
Abstract
In this study, we examined the inhibitory effects of bergamottin on melanogenesis in B16F10 murine melanoma cells, together with its effects on the mechanism of melanin synthesis. α-Melanocyte stimulating hormone-stimulated B16F10 cells were treated with various concentrations of bergamottin, with arbutin as a positive control. Bergamottin significantly decreased the melanin content and tyrosinase activity without showing any cytotoxicity. In addition, bergamottin treatment significantly downregulated the expression of tyrosinase-related protein-1,2 and tyrosinase by suppressing the expression of microphthalmia-associated transcription factor. The phosphorylation status of mitogen-activated protein kinases (MAPKs) and protein kinase B (AKT) was examined to determine the mechanism underlying the antimelanogenic effects of bergamottin. Bergamottin treatment increased the phosphorylation of extracellular signal-regulated kinase (ERK) and AKT, but decreased the phosphorylation of p38 and c-Jun N-terminal kinase in the B16F10 cells. Moreover, the use of PD98059 (ERK inhibitor) and LY294002 (AKT inhibitor) corroborated these findings, indicating that bergamottin inhibits melanogenesis via the MAPKase and AKT signaling pathway. Thus, bergamottin has potential for treating hyperpigmentation disorders and can be a promising chemical for skin-whitening in the cosmetic industry.
Melanogenesis describes the production process of melanin pigment in melanocytes. 1,2 Melanocytes are dendritic cells of the neuroectoderm located at the basal layer in the epidermal-dermal junction, which deliver melanosomes to surrounding keratinocytes. 3,4 In mammals, the biosynthesis of melanin is induced by several stimuli, including UV radiation, skin irritation, metabolic disorders, and other underlying problems. 5 Melanin is important for protecting against photo-damage caused by harmful UV radiation and coloration of skin and hair in mammals. 6 However, overproduction and accumulation of melanin may cause skin diseases, such as lentigo, aging, freckles, and other hyperpigmentation disorders. 7,8
Tyrosinase largely regulates the process of melanin synthesis in early stages, as it catalyzes the o-hydroxylation of L-tyrosine (monophenol) to L-dihydroxyphenylalanine (L-DOPA) and the subsequent oxidation of L-DOPA to dopaquinone. 9 In addition, tyrosinase-related protein-1 (TRP-1) oxidizes 5,6-dihydroxyindole-2-carboxylic acid (DHICA) to indole-5,6-quinone-2-carboxylic acid in mice but has not been reported to have the same activity in humans. Tyrosinase-related protein-2 (TRP-2), also known as dopachrome tautomerase, is able to isomerize dopachrome to form DHICA. 10
Recent studies have indicated that microphthalmia-associated transcription factor (MITF) is an important transcriptional regulator which regulates the expression of essential melanogenic enzymes, such as TRP-1, -2, and tyrosinase. UV-induced α-melanocyte stimulating hormone (α-MSH) stimulates the expression of MITF by activating signaling pathway-related enzymes, including protein kinase A (PKA) and cyclic adenosine monophosphate (cAMP) responder element binding protein (CREB). In addition, the increased MITF binds to the M-box of tyrosinase promoters, further regulating the expression of melanogenic enzymes. Accordingly, it has recently been reported that hyperpigmentation can be prevented by the inhibition of MITF. 11,12
