
Abstract
Working memory refers to the cognitive capacity to temporarily store and manipulate information from multiple sensory domains. Recent studies have shown that cognitive training can improve performance in both visual working memory and tactile working memory tasks. However, it is still unclear whether the effects of training can be transferred from one sensory modality to another. The current study assessed whether the training effect of the tactile orientation sequence task could transfer to visual orientation sequence and visuospatial working memory tasks. The results showed that participants’ accuracy in the tactile orientation sequence task was significantly increased after 9 days of training compared with that before training. Remarkably, participants’ accuracy in both the visual orientation sequence task and the visuospatial task was significantly improved after 9 days of training. These results suggest that it is possible to improve visual working memory through a transfer effect from tactile task training without practice in the visual domain, which opens a wide range of applications for tactile orientation sequence tasks.
Keywords
Introduction
Working memory refers to the cognitive capacity to temporarily store and manipulate information from multiple sensory domains, including but not limited to visual and tactile domains (A. Baddeley, 2012; Constantinidis, 2016; Fassihi et al., 2014). A model of working memory is a multicomponent model that consists of a central executive system and three subsystems, namely, a phonological loop, a visuospatial sketchpad, and an episodic buffer, for maintaining specific types of information (A. Baddeley, 2012; A. D. Baddeley et al., 2019). Working memory deficits have also been identified as a potential cause of multiple cognitive impairments in children and adults (Li et al., 2016; Melby-Lervag & Hulme, 2013; Swanson et al., 2009). Increasing evidence has shown that adults are capable of strong cognitive and brain plasticity that, with a short period of intensive practice via behavioral tasks, can improve working memory ability (Au et al., 2015; Jaeggi et al., 2008; Likova, 2012).
Previous studies have identified numerous behavioral tasks that are effective in improving working memory for both temporal and spatial tasks. Baker et al. (2014) used a temporal sequence task to train participants to learn sequential grating stimuli. In this task, participants were presented a sequence of visual gratings with the goal of acknowledging the concept of temporal sequences. Their results indicated that the performance of the temporal sequence task could be improved by training (Baker et al., 2014). Furthermore, the training effects of spatial tasks were suggested by a representative prior study, in which Corsi boards were used to train preschool children; the study found that visuospatial working memory performance substantially improved over 12 sessions (Gade et al., 2017). Clearly, both temporal and spatial tasks can benefit from training in the visual working memory domain and have a traceable effect on visual working memory processes.
In fact, working memory is not related exclusively to the visual domain. Schmidt and Blankenburg (2018) proposed that human brains can represent mental content in tactile formats by “tactospatial sketchpad” in working memory model. It is assumed that tactile processing can draw upon a modality-specific working memory system to maintain extracted tactile features (Kaas et al., 2007; Katus et al., 2015; Schmidt & Blankenburg, 2018). Previous studies have attempted to find improvements in tactile working memory as a result of behavioral training. A recent study used a novel cognitive-kinesthetic method that trained blind participant to draw complex images by tactile memory alone. The participant was instructed to use left hand to tactually explore a raised-line drawing model in the left section of the drawing tablet and to develop a full representation of the image in her memory to prepare for the next step. Then, the model image was removed, and the right hand was used to draw the image (from tactile memory) in the right section of the tablet in the memory drawing phase. This study indicated that cortical reorganization in V1 could be harnessed for tactile memory training and testing (Likova, 2012). Bonino et al. (2008) also found that both tactile and visuospatial working memory tasks employed similar frontoparietal cortical areas in sighted participants and that blind participants also recruited similar cortical areas for the same tactile task, providing support for the supramodal functional organization of the visual and tactile cortex. Furthermore, Reuter et al. (2014) used event-related potentials (ERPs) to assess differences in learning tactile stimuli, and they found that P300 latencies were reduced in young adults and that the amplitudes were increased in late middle-aged adults after intervention with a temporal discrimination task. These brain physiology results demonstrated that tactile stimulation training could partially reverse age-related cognitive decline among older adults and increase processing speed in younger participants (Reuter et al., 2014). These studies confirmed that there is a modality-specific working memory system for tactile working memory and that training tasks can improve tactile working memory performance.
Working memory training via tactile tasks and the transfer of training effects have important applications for blind participants. Previous studies used tactile or auditory tasks to train the working memory of blind and sighted participants and observed positive training effects (Gudi-Mindermann et al., 2018; Leo et al., 2018; Setti et al., 2018). Spatial information processing across visual and tactile sensory modalities recruited similar frontoparietal networks, specifically in the posterior parietal cortex, the dorsolateral prefrontal cortex, and the anterior cingulate cortex (Bonino et al., 2008). Wang et al. (2015) found that neural networks in the dorsolateral prefrontal cortex may consist of different populations of neurons. These neurons act in sequence during a spatial task, from visual stimulus encoding and cross-modal information transfer between visual and tactile stimuli to tactile action. These findings provide neurobiological support for behavioral observations by indicating that common cerebral regions subserve the generation of higher-order mental representations involved in working memory independently from a specific sensory modality (Bonino et al., 2008; Ricciardi et al., 2006). Although many studies have evaluated the effects of tactile training or visual training in behavioral tasks, it remains unclear whether the performance of tactile training tasks can affect performance in visual and other tasks. Based on the fact that visual and tactile information share common processing and a common neural mechanism in working memory, we hypothesized that if tactile working memory could benefit from training, then the training effects should be transferred to other modalities, such as vision.
