay quality.
For liquid chromatography–tandem mass spectrometry (LC/MS-MS) analysis, 10 µL of the enzyme reaction solution was loaded onto a ZIC-pHILIC column (Merck KGaA, Burlington, MA) and washed with 10:90 (v/v) water–acetonitrile containing 10 mM ammonium acetate for 2 min. The Arg II product l-ornithine was eluted for 10 min using a linear gradient of water–acetonitrile (10:90, v/v, to 60:40, v/v) containing 10 mM ammonium acetate and analyzed using a 3200QTRAP quadrupole mass spectrometer (Sciex, Framingham, MA) in the positive electrospray ionization mode. Multiple reaction monitoring (MRM) of l-ornithine and 13C l-ornithine was performed at m/z 133 (Q1)/116 (Q3) and m/z 138 (Q1)/121 (Q3), respectively.
Measurement of Arg II Activity via RF-MS
We examined the enzyme activity using 384-well V-bottom polypropylene plates with a final reaction volume of 10 µL. The reaction mixture containing 50 mM Tris-HCl (pH 7.5), 130 mM NaCl, 0.5 mM MnCl2, 0.01% Tween 20, 6.3 mM (1× KM) l-arginine, and 2 nM purified Arg II (unless otherwise stated) was incubated for 30 min at room temperature. The reaction was stopped by the addition of 80 µL of 10:90 (v/v) water–acetonitrile containing 10 mM ammonium acetate and 20 µM 13C l-ornithine.
For RF-MS analyses, the enzyme reaction solution was aspirated from each well for 600 ms and then directly loaded onto a Type H1 (HILIC) cartridge (Agilent Technologies) with 10:90 (v/v) water–acetonitrile containing 10 mM ammonium acetate for 6000 ms. l-Ornithine was eluted using 60:40 (v/v) water–acetonitrile containing 10 mM ammonium acetate for 12,000 ms and analyzed using a 3200QTRAP as described for the LC/MS-MS assay method. The cartridge was changed approximately every 1400 data points.
High-Concentration Fragment Screening
From our original fragment library consisting of approximately 18,000 compounds with a molecular weight of 100–300 Da, we performed high-concentration screening at a final concentration of 250 µM for each compound. The enzyme reaction conditions were as described above (see Measurement of Arg II Activity via RF-MS). Purified Arg II was dispensed into plates as a 4 µL solution using a Multidrop Combi (Thermo Fisher Scientific, Waltham, MA), followed by the addition of 3 µL of the test compounds using an HT Station 1200 (MSTechnos, Tokyo, Japan). The reactions were initiated by adding 3 µL of the Arg II substrate l-arginine, followed by incubation for 30 min at room temperature. After the reaction was stopped and diluted 9-fold by the addition of 80 µL of 10 mM ammonium acetate, 2 µL of the sample solutions was transferred to 384-well V-bottom polypropylene plates and diluted with 78 µL of 10:90 (v/v) water–acetonitrile containing 10 mM ammonium acetate, resulting in a 360-fold dilution of the reaction solutions. 13C l-ornithine was not added to the solutions since the internal standardization negatively affected Z′-factor values. RF-MS analysis was performed as described above.
SPR-Based Binding Assay
All experiments were performed using a Biacore 4000 instrument (GE Healthcare) at 25 °C. His8-tagged Arg II was immobilized onto a Series S Sensor Chip NTA (GE Healthcare) using the capture coupling method, which results in capturing of His8-tagged Arg II in a nonrandom orientation by the His8 tag prior to amine coupling.17 Prior to His8-tagged Arg II capture, the sensor chip was activated using 0.1 M N-ethyl-N′-(dimethylaminopropyl) carbodiimide and 0.4 M N-hydroxysuccinimide, which enabled His8-tagged Arg II to be covalently immobilized to the chip surface. Test compounds diluted with running buffer (50 mM HEPES-KOH [pH 7.4], 150 mM KCl, 0.5 mM MnCl2, and 0.01% Tween 20) were injected at a flow rate of 30 µL/min. For the competitive binding assay, test compounds were diluted with running buffer containing an excess amount of S-(2-boronoethyl)-l-cysteine (BEC) (more than 10× dissociation constant [KD] concentration) so that the binding pocket was almost fully occupied. The running buffer was used as a blank buffer and injected both before and after all concentration series of test compounds. Acquired sensorgrams were double referenced against an intact surface and the average of two blank injections.