Mitogen-activated protein kinases (MAPKs), which are serine/threonine kinases that include p38, extracellular signal-regulated kinase (ERK), and c-Jun N-terminal kinase (JNK), are well-known important factors for melanogenesis. Recent studies have also demonstrated that these MAPKs are involved in regulating MITF expression. In addition, several studies have indicated that the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (also known as AKT) signaling pathway is implicated in mammalian melanogenesis. Phosphorylated ERK and AKT promote MITF ubiquitination and degradation, as well as decreased tyrosinase expression, which suppresses melanogenesis. In contrast, phosphorylated JNK and p38 can induce melanogenesis by inhibiting MITF degradation. 13 -16
Recently, numerous studies regarding the treatment of hyperpigmentation disorders have been conducted and have identified key tyrosinase inhibitors, including hydroquinone, kojic acid, ascorbic acid, and arbutin. However, these whitening agents are limited by their cytotoxicity, low stability to oxygen and water, cutaneous irritation, and dermatitis. Thus, developing skin-whitening agents derived from natural sources that show reduced side effects is urgently needed. 17 -20
Bergamottin (4-[(2E)-3,7-dimethylocta-2,6-dienoxy]furo[3,2-g]chromen-7-one), a natural furanocoumarin, is found in the pulp of Citrus maxima and Citrus х paradisi. 21 It has been reported that bergamottin occurs naturally in the peel and pulp of Citrus bergamia and in the roots and rhizomes of Notopterygium incisum, which are traditional Chinese medicinal herbs. 22 Bergamottin exerts strong pharmaceutical bioactivities due to its anticancer (antiproliferation, anti-invasion, and antimigration), antimycobacterial, and antimutagenicity activities. 23 -27 In addition, bergamottin has been reported to increase drug bioavailability via interactions with some isoforms of cytochrome P450. 28 -30 However, the antimelanogenic effects of bergamottin have not been studied to date. Therefore, this study aimed to determine the inhibitory effect of bergamottin on melanogenesis in B16F10 murine melanoma cells and determine the underlying molecular mechanisms of this process.
To determine the cytotoxicity of bergamottin on B16F10 melanoma cells, cell viability was evaluated using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The cells were treated with bergamottin (either 1.56, 3.13, 6.25, 12.5, or 25 µM) in the absence or presence of α-MSH for 72 hours. As shown in Figure 1(b), no significant differences were observed with concentrations ranging from 1.56 to 12.5 µM compared to the respective untreated control cells. Therefore, these concentrations of bergamottin (3.13, 6.25, and 12.5 µM) were used for further experimentation.
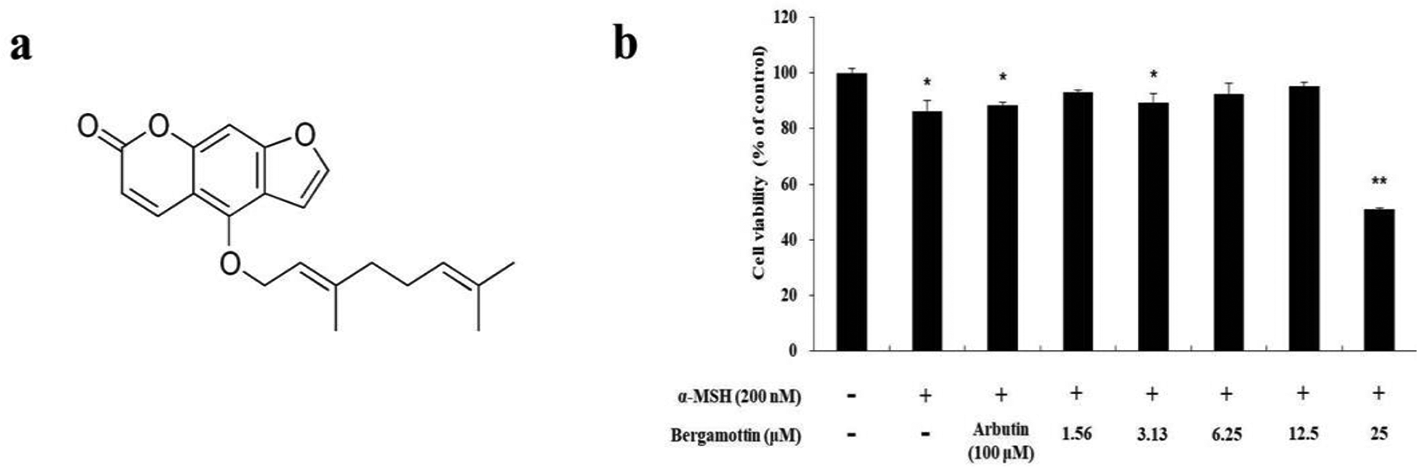
(a) The chemical structure of bergamottin. (b) The effects of bergamottin on the viability of B16F10 melanoma cells. The cells were treated with bergamottin (1.56, 3.13, 6.25, 12.5, and 25 µM) for 72 hours. Cell viability is expressed as percentages compared to the respective values obtained for untreated control cells. The data are presented as mean ± SD of at least 3 independent experiments. *P < 0.05; **P < 0.01.