Cattaneo and Vecchi (2008) asked participants to memorize a series of locations presented on different matrices and later asked them to reconstruct patterns of the matrices on empty frames. The results of their experiments in visual and tactile modalities indicated that while visual stimuli were processed in a holistic, simultaneous manner, tactile information was acquired and retrieved in a way similar to the sequential method. Furthermore, Baker et al. (2014, 2015) used a visual sequence task to confirm that the processing of sequence tasks could be improved through training in a visual modality. The current study aimed to develop a tactile training task for working memory and attempted to verify that the improved effect could be transferred to a visual modality. Therefore, we redesigned a tactile sequential task that was slightly different from Baker’s study and used it to train participants’ working memory. The benefits of working memory training were expected to show both near-transfer and far-transfer effects. Near-transfer refers to the training effect observed in related working memory processing, which was assessed using similarity evaluation tasks (Melby-Lervag et al., 2016); this study used the tactile orientation sequence task as the training task, whereas the visual orientation sequence task was used to evaluate the training effects from the modality visual to the tactile modality. In both visual and tactile orientation sequence tasks, participants were presented a sample sequence consisting of eight stimuli and were required to memorize both their sequence and temporal order to determine whether it was the same as the test sequence (Baker et al., 2014). As the two tasks were designed similarly, we expected to observe the near-transfer effects evaluated by the visual orientation sequence task. Moreover, far transfer refers to the training effects observed on general cognitive processes, which were assessed using seemingly unrelated evaluation tasks (Melby-Lervag & Hulme, 2013; Melby-Lervag et al., 2016). In the visuospatial task, participants needed to simultaneously memorize multiple color points displayed on a reference screen in terms of their locations and colors and with the color points presented on a probe screen after a short delay. This experiment was designed to follow the typical process of a visual modality (Cattaneo & Vecchi, 2008; Luck & Vogel, 1997). Although the visuospatial task did not require sequential perception, it still shared processing mechanisms with the training task, such as temporary storage and retrieval objectives in visual working memory (Gazzaley & Nobre, 2012). Therefore, it can be speculated that if these common processes were improved by the training task, training effects may be transferred to visuospatial tasks; this outcome, the far-transfer effect, was expected to be observed.
The current study used a tactile orientation sequence task to train participants’ temporal sequence working memory ability. It is presumed that tactile training can improve the performance of working memory for temporal sequence tasks in tactile modality, which can be verified by the improvement of accuracy in tactile orientation sequence tasks through training sessions. Moreover, a near-transfer effect is expected from tactile training to the visual orientation sequence task, which depends on the sequential processing similarity with the training task. The near-transfer effects can be verified by the improved accuracy in the visual orientation sequence task after training. Last, we assumed a far-transfer effect from the tactile orientation sequence task to a visuospatial task, which depends on the general training effects for working memory. The far-transfer effects can be verified by the increase in item capacity in the visuospatial task after training.
Method
Participants
Thirty-two healthy students from Okayama University participated in this experiment, comprising 16 in the training group (12 males, aged between 20 and 28 years, M = 21.94, SD = 1.88) and 16 in the control group (11 males, aged between 21 and 29 years, M = 23.13, SD = 2.45); they were compensated for their participation. The participants were given written consent forms with details of the experiment, which they signed prior to participation. All experimental protocols were approved by the Institutional Review Board of Okayama University.
Experimental Design
We recruited two groups of participants and randomly assigned them to the training group and the control group. Before the main experiment, the working memory levels of all participants were evaluated via a set of cognitive tasks as pretests (a visual orientation sequence task and a visuospatial task). These evaluation tasks were designed according to the studies of Baker et al. (2015) and Luck and Vogel (1997), and we explained the difference between our paradigm and that of previous studies in the methods for each task (see Figures 4 and 5). After these pretests, the training group completed five training sessions for the tactile orientation sequence task with an average interval of 2 days between sessions. The control group was not contacted during this period. After the training or no-contact period of approximately 9 days, both groups performed the working memory tasks (posttest) again with the same procedure as the pretest (see Figure 1).

Experimental design of the current study.
Training Experiment—Tactile Orientation Sequence Task
Apparatus and stimulus
The JVP domes (Johnson et al., 1997; diameter: 25 mm, width: 3 mm, see Figure 2A) were used as tactile stimuli in the tactile training task. The JVP domes were used to measure the spatial acuity of skin surfaces. Participants could report the orientation of the presented stimuli as left or right.