Data Analysis
Analyst software (Sciex) or RapidFire Integrator software (Agilent Technologies) was used to quantify the peak area of each well. Assay validation was assessed using the Z′ factor calculated by
where ctrl is the 100% enzymatic activity control, blank is the 0% enzymatic activity control, and SD is the standard deviation.
For the mechanism of action analysis, enzyme kinetic data were fitted to the Michaelis–Menten kinetic model using
where v is the reaction velocity at a substrate concentration [S], Vmax is the maximum velocity at the saturating substrate concentration, [I] is the concentration of the inhibitor, KM is the Michaelis constant, and Ki is the inhibition constant. The mode of inhibition was determined on the basis of the value of α using GraphPad Prism software (GraphPad Software, San Diego, CA). Data are presented as the mean ± standard error of experiments performed in triplicate unless otherwise stated.
SPR data for inhibitor binding (5 s after fragment injection or 55 s after BEC injection) were analyzed using BIAevaluation software (GE Healthcare). Global fitting of multiple concentrations data was performed using a standard reversible 1:1 Langmuir interaction model. KD and the experimental maximal binding response (Rmax) were determined via affinity analysis using analytical software.
The theoretical Rmax of each test compound was calculated using
where theoretical Rmax [RU] is the saturated binding response of the compound, Rprotein is the immobilization level of protein [RU], and Mwcomp and Mwprotein are the molecular weights of the compound and protein, respectively.
Results
LC/MS-MS Assay Development
MS is considered suitable for screening fragment compounds that require high compound concentrations, as it can avoid any influence of detection interference caused by colored or autofluorescent compounds.18 To evaluate Arg II activity (
Fig. 1A
), we decided to directly measure l-ornithine levels via LC-MS detection with a HILIC cartridge, as l-ornithine is a hydrophilic amino acid and it should be separated using a cartridge that can effectively separate highly polar molecules.19 First, we determined the MRM condition in a triple quadrupole tandem mass spectrometry (MS/MS) instrument for l-ornithine at m/z 133/166 using the positive ion mode. After optimization of the condition, commercially available purified l-ornithine was applied to a ZIC-pHILIC column and eluted as a single peak after approximately 8 min using a linear gradient of 90%–40% acetonitrile (
Fig. 1B
). We evaluated the peak area, obtaining concentration-dependent signals over a linear range of at least 1–100 µM (
Fig. 1C