Tyrosinase is a rate-limiting enzyme in melanin synthesis and recent studies have indicated that its overexpression leads to hyperpigmentation disorders by inducing melanogenesis. 31 Thus, determination of melanin content indicates that melanogenesis is related to tyrosinase activity, which is involved in melanin synthesis. Because melanogenesis is regulated by tyrosinase, the melanin content and tyrosinase activity were examined in B16F10 cells. The cells were treated with bergamottin at various concentrations (3.13, 6.25, and 12.5 µM), whereas α-MSH (200 nM) and arbutin (100 µM) were used as negative and positive controls, respectively. As shown in Figure 2, bergamottin significantly decreased the melanin content and tyrosinase activity in a concentration-dependent manner in α-MSH-induced B16F10 cells. The melanin content and tyrosinase activity of 3.13, 6.25, and 12.5 µM bergamottin-treated cells decreased by 8.2%, 13.1%, and 37.6%, and 1.1%, 10.4%, and 44.6%, respectively, compared to the α-MSH-stimulated cells. These results indicated the strong relation between reduced melanin content and inhibition of tyrosinase activity.

Effects of bergamottin on (a) melanin content and (b) tyrosinase activity of B16F10 melanoma cells. The cells were treated with bergamottin (3.13, 6.25, and 12.5 µM) for 72 and 24 hours, respectively. α-MSH (200 nM) and arbutin (100 µM) were used as negative and positive controls, respectively. The melanin content and tyrosinase activity in the treated cells were expressed as percentages compared to the respective values obtained for the control groups. The data are presented as mean ± SD of at least 3 independent experiments. ## P < 0.01 in comparison with the untreated cell group and *P < 0.05, **P < 0.01 in comparison with the α-MSH-treated group.
To further understand the mechanism of action of bergamottin, Western blot assays were performed to examine the expression of key enzymes associated with the signaling pathways of melanin synthesis in B16F10 cells. MITF is known to be an important transcription factor which regulates the expression of melanogenic enzymes including TRP-1, TRP-2, and tyrosinase by binding to the M-box in the respective promoter regions, enhancing melanogenesis. 11,12 As expected, bergamottin reduced the expression of MITF, TRP-1, TRP-2, and tyrosinase at the treatment concentrations, further reducing expression at the highest treatment concentration of 12.5 µM (Figure 3). These results suggested that bergamottin inhibited the expression of melanogenic enzymes by downregulating MITF expression.

The effects of bergamottin on TRP-1, TRP-2, tyrosinase, and MITF expression in B16F10 cells. Cells were treated with the indicated concentrations of bergamottin for 24 hours. Protein levels were determined by Western blotting. (a) Western blotting results and protein levels of (b) TRP-1, (c) TRP-2, (d) tyrosinase, and (e) MITF. Results are expressed as percentages of the control groups. The data are presented as mean ± SD of at least 3 independent experiments. ## P < 0.01 in comparison with the untreated cell group and **P < 0.01 in comparison with the α-MSH-treated group.
The phosphorylation of p38 and JNK in MAPKs induces melanogenesis via increasing MITF expression. In contrast, ERK phosphorylation is involved in downregulating melanogenesis. 13,14,16 Therefore, we evaluated the effects of bergamottin on MAPK phosphorylation in B16F10 cells. As shown in Figure 4, bergamottin decreased the phosphorylation of p38 and JNK, while increasing ERK phosphorylation, especially at the highest concentration tested (12.5 µM). To determine whether the increased ERK phosphorylation is involved in reducing melanogenesis via bergamottin treatment, the cells were treated with PD98059, an ERK inhibitor. As shown in Figure 5, the Western blot and tyrosinase activity test results indicated that tyrosinase activity reduced in α-MSH-stimulated B16F10 melanoma cells treated with bergamottin, but it increased upon the inhibitor treatment. These results suggest that bergamottin inhibits melanogenesis by decreasing MITF expression through the MAPK signaling pathway.