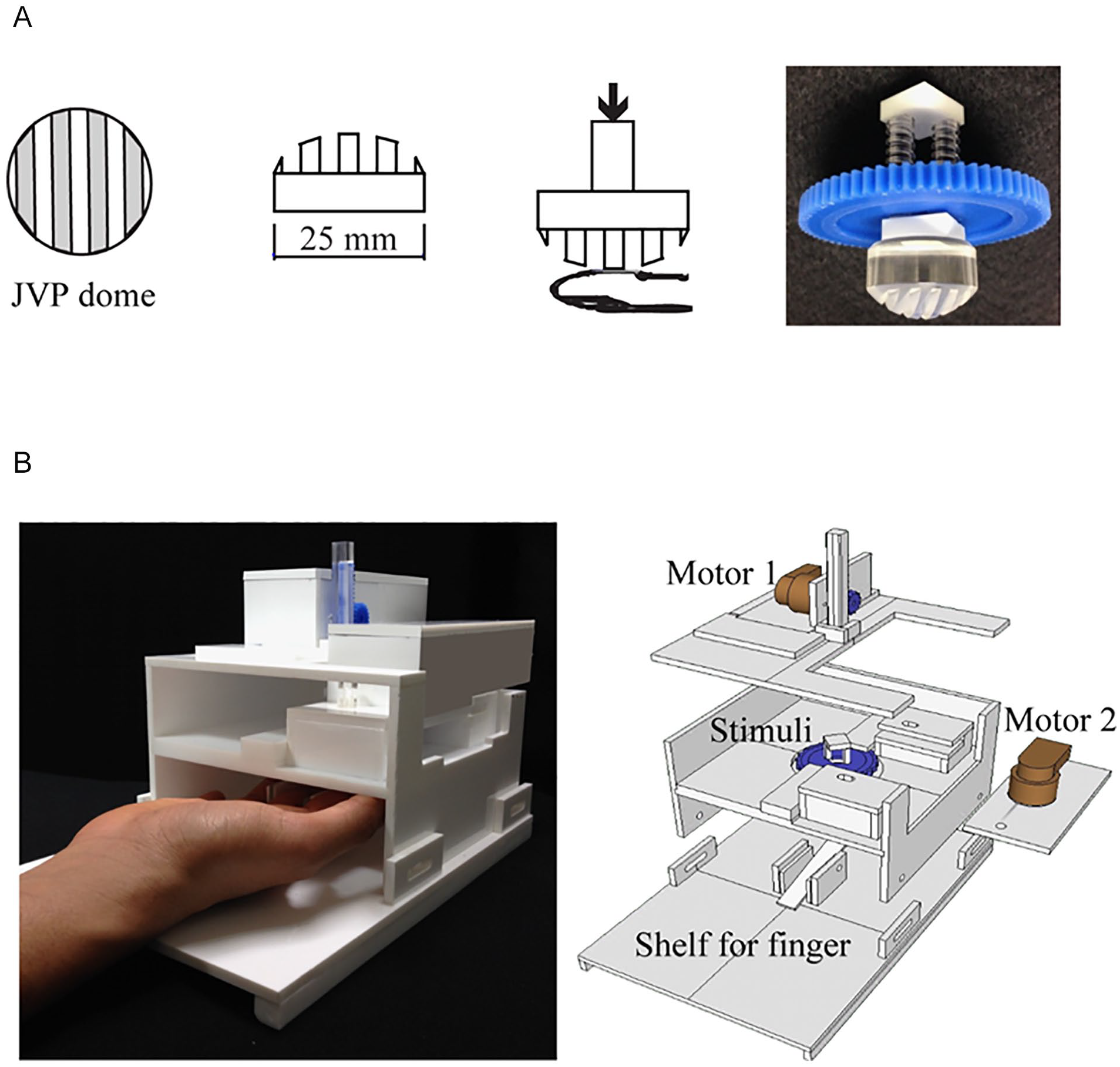
Stimuli and conveyor device for the tactile orientation sequence task: (A) Stimuli for tactile orientation task. JVP domes are small gratings for cutaneous spatial resolution measurement. The gratings are placed on the skin, and participants are required to identify the orientation of the grooves and ridges. (B) Conveyor device for the tactile orientation task. An Application Programming Interface program was used to control the rotation speed and distance of the step motors, ensuring that the stimuli could be applied to the participant’s index finger.