), indicating that the method can quantitatively detect l-ornithine. We thereafter examined whether l-ornithine produced by Arg II could be detected.
Figure 1D
shows that l-ornithine signals were linear for Arg II concentrations ranging from 0.25 to 2 nM. Moreover, we investigated the inhibitory effect of BEC, a substrate analogous inhibitor of Arg I and Arg II, obtaining an IC50 of 1.1 µM (
Fig. 1E
), which was similar to that in previous reports.20 It should be noted that the l-arginine peak was tailed under the current measurement condition. Thus, we chose to quantify only the product rather than substrate/product ratios to represent Arg II activities. These results clearly indicate that our LC-MS conditions can precisely detect the enzymatic activities of Arg II.
RF-MS Assay Development
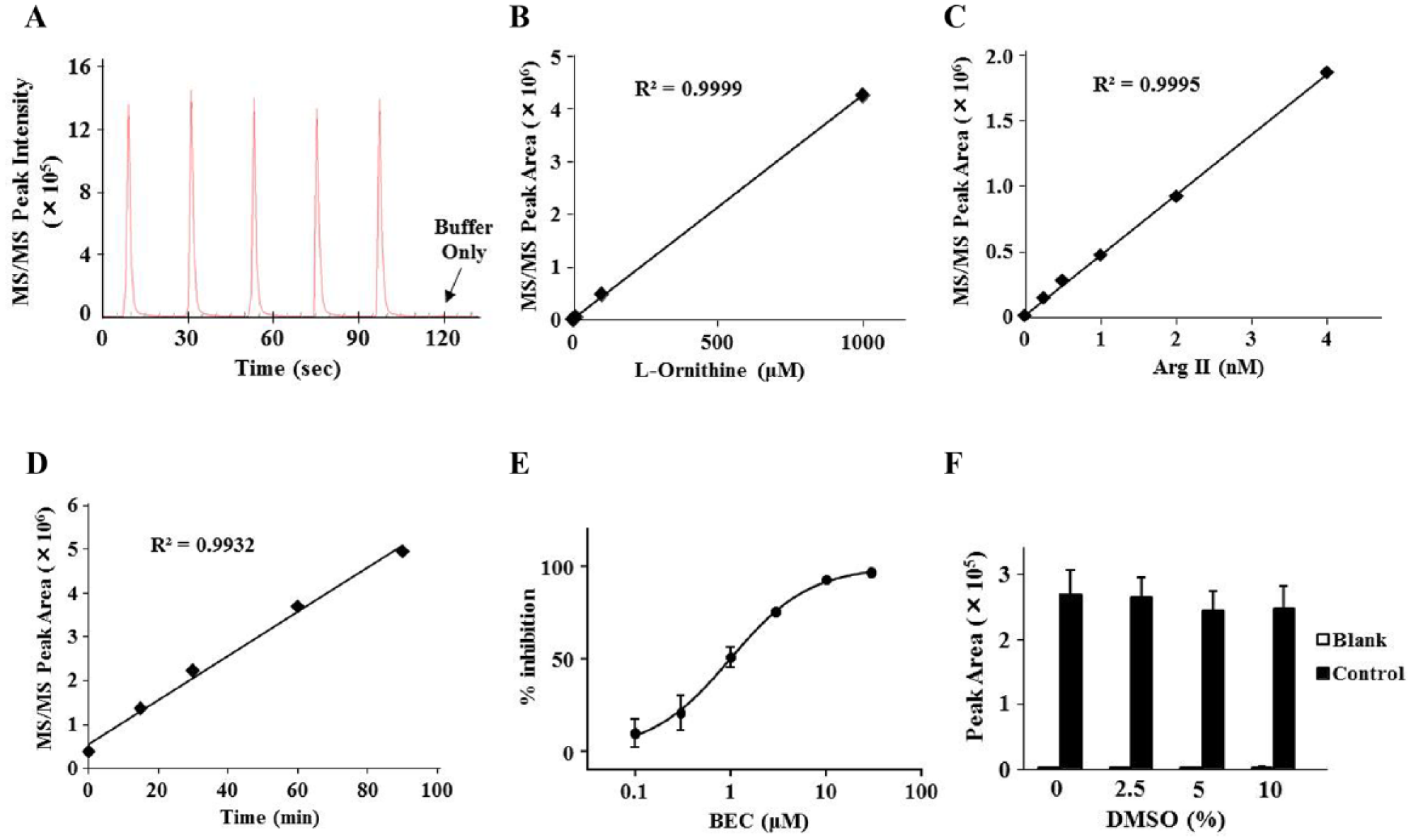
Based on the LC-MS method, we next developed an RF-MS assay using a solid-phase HILIC cartridge (Type H1) for high-throughput analysis. Standard l-ornithine was eluted from the cartridge in an isocratic phase of 40% acetonitrile, and no significant sample carryover was observed (
Fig. 2A
). We evaluated the sample peak area and found that l-ornithine can be quantitatively detected from at least 1 µM (
Fig. 2B
). Then, we confirmed enzyme concentration- and reaction time-dependent signal increases in a linear manner (
Fig. 2C
,
D
). We also confirmed the inhibitory effect of BEC on Arg II with an IC50 of 0.96 µM (
Fig. 2E
), which was nearly equal to that determined using LC/MS-MS. For assay optimization, we performed a DMSO tolerance test and found that the signals were not significantly affected by the addition of up to 10% DMSO (
Fig. 2F
). These results suggest that our RF-MS assay can quantitatively detect the activities of Arg II as well as its inhibitors.
Detection of recombinant human Arg II activity by RF-MS. (A) A representative peak shape of purified 100 µM l-ornithine eluted via an isocratic phase of 40% acetonitrile. (B–D) Quantitative analysis of the analyte peak area for MS/MS signals and confirmation of the linearity and dynamic range for concentration-dependent purified standard l-ornithine (0, 1, 10, 100, and 1000 µM) (B) and concentration-dependent (0, 0.25, 0.5, 1, 2, and 4 nM for 30 min) (C) and time-dependent (0, 15, 30, 60, and 90 min in 2 nM Arg II) (D) enzyme reactions under the condition of 6.3 mM (1× KM) l-arginine. (E) Concentration-dependent inhibition of Arg II activity by BEC (0.1, 0.3, 1, 3, 10, and 30 µM) under the standard assay conditions. (F) Confirmation of the DMSO tolerance of Arg II activity (0%, 2.5%, 5%, and 10%) under the standard assay conditions.