Effects of bergamottin on the phosphorylation of ERK, JNK, and p38. B16F10 cells were treated with bergamottin at the indicated concentrations for 4 hours. (a) Western blotting results and corresponding protein levels of (b) p-ERK, (c) p-JNK, and (d) p-p38. Data are expressed as percentages of the control groups. The data are presented as mean ± SD of at least 3 independent experiments. ## P < 0.01 in comparison with the untreated cell group and **P < 0.01 in comparison with the α-MSH-treated group. P, phosphorylated; T, total.

Effects of the ERK inhibitor PD98059 on (a) Western blot and (b) tyrosinase activity in B16F10 cells. Cells were treated with α-MSH (200 nM) or PD98059 (2 µM) alone or in combination, as well as with or without 12.5 µM bergamottin. Protein expression levels were investigated by Western blotting. Data are expressed as percentages of the control groups. The data are presented as mean ± SD of at least 3 independent experiments. ## P < 0.01 in comparison with the untreated cell group and *P < 0.05, **P < 0.01 in comparison with the α-MSH-treated group.
cAMP-induced PI3K/AKT signaling pathway inhibition results in decreased AKT phosphorylation and diminishes the phosphorylation of glycogen synthase kinase 3 beta (GSK-3β), suppressing melanogenesis. 32,33 To determine the effect of bergamottin on AKT phosphorylation in B16F10 cells, Western blot analysis was performed after treating cells with varying concentrations of bergamottin. As shown in Figure 6a,b, bergamottin dramatically increased AKT phosphorylation in the treated cells compared with the untreated controls. Next, we examined the effect of LY294002 (an AKT inhibitor) on tyrosinase activity to investigate whether AKT activation was involved in melanogenesis. As shown in Figure 6(c), tyrosinase activity in α-MSH-stimulated B16F10 melanoma cells treated with bergamottin was increased after the LY294002 treatment, implying the direct involvement of PI3K/AKT signaling in bergamottin-mediated melanogenesis.