A conveyor device was used to press the grooves of the JVP domes against the participants’ skin in different orientations (120° [left] or 60° [right]). This conveyor device consists of one closed box with an adjustable shelf, two pairs of gears, two step motors, and a group of JVP domes. Almost all of the components were made of acrylic and assembled using resin glue and gears, and the entire device was controlled by a computer. In tactile task, the participants put their index finger on an adjustable stand to ensure that it was held in place in a comfortable position. The JVP dome was installed under a round gear, and a one-step motor was used to rotate it; the other gear and step motor were placed above the round gear to control the upward and downward motion of the stimuli. An API (Application Programming Interface) program was used to control the rotation speed and distance of the step motors, ensuring that the stimuli could be applied to the participant’s index finger under different experimental conditions (see Figure 2B).
The tactile training task was executed in a test room with normal lighting, and participants wore black eye masks throughout the experiment.
Design and procedure
The tactile training task was adapted from Baker et al. (2015). Different from Baker’s visual experiment, we used JVP domes that presented tactile orientation information to replace visual stimuli (Johnson et al., 1997; see Figure 2A). The JVP dome was presented against the index fingertips of the participants and assigned them to report the orientation of each JVP dome. If we define the left orientation as 1 and the right orientation as 2, one example pair of sequences would be as follows: first sequence: 2 1 2 1 1 2 1 2; second sequence: 2 1 2 1 1 2 1 2 or 2 1 2 1 1 2 1 1. Each initial stimulus sequence was composed of four right orientations and four left orientations. Each sequence was repeated twice, resulting in an overall presentation that included 16 stimuli, with a variable target in the second sequence. All orientation stimuli were presented at the same rate. The orientation stimuli were randomly presented in each sequence (see Figure 3). To train working memory capacity and the ability to maintain information, the participants were instructed to memorize a total of eight stimulus presentations in the first sequence and maintain the sequence content in memory until they could match the target items in the second sequence. The participants were required to answer verbally whether the first sequence and the second sequence were the same. In the tactile training task, we simply recorded the accuracy of participants’ answers in that experiment.

Time course of tactile orientation sequence task.
The purpose of the current study is to investigate whether the storage and retrieval ability of working memory can be improved with training, which can be reflected by accuracy. Therefore, the participants were instructed to respond as accurately as possible in all training and evaluation experiments.
Evaluation Experiment—Visual Orientation Sequence Task
Apparatus and stimulus
The parameters and program used in the visual orientation task were designed according to Baker et al. (2015). The visual orientation stimuli, consisting of grayscale sinusoidal gratings, were presented at a 10.8° visual angle. The spatial frequency was 1 cycle per degree across trials, using 100% contrast and a randomized phase. These gratings were rotated ±10° from the vertical orientation (90°), resulting in two different gratings oriented at either 100° (left) or 80° (right). The stimuli were generated and presented by Psychtoolbox-3 (Pelli, 1997) and presented on a 24-inch LCD monitor (ProLite GB2488HSU 1920 × 1,080 pixels, 60 Hz frame rate) at a 75 cm distance. The experiment was conducted in a darkened testing room.
Design and procedure
Each sequence was composed of eight gratings, and each trial had two sequences, as shown in Figure 4. The sequence design was the same as that of the tactile sequence task. For each trial, the participants viewed 16 gratings that were presented sequentially on a gray background at the center of the display. The grating stimuli were presented for 300 ms, followed by a white fixation dot at an interval of 300 ms. The first eight gratings were the sample sequence. The target stimulus appeared only in the test sequence composed of the last eight gratings, and its position was randomized across trials. The target stimuli appeared after a cue (a red dot presented for 1,000 ms). The target stimulus appeared for up to 1,000 ms surrounded by a red circle and was followed by a red fixation point, which disappeared as soon as the participant responded. For each block, 50% of the target stimuli were presented in the same orientation as in the sample sequence, and the other 50% trials featured a different orientation. Participants were instructed to remember the sample sequence and then compare it with target stimuli in the test sequence by pressing the “1” key when the target stimuli had the same orientation (left vs. right) as the grating presented in the sample sequence and pressing the “2” key when the target stimuli were oriented differently from the sample. After the participant’s response, the remaining part of the sequence was presented; a black cross was then presented for 1,000 ms, indicating the end of the sequence and the start of a new trial. Except for the target stimuli, the other gratings in the second sequence were identical to those in the sample sequence.

Time course of the visual orientation sequence task.
There was no feedback in our experiment. In each evaluation task, the participants performed the visual task for two blocks of 40 trials, with at least a 10-min break between blocks. Each participant’s response was recorded to calculate the accuracy in the visual orientation sequence task.
Evaluation Experiment—Visuospatial Task
Apparatus and stimulus
The design of the visuospatial task was based on prior research (Baker et al., 2015; Luck & Vogel, 1997). The stimulus was composed of colored points and was presented within a 9.8°×7.3° region displayed on a black background, and the items in a given array were separated by at least 2.0° (center to center). The experiments shown in Figure 5 used sample arrays consisting of one to 14 colored dots (0.65°), each of which was selected at random from a set of seven highly discriminable colors (red, blue, violet, green, yellow, brown, and white). The colored points were displayed in random locations within a 10 × 10 grid (jittered ± 10 pixels), and each color appeared in the reference display on a maximum of two points.