Fragment Library Screening and Hit Identification
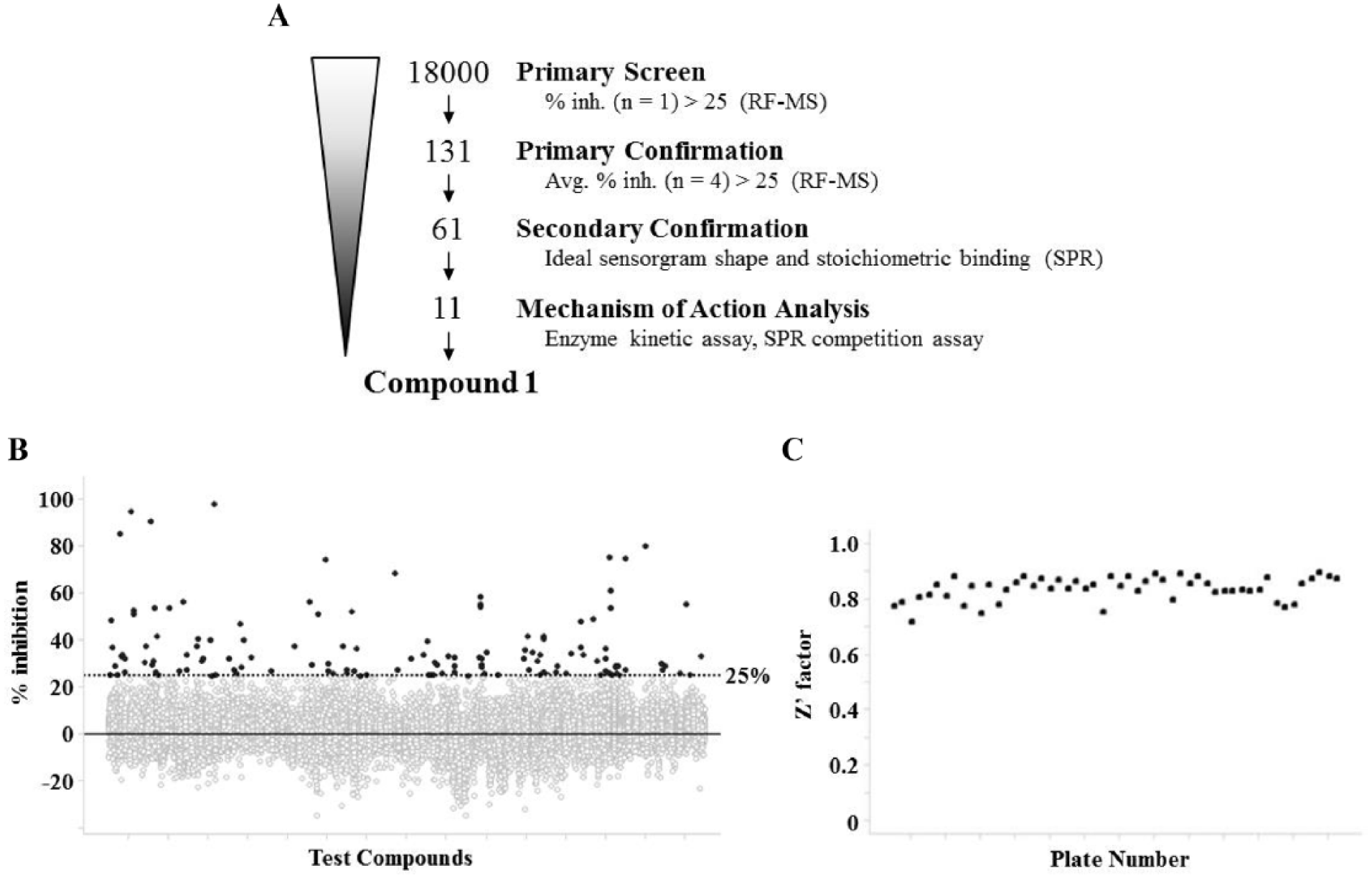
Our screening campaign was conducted as described in
Figure 3A
. Prior to the high-concentration fragment screen, we checked the assay quality of several consecutive 384-well plates, confirming Z′ factor >0.7 and percent of coefficient variation <7 for each plate, indicating excellent assay robustness. Because normalization by internal standard (13C l-ornithine) adversely affected Z′-factor values, we chose to only evaluate product peak area. We then conducted fragment screens on 18,000 small compounds at a single concentration of 250 µM in 10% DMSO via RF-MS (
Fig. 3B
), for which BEC was used as a positive control. The Z′ factor of the entire screening campaign was 0.77 ± 0.11 (
Fig. 3C
). After reproducibility testing in quadruplicate, we chose 61 compounds with >25% inhibitory activity, which was a robust statistical cutoff (
Fig. 3A
,
B
). These results indicate that the screening performance was sufficient for selecting Arg II fragment inhibitors.