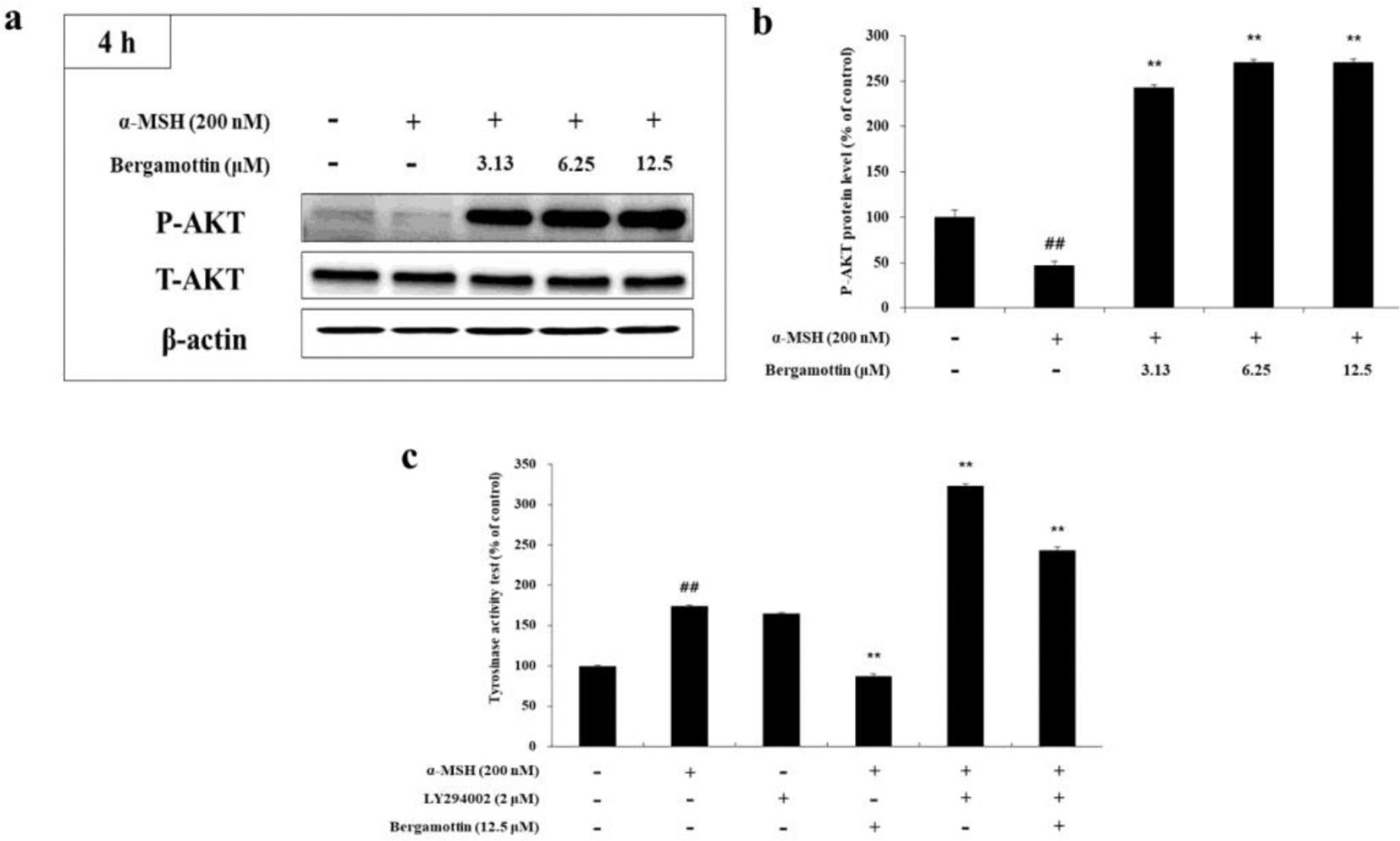
The effects of bergamottin on AKT phosphorylation. B16F10 cells were treated with the indicated concentrations of bergamottin for 4 hours. (a) Western blotting and (b) protein levels of p-AKT. (c) Effect of the AKT inhibitor LY294002 on tyrosinase activity in the treated B16F10 cells. Cells were treated with α-MSH (200 nM) or LY294002 (2 µM) alone or in combination as well as with or without 12.5 µM of bergamottin. Data are expressed as percentages of the control groups. The data are presented as mean ± SD of at least 3 independent experiments. ## P < 0.01 in comparison with the untreated cell group and *P < 0.05, **P < 0.01 in comparison with the α-MSH-treated group. P, phosphorylated; T, total.
In conclusion, herein we have demonstrated the inhibitory effect of bergamottin on melanin synthesis and cellular tyrosinase activity in α-MSH-stimulated B16F10 cells. We also showed that bergamottin can suppress the expression of MITF, as well as melanogenic enzymes such as TRP-1, TRP-2, and tyrosinase. Moreover, bergamottin treatment decreased p38 and JNK phosphorylation and increased ERK and AKT phosphorylation. The use of PD98059 (an ERK inhibitor) and LY294002 (an AKT inhibitor) corroborated these findings. The results presented herein indicate that bergamottin inhibits melanogenesis via MAPK and AKT signaling pathways in α-MSH-stimulated B16F10 cells. Therefore, bergamottin has a potential to be used for treating hyperpigmentation disorders and may be a promising candidate as an active ingredient in skin-whitening creams in the cosmetic industry.