Time course of the visuospatial task.
Design and procedure
Visuospatial task started with a white fixation point; the display then presented the reference array with two randomly colored points for 200 ms. After a delay of 1,000 ms, only one of the colored points reappeared on the display at the same location as in the reference. The color of the test stimuli was the same as that of the corresponding item in the sample array in 50% of the trials and different in the other 50% trials. The participants were instructed to press the “1” key if the test stimuli were the same color as the reference and press the “2” key if they were different.
In the current visual evaluation task, a two-down, one-up staircase paradigm was used to manipulate the number of colored points in the display. For example, the first three trials of the visuospatial task were set to contain two points in the display as the first step of the staircase. If the participant had two correct responses in these three trials, an additional colored point was added to the display in the next step so that the new, higher step would contain three colored points. If the participant provided two or more incorrect responses in a step, one colored point was removed from the display in the next staircase, reversing the direction of the staircase. Each block was ended after 10 staircase reversals. Thresholds of item capacity in each block (i.e., the number of colored points in the display) were calculated by averaging the last two thirds of the reversals in each block. All participants completed two blocks, after which the threshold of two blocks was computed as their item capacity. In this task, a higher capacity (a greater number of items in the display) denoted better performance.
Data Analysis
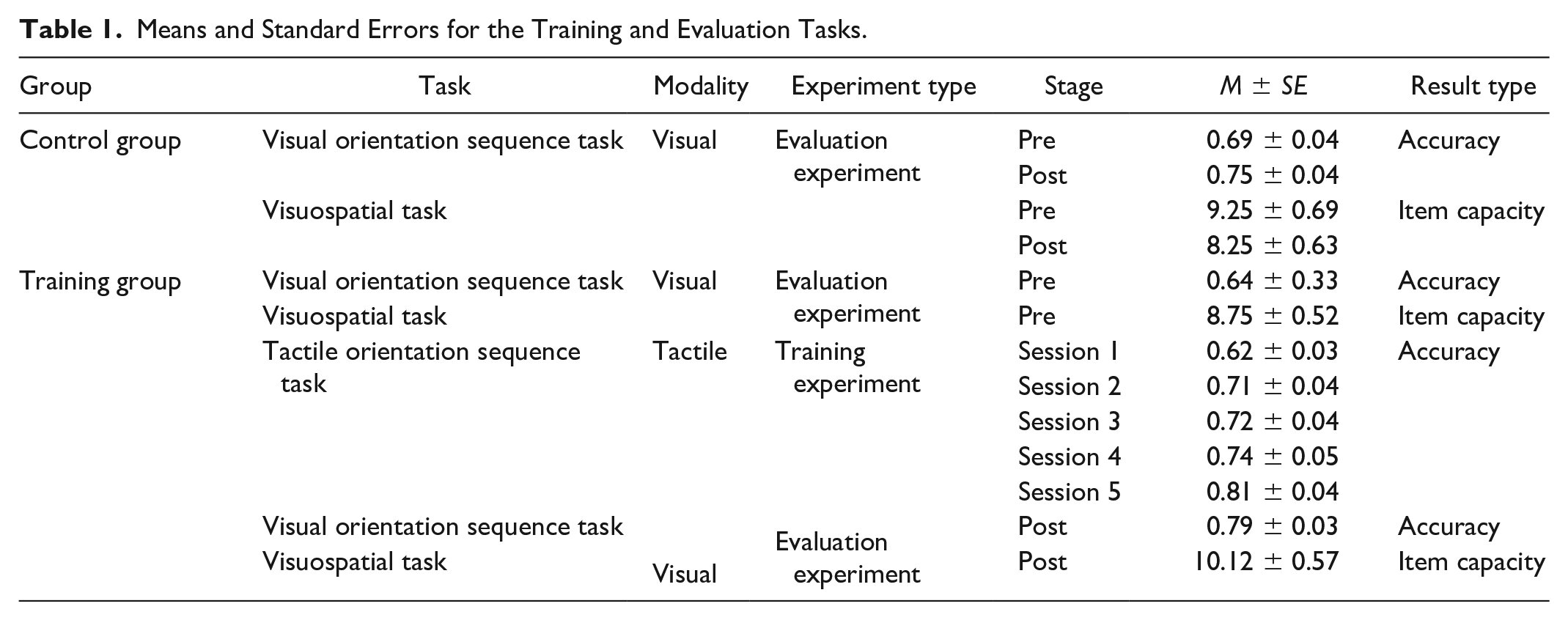
The behavioral performance on the tactile orientation sequence tasks was assessed according to accuracy (the proportion of correctly answered trials). Accuracy was regarded as the measure of participants’ performance for the visual orientation sequence task. Furthermore, item capacity was used as the performance measure for the visuospatial task. The descriptive statistics of the training and evaluation tasks are presented in Table 1. All data were tested for normality using the Shapiro–Wilk test before statistical analysis. Because the accuracy in the training experiment (tactile orientation sequence task) and the item capacity of Evaluation Experiment 2 (visuospatial task) did not fit a normal distribution, nonparametric statistics for the tactile orientation sequence task and visuospatial task were used. The accuracy data in Evaluation Experiment 1 (visual orientation sequence task) fit a normal distribution, so mixed-model analysis of variance (ANOVA) was used for the visual orientation task. The statistical significance threshold was set at p < .05, and Bonferroni correction was applied for all post hoc analyses (Burge et al., 2013; Kowalewski et al., 2012).