High-concentration fragment screening via RF-MS. (A) Flowchart and summary of the screening campaign. (B) Scatterplots of primary screening data and selection of hit compounds exhibiting >25% inhibition activity of purified Arg II at 250 µM. (C) Z′-factor values of the overall primary screening.
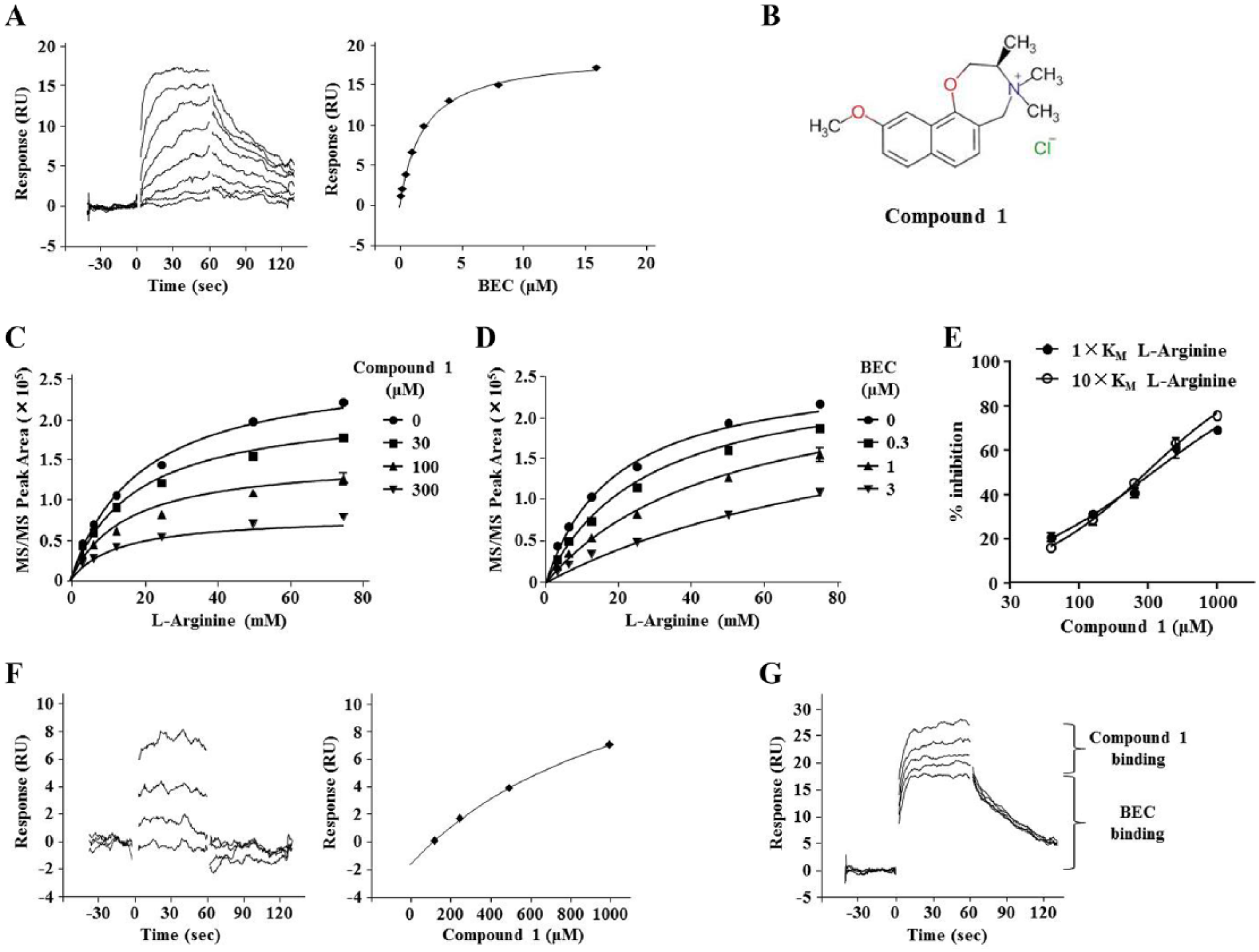
We subsequently performed compound concentration-dependent SPR analysis using a Biacore 4000 to assess the binding capacity and specificity of Arg II. His8-tagged Arg II was captured and immobilized on a Ni-NTA sensor chip, and BEC was confirmed to bind to the protein at a KD of 1.9 µM (
Fig. 4A
), which is nearly consistent with the reported value for binding to Arg I,21 suggesting that the assay system functions suitably. Of the 61 hit compounds, we selected 11 compounds with square-shaped sensorgrams and stoichiometric binding curves, which indicate specific interactions of the compounds with Arg II. We further performed compound concentration-dependent inhibition analysis and investigated the mode of inhibition of the compounds with different concentrations of l-arginine. Consequently, we identified compound 1 (
Fig. 4B
), which had a Ki of 241 µM, versus 0.71 µM for BEC. The compound displayed a noncompetitive inhibition pattern via a nonlinear regression model with Michaelis–Menten kinetics and an α value of 0.53 (
Fig. 4C
), whereas BEC exhibited a competitive inhibition pattern with an α value of 24.5 (
Fig. 4D
). Indeed, the IC50 of compound 1 at a 1× KM substrate concentration was similar to that at 10× KM (
Fig. 4E
). Furthermore, compound 1 had an ideal square-shaped sensorgram, and its experimental Rmax calculated via affinity analysis was 38% of theoretical Rmax (
Fig. 4F
), which is nearly consistent with that of BEC (47% of theoretical Rmax), suggesting that compound 1 binds to Arg II at a 1:1 ratio. Lastly, we performed a competition assay with SPR analysis, finding that compound 1 could bind to Arg II even in the presence of excess amounts of BEC, indicating that the Arg II substrate-binding pocket was fully occupied (
Fig. 4G
). From these results, we concluded that compound 1 is a novel type of Arg II inhibitor that could bind to its potential allosteric site.
Identification and characterization of compound 1. (A) Left: A representative binding sensorgram of BEC (0.032, 0.063, 0.125, 0.25, 0.5, 1, 2, and 4 µM) to Arg II. Right: The steady-state fitting of the equilibrium responses versus compound concentrations based on the 1:1 binding model. (B) Chemical structure of compound 1. (C,D) Nonlinear regression analysis using GraphPad Prism to determine the Ki and α values (the mode of inhibition) of compound 1 (C) and BEC (D). (E) Concentration-dependent inhibition of Arg II activity by