Means and Standard Errors for the Training and Evaluation Tasks.
Results
Training Experiment (Tactile Orientation Sequence Task)
The accuracy of each training sessions was calculated and presented in Figure 6. To investigate how accuracy varied across sessions (on Days 1, 3, 5, 7, and 9) for the training group, Friedman ANOVAs was performed, followed by post hoc paired Wilcoxon tests. The results showed a significant training effect in the training group (χ2 = 21.38, p < .001, η2 = 0.344, Kendall’s W = 0.334). The accuracy rates in Sessions 4 and 5 were significantly higher compared with the Session 1 baseline (p = .02 and .016, respectively; see Figure 6). These results indicated an escalating trend of accuracy from Session 1 to Session 5.

Accuracy for the tactile orientation sequence task.
Evaluation Experiment 1 (Visual Orientation Sequence Task)
The mean accuracy on the visual orientation sequence task for the pre- and posttests is shown in Figure 7. To compare the performance between the control group and training group before and after training, the mixed-model ANOVA was used to investigate the effect of stage (pretest vs. posttest) and group (training vs. control). The results revealed a significant main effect of stage, F(1, 30) = 29.94, p < .001, η2 = 0.666; a significant interaction effect was observed in Stage × Group, F(1, 30) = 5.21, p = .041, η2 = 0.251. The main factor group was not significant. Multiple comparisons showed that the training group had significantly improved after training in the tactile task (t = 5.68, p < .001, rYλ = .484), and there was no significant difference between the pre- and posttests in the control group (t = 1.97, p = .069, rYλ = .17). In addition, there was no significant difference in pretest performance between the control and training groups (t = 0.51, p = .618, rYλ = .126).

Accuracy for the visual orientation sequence task.
Evaluation Experiment 2 (Visuospatial Task)
The item capacity of the visuospatial task for the pre- and posttests is shown in Figure 8A. Differences across groups in the pretest were compared by Mann–Whitney tests, and we found no significant difference in the performance of the training and control groups before training (U = 85.5, p = .104, r = .41). Differences within groups between the pre- and posttests were compared using paired Wilcoxon tests. There was no significant difference observed in the control group between the pre- and posttests (p = .181, r = .35). A significant difference between pre- and posttests was found in the training group, indicating that the performance of the visuospatial task was significantly improved after tactile training (p = .039, r = .52). Moreover, the cumulative distribution was used to assess the trend of task difficulty between the pre- and posttests (see Figure 8B). Significant changes between the pre- and posttests were observed in the training group, and this trend did not appear in the control group.

Item capacity and response time for the visuospatial task: (A) Threshold of item capacity in the pre- and posttests. The error bars correspond to ±SE. (B) The frequency of item capacity in each staircase. These results reflect the change in item capacity between the pre- and posttests. The results indicated an improvement after training.
Correlation Between Training and Evaluation Experiment
To further investigate the correlation between the training and evaluation experiments, we correlated behavioral performance before and after training in these tasks (see Figure 9). There is no significant correlation between the visual orientation sequence task for the pretest and the tactile orientation sequence task in the first training sessions (r = −.05, p = .84, Pearson’s correlation analysis). However, significant correlation was observed between the visual orientation sequence task for the posttest and the tactile orientation sequence task in the last training sessions (r = .63, p = .009). Furthermore, there is no significant correlation detected between the visuospatial task and the tactile orientation sequence task (pretest and first training sessions: r = .29, p = .27; posttest and last training sessions: r = .123, p = .651).

Correlation between tactile orientation sequence task (training) and visual orientation sequence task (evaluation). The line of best fit was obtained from “Pearson” correlation analysis. (A) There was no significant correlation between accuracy on the pretest visual orientation sequence task and Day 1 of the tactile training task. (B) There was a significant correlation between accuracy on the posttest visual orientation sequence task and Day 9 of the tactile training task.
Discussion
In the current study, the behavioral training effects of tactile orientation sequence task training on two visual working memory tasks were investigated. The results showed that participants’ accuracy in the tactile orientation sequence task was improved with training sessions. In addition, participants’ accuracy in the visual orientation sequence task was significantly higher after tactile training. The item capacity of the visuospatial task at the posttest was also higher than that at the pretest in the training group. Moreover, a significant correlation was observed between the visual sequence task for the posttest and the tactile sequence task in the last training sessions.
Tactile orientation sequence task performance showed significant training-related improvement over the five training sessions. These results were consistent with our hypothesis, which indicated that tactile training could improve the performance of working memory for temporal sequence tasks in the tactile modality. It is assumed that the improvements in accuracy were caused by the practice of memorizing orientation stimuli with temporal sequences in the training task. Recent research has demonstrated that exposure to temporal sequences without feedback boosts participants’ ability to predict upcoming events, and these improvements are correlated with working memory (Baker et al., 2014, 2015). The present experimental paradigm was used to investigate the training effects in the tactile modality and found that participants’ accuracy in the tactile orientation sequence task was improved with training sessions. Studies in humans and nonhuman primates have suggested that working memory entails a gradient of abstraction from sensory areas reflecting low-level sensory features to prefrontal regions encoding more abstract, semantic, and response-related aspects of stimuli (Tulving & Thomson, 1973). It is speculated that information processing consisted of two steps in our training task: the first step was to detect the orientation of the stimuli, which was a similar process for the visual and tactile modalities, and the second step was to remember the temporal sequence rule. Thus, a potential explanation can be inferred for these training effects: participants were accustomed to holding the rule in temporary storage based on active learning, enabling them to better remember the regular sequence information. Recent research has shown that the human brain is highly plastic and adapts to new experiences (Jolles & Crone, 2012). It has been demonstrated that the essence of training exercises may produce either a specific impact on the efficiency of the targeted brain network or a more general influence affecting the dynamic processing of the human brain (Tang & Posner, 2009). These results indicated a significant positive training effect in the tactile working memory task.
Notably, participants’ accuracy in the visual orientation sequence task was significantly higher after tactile training, indicating a near-transfer effect from tactile training to the visual orientation sequence task, which might depend on sequential processing similar to that used in the training task. Previous studies have supported the hypothesis that visual and tactile recognition regions may provide a supramodal representation of objects to integrate information delivered by different sensory sources at higher cognitive levels (Ricciardi et al., 2006). The improvement between the visual pre- and posttests indicated the transfer of training effects from tactile to visual memory, which was supported by the supramodal hypotheses for visual and tactile modalities (Bonino et al., 2008; Cattaneo & Vecchi, 2008). The training and evaluation task used in the current study contains two different types of training styles: one that influences perceptual ability and one that affects cognitive ability. Perceptually, the current study used the stimuli of the visual grating and JVP domes consisting of tactile grating for orientation detection (Johnson et al., 1997). A previous study indicated that tactile working memory depends on a distributed network of brain regions analogous to the representation of visuospatial information (Schmidt & Blankenburg, 2018). It can be speculated that our transfer effects depend on the similarity of the distributed network for both visual and tactile working memory. In addition, the common perception task for orientation detection was associated with training and evaluated tasks, which contributed to these transfer effects. Another type of training is based on cognition for working memory; the tactile orientation sequence task used in the current study required the participants to temporarily store a stimulus and compare it with a specified target. These cognitive abilities are not specific to any single modality and present general improvements in temporal working memory. In conclusion, these results provide evidence for the transfer of training effects between visual and tactile modalities and indicate that general improvements in cross-modal working memory benefit from tactile training in a single modality.
In addition, we noticed that Schneiders et al. (2012) trained working memory with an auditory N-back task and evaluated the training effects with both auditory and visual N-back tasks. The transfer effect in the visual modality was not as strong as the training effect in the auditory modality. A probable reason for this outcome is that the perceptual tasks between visual and auditory modalities are quite different. In the study by Schneiders et al. (2012), rhythmic three-tone auditory melodies consisting of pure tones were used in the auditory training task, and abstract visual black-and-white pattern stimuli were used in the visual transfer task. Both auditory and visual stimuli cannot be abstracted from a sensory nature, so the results employed dissociated modality-specific working memory (Schneiders et al., 2011). In addition, many studies have suggested that visual and tactile modalities share abstract representations of spatial features such as shape, orientation, and structure (Loomis, 1982; Newell et al., 2005). However, the inputs of auditory stimuli consist of an acoustic phonological code that is entirely different from that encountered in a visual modality (Easton et al., 1997; Greene et al., 2001); this difference is a possible reason for the lack of transfer effects from auditory training to the visual modality. Therefore, it can be speculated that the transfer effects from tactile training tasks to visual evaluation tasks were caused by the shared representations in visual and tactile modalities.