compound 1 under the condition of 6.3 mM (1× KM) or 63 mM (10× KM) l-arginine. (F) Left: A representative binding sensorgram displaying the binding of compound 1 (125, 250, 500, and 1000 µM) to Arg II. Right: The steady-state fitting of the equilibrium responses versus compound concentrations based on the 1:1 binding model. (G) A representative binding sensorgram displaying the binding of compound 1 (125, 250, 500, and 1000 µM) to Arg II in the presence of 16 µM BEC.
Discussion
PAD progression is often accompanied by severe gait disorder and cardiovascular events. From the standpoints of quality of life and mortality, prevention and treatment of the disease have been crucial issues worldwide.5 Decreases in NO production are considered a hallmark of endothelial dysfunction and an inducer of PAD, and Arg II is one such candidate that contributes to NO production by competing with eNOS.11 Although their efficacy in vivo has been evidenced by knockout mice experiments,13 selective Arg II inhibitors have not been reported, presumably due to the high similarity between Arg I and Arg II, especially at the active site where l-arginine approaches.14 Therefore, we postulated that the discovery of compounds displaying noncompetitive inhibition would be desirable to develop selective Arg II inhibitors. Furthermore, we assumed that a fragment compound screen would be suitable because no potential allosteric sites that can modulate Arg II activity or allosteric cavities to which small molecules may bind have been clarified. In addition, currently known substrate analog inhibitors do not possess ring structures, which are generally considered one of the ideal drug structures; thus, we assumed that any compounds with novel backbone structures might be appreciated.
In this study, we established a novel system for assessing Arg II activity using an RF-MS system, which enables automatic rapid sample separation via on-line solid-phase extraction.22 Because l-ornithine detection by HILIC appeared not to be standardized, we preliminarily used LC-MS, which enables high-resolution separation of the sample, and confirmed assay conditions. Direct measurement of enzymatic products can be an ideal method in terms of avoiding the interference associated with absorbance or fluorescence outputs, thereby enabling robust assays such as testing a mixture or high concentration of compounds. Significantly, although several high-throughput screens have been attempted using RF-MS,23–26 reports on the high-throughput measurement of hydrophilic molecules in the normal-phase mode are lacking. We believe that our results would further expand the application of RF-MS, thus promoting high-throughput screening, especially when label-free or high-concentration compounds are required.
By conducting a high-concentration screen using a fragment compound library, we identified compound 1 as a noncompetitive Arg II inhibitor, which was revealed by nonlinear regression analysis. Lineweaver–Burk plots further supported this mode of inhibition (
Suppl. Fig. S1
). Importantly, there are no small molecules reported to date that can directly regulate Arg II activity, excluding its substrate analogs (amino acid derivatives). Of note, no hit compounds were discovered from our original compound library for high-throughput screening that was developed according to Lipinski’s rule of five, implying that the potential allosteric site is not a typical ligandable pocket and the substrate-binding site is not sufficiently large enough for our high-throughput screening library to bind with high affinity.
From the SPR data, we believe that compound 1 is a specific allosteric inhibitor of Arg II, including the reasonable experimental Rmax and sensorgram. However, the KD of Arg I was 1.1 mM (
Suppl. Fig. S2
), meaning that the compound did not exhibit selectivity against Arg I. The reason for this is that, in addition to it being difficult to achieve selectivity of fragment compounds,27,28 Arg I and Arg II exhibit 100% homology in the responsible region of enzymatic activities, suggesting that it may be possible that the allosteric binding sites of compound 1 also resemble each other. However, given that they only exhibit approximately 60% similarity of total amino acid sequences, we expect that selectivity might be obtained via structure analysis of the Arg II–compound 1 complex, as well as investigation of the structure–activity relationship of compound 1 with the derivatives.
In summary, we developed a new method for directly measuring Arg II activity using a HILIC cartridge in a high-throughput manner. To the best of our knowledge, this is the first report to separate hydrophilic molecules through the normal-phase mode of chromatography in combination with RF-MS technology, which we believe may contribute to the expansion of its applications. Using the method, we performed high-concentration screens and identified compound 1 as a noncompetitive fragment inhibitor, which was validated by our developed SPR assay. Although the inhibition activity of compound 1 is weak and requires further development before animal testing and subsequent clinical studies, we hope that this approach will lead to the development of novel drugs for treating PAD.
Supplemental Material
20181107_Hantani_Supplemental_Material_revised – Supplemental material for Identification of an Arginase II Inhibitor via RapidFire Mass Spectrometry Combined with Hydrophilic Interaction Chromatography
Supplemental material, 20181107_Hantani_Supplemental_Material_revised for Identification of an Arginase II Inhibitor via RapidFire Mass Spectrometry Combined with Hydrophilic Interaction Chromatography by Wataru Asano, Yu Takahashi, Motoaki Kawano and Yoshiji Hantani in SLAS Discovery
Footnotes
Acknowledgements
We thank Drs. Hideo Tanaka and Shohei Oie for helpful discussions. We also thank Mr. Kenji Yamanaka for technical support.
Supplemental material is available online with this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
References
1.
JoostenM. M.PaiJ. K.BertoiaM. L.et al. Associations between Conventional Cardiovascular Risk Factors and Risk of Peripheral Artery Disease in Men. JAMA2012, 308, 1660–1667.
2.
GBD 2015 Disease and Injury Incidence and Prevalence Collaborators. Global, Regional, and National Incidence, Prevalence, and Years Lived with Disability for 310 Diseases and Injuries, 1990–2015: A Systematic Analysis for the Global Burden of Disease Study 2015. Lancet2016, 388, 1545–1602.
ChiY. W.LavieC. J.MilaniR. V.et al. Safety and Efficacy of Cilostazol in the Management of Intermittent Claudication. Vasc. Health Risk Manage. 2008, 4, 1197–1203.
OuZ. J.ChenJ.DaiW. P.et al. 25-Hydroxycholesterol Impairs Endothelial Function and Vasodilation by Uncoupling and Inhibiting Endothelial Nitric Oxide Synthase. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E781–E790.
7.
LiaoJ. K.ShinW. S.LeeW. Y.et al. Oxidized Low-Density Lipoprotein Decreases the Expression of Endothelial Nitric Oxide Synthase. J. Biol. Chem. 1995, 270, 319–324.
8.
ZhouJ.AbidM. D.XiongY.et al. ox-LDL Downregulates eNOS Activity via LOX-1-Mediated Endoplasmic Reticulum Stress. Int. J. Mol. Med. 2013, 32, 1442–1450.
9.
YoshizumiM.PerrellaM. A.BurnettJ. C.Jr.et al. Tumor Necrosis Factor Downregulates an Endothelial Nitric Oxide Synthase mRNA by Shortening Its Half-Life. Circ. Res. 1993, 73, 205–209.
10.