Moreover, the item capacity of the visuospatial task at the posttest was also higher than that at the pretest in the training group. Consistent with our hypothesis, these results revealed a far-transfer effect from the tactile sequential task to the visuospatial task, which indicated the generality of training effects for working memory. In the visuospatial task, participants needed to compare the reference stimuli and target stimuli based on spatial information and report whether the two stimuli were identical. The spatial stimuli were processed by the visuospatial sketchpad, which is associated with the processing of visual information (A. Baddeley, 2012). Recent research has demonstrated that tactile delayed-match tasks activate the dorsolateral prefrontal cortex and the somatosensory cortex when participants encode tactile stimuli. The prefrontal cortex and parietal cortex have been associated with the maintenance phase of working memory (Di Zhao et al., 2017; Gazzaley & Nobre, 2012). We speculate that the training task not only had a practice effect on the visual and tactile sequence tasks through working memory but also increased the participants’ ability to memorize specific information and recall it. These results showed a significant upward trend between the pre- and posttests in the training group and demonstrated that sequence training could improve working memory ability as evaluated by different tasks.
Furthermore, a significant correlation between tactile and visual sequence tasks was detected, which indicated that the benefits from training were obvious in sequential task performance across different modalities. It can be speculated that participants did not understand the connection between the training task and the evaluation task at the beginning of training. Then, the correlation between pretest and Training Session 1 was not significant. With additional training sessions, the sequential processing aspect of the task started to become fully developed. Participants began to use the same strategy to execute visual and tactile sequence tasks, but this did not occur until later sessions; therefore, a significant correlation was reflected only between the posttest and Session 5 of the training task. There was no significant correlation between the tactile sequence training task and the visuospatial task. A possible reason for this outcome is that these two visual evaluation tasks rely on different processes (Manohar et al., 2017); the visual orientation sequence task is a typical serial processing task that is similar to tactile training (Hurlstone et al., 2014), whereas the visuospatial task uses parallel processing (Manohar et al., 2017). In addition, the visual gratings used in the visual orientation sequence task were similar to tactile JVP domes used in tactile training, and both stimuli were used to perform the orientation detection task (Tremblay et al., 2000; Zangaladze et al., 1999). The color points used in the visuospatial task simultaneously presented the combination of color and location; participants needed this information to perform the color change detection task (Luck & Vogel, 1997). The improvement observed between pre- and posttest indicated that visuospatial task performance benefited from training due to the general processes of working memory. However, the difference in perceptual processes between orientation detection and color change detection is a possible reason for the lack of correlation between the temporal training task and visuospatial task.
A previous study used a series of visuospatial working memory tasks with passive storage and active processing conditions to evaluate working memory performance in younger and older participants. The results indicated that active tasks were more sensitive in detecting differences within the older group (Vecchi & Cornoldi, 1999). One of the purposes of training is to delay the decline in cognitive ability for older participants. It is become clear that perception, behavior, and cognition in human subjects can be systematically improved by training tasks (Ball et al., 2002; Jaeggi et al., 2008; Vecchi & Cornoldi, 1999). Previous studies have adopted many different experimental paradigms to train tactile working memory (Gui et al., 2018; Leo et al., 2018). Based on the advantages of sequential processing for the tactile modality, most tasks, such as the N-back and delayed-match tasks, adopt a sequential mode of presentation (Dehghan Nayyeri et al., 2019; Gui et al., 2017). These different tasks have revealed positive training effects. We expected that future studies with simultaneous processing tasks could provide additional possibilities for tactile working memory training. Furthermore, we speculate that we might obtain better training effects via more sophisticated phased training and evaluation tasks.
Conclusion
The current study showed that practicing a tactile sequence task had a significant positive effect on the performance of a visual orientation sequence task. We demonstrated that a positive training effect could be transferred from a tactile task to a visual task. This training effect could also be transferred to different behavioral tasks that shared the same cognitive processes. Based on these results in young adults, our future studies may focus on developing effective training methods for older adults and exploring more effective methods to delay cognitive decline in the human brain through training.
Footnotes
Author Contributions
All the authors contributed to the conception of this work and to the drafting/revising of the manuscript. All the authors have approved the final version and agree to be accountable for all aspects of the work. T.G., Y.R., and Y.Y. designed the experiments with the help of Y.E. and J.W. The experiments were carried out by T.G., Y.Y., and Y.H., and the data were analyzed by T.G. with the help of Y.R. and Y.Y. T.G., Y.Y., and Y.R. drafted the full manuscript. Q.W., J.Y., S.T., and Y.E. commented on the manuscript and aided in the interpretation of the data.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: This study was supported by the Japan Society for the Promotion of Science (JSPS) KAKENHI Grant Nos. 17K18855, 17K00288, 18K18835, 18H01411, 19KK0099, 20K07722, and 20K04381 and a Grant-in-Sid for Strategic Research Promotion from Okayama University, the National Natural Science Foundation of China (31800932), Science and Technology Planning Project of Guizhou Province (QianKeHeJiChu-ZK [2021] General 120), and the Innovation and Entrepreneurship Project for High-level Overseas Talent of Guizhou Province ((2019)04).
Ethical Statement
All participants provided written informed consent to the procedure, which was previously approved by the ethics committee of Okayama University.