VenugopalS. K.DevarajS.YuhannaI.et al. Demonstration That C-Reactive Protein Decreases eNOS Expression and Bioactivity in human aortic endothelial Cells. Circulation2002, 106, 1439–1441.
11.
CaldwellR. B.ToqueH. A.NarayananS. P.et al. Arginase: An Old Enzyme with New Tricks. Trends Pharmacol. Sci. 2015, 36, 395–405.
12.
IyerR. K.YooP. K.KernR. M.et al. Mouse Model for Human Arginase Deficiency. Mol. Cell. Biol. 2002, 22, 4491–4498.
13.
RyooS.GuptaG.BenjoA.et al. Endothelial Arginase II: A Novel Target for the Treatment of Atherosclerosis. Circ. Res. 2008, 102, 923–932.
14.
SteppanJ.NyhanD.BerkowitzD. E.Development of Novel Arginase Inhibitors for Therapy of Endothelial Dysfunction. Front. Immunol. 2013, 4, 278.
15.
Van ZandtM. C.WhitehouseD. L.GolebiowskiA.et al. Discovery of (R)-2-Amino-6-Borono-2-(2-(Piperidin-1-yl)ethyl)Hexanoic Acid and Congeners as Highly Potent Inhibitors of Human Arginases I and II for Treatment of Myocardial Reperfusion Injury. J. Med. Chem. 2013, 56, 2568–2580.
16.
ColleluoriD. M.MorrisS. M.Jr.AshD. E.Expression, Purification, and Characterization of Human Type II Arginase. Arch. Biochem. Biophys. 2001, 389, 135–143.
17.
KimpleA. J.MullerR. E.SiderovskiD. P.et al. A Capture Coupling Method for the Covalent Immobilization of Hexahistidine Tagged Proteins for Surface Plasmon Resonance. Methods Mol. Biol. 2010, 627, 91–100.
18.
LeveridgeM. V.BarderaA. I.LaMarrW.et al. Lead Discovery for Microsomal Prostaglandin E Synthase Using a Combination of High-Throughput Fluorescent-Based Assays and RapidFire Mass Spectrometry. J. Biomol. Screen. 2012, 17, 641–650.
KimN. N.CoxJ. D.BaggioR. F.et al. Probing Erectile Function: S-(2-Boronoethyl)-l-Cysteine Binds to Arginase as a Transition State Analogue and Enhances Smooth Muscle Relaxation in Human Penile Corpus Cavernosum. Biochemistry2001, 40, 2678–2688.
21.
Di CostanzoL.IliesM.ThornK. J.et al. Inhibition of Human Arginase I by Substrate and Product Analogues. Arch. Biochem. Biophys. 2010, 496, 101–108.
22.
JonasM.LaMarrW. A.OzbalC.Mass Spectrometry in High-Throughput Screening: A Case Study on Acetyl-Coenzyme A Carboxylase Using RapidFire–Mass Spectrometry (RF-MS). Combin. Chem. High Throughput Screen. 2009, 12,752–759.
23.
HoltT. G.ChoiB. K.GeoghagenN. S.et al. Label-Free High-Throughput Screening via Mass Spectrometry: A Single Cystathionine Quantitative Method for Multiple Applications. Assay Drug Dev. Technol. 2009, 7, 495–506.
24.
PlantM.DineenT.ChengA.et al. Screening for Lysine-Specific Demethylase-1 Inhibitors Using a Label-Free High-Throughput Mass Spectrometry Assay. Anal. Biochem. 2011, 419, 217–227.
25.
LeveridgeM.CollierL.EdgeC.et al. A High-Throughput Screen to Identify LRRK2 Kinase Inhibitors for the Treatment of Parkinson’s Disease Using RapidFire Mass Spectrometry. J. Biomol. Screen. 2016, 21, 145–155.
26.
AdachiR.IshiiT.MatsumotoS.et al. Discovery of Human Intestinal MGAT Inhibitors Using High-Throughput Mass Spectrometry. SLAS Discov. 2017, 22, 360–365.
27.
ChenY.ShoichetB. K.Molecular Docking and Ligand Specificity in Fragment-Based Inhibitor Discovery. Nat. Chem. Biol. 2009, 5, 358–364.
28.
ShipeW. D.SharikS. S.BarrowJ. C.et al. Discovery and Optimization of a Series of Pyrimidine-Based Phosphodiesterase 10A (PDE10A) Inhibitors through Fragment Screening, Structure-Based Design, and Parallel Synthesis. J. Med. Chem. 2015, 58, 7888–7894.
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.