Experimental
Chemicals and Reagents
Bergamottin, dimethyl sulfoxide (DMSO), α-MSH, NaOH, MTT, radioimmunoprecipitation assay (RIPA) buffer, and L-DOPA were obtained from Sigma-Aldrich (St Louis, MO, USA). Dulbecco’s Modified Eagle’s Medium (DMEM), fetal bovine serum (FBS), penicillin/streptomycin, and trypsin-ethylenediaminetetraacetic acid were purchased from Thermo Fisher Scientific (Waltham, MA, USA). LY294002 (AKT inhibitor) and antibodies for tyrosinase, TRP-1, TRP-2, and MITF were purchased from Santa Cruz Biotechnology (Dallas, TX, USA). Antibodies for p-p38, p38, p-JNK, JNK, p-ERK, ERK, p-AKT, AKT, and β-actin were obtained from Cell Signaling Technology (Danvers, MA, USA). PD98059 (p38 inhibitor) was purchased from Cayman Chemical (Ann Arbor, MI, USA). The enhanced chemiluminescence (ECL) kit and 2 × Laemmli sample buffer were obtained from Biosesang (Seongnam, Gyeonggi-do, Korea) and Bio-Rad (Hercules, CA, USA), respectively.
Cell Culture
Murine melanoma cells (B16F10) were purchased from the Korean Cell Line Bank (Seoul, Korea) and cultured in DMEM supplemented with 1% penicillin/streptomycin and 10% FBS. The cells were incubated in a humidified atmosphere containing 5% CO2 at 37°C and were subcultured every 4 days.
Measurement of Cell Viability
Cell viability was measured using the MTT assay. The B16F10 cells were cultured in 24-well plates for 24 hours and subsequently treated with various concentrations of bergamottin (1.56, 3.13, 6.25, 12.5, and 25 µM) for 72 hours. The MTT solution was added to the plate and subsequently removed after incubation for 4 hours. The formed formazan crystals were dissolved in 1 mL of DMSO and the absorbance was measured in a 96-well plate at 540 nm using an ELISA reader (Tecan, Mannedorf, Switzerland).
Measurement of Intracellular Melanin Content
The B16F10 cells were treated in a 6-well plate with various bergamottin concentrations (3.13, 6.25, and 12.5 µM) for 72 hours. In addition, α-MSH (200 nM) and arbutin (100 µM) were used as negative and positive controls, respectively. After removal of DMEM, the cells were washed twice with cold phosphate-buffered saline (PBS) and dissolved in 1 N NaOH for 1 hour at 80°C. Subsequently, 200 µL aliquots of the media were placed into 96-well plates and the absorbance was measured at 405 nm using an ELISA reader. All experiments were performed in triplicate.
Measurement of Intracellular Tyrosinase Activity
The B16F10 melanoma cells (8.0 × 104 cells/dish) were seeded into 60 mm dishes and treated with various bergamottin concentrations (3.13, 6.25, and 12.5 µM) and α-MSH (200 nM) for 24 hours. α-MSH (200 nM) and arbutin (100 µM) were used as negative and positive controls, respectively. After DMEM was removed, the cell pellets were collected and lysed using RIPA buffer containing a 1% protease inhibitor cocktail. The lysates were clarified by centrifugation at 15 000 rpm for 30 minutes. The amount of protein in each lysate was quantified using the bicinchoninic acid (BCA) kit. Subsequently, 20 µL of each lysate was mixed with 80 µL of L-DOPA (2 mg/mL) in a 96-well plate. After incubation at 37°C for 2 hours, the absorbance of each well was measured at 490 nm.
Western Blot Assay
The B16F10 cells were seeded into 60 mm dishes and subsequently incubated with α-MSH (200 nM) and various bergamottin concentrations (3.13, 6.25, and 12.5 µM). After incubation, the cells were washed thrice with cold PBS and treated with trypsin/EDTA to collect cell pellets in 1.5 mL e-tubes. The cells were subsequently lysed for 1 hour in RIPA buffer containing a 1% protease inhibitor cocktail. After centrifugation at 15 000 rpm for 30 minutes at -8°C, the supernatant was collected in a new 1.5 mL e-tube. The amount of protein in each cell lysate was quantified using the BCA kit. After adjusting the protein levels, Western blot samples were denatured at 100°C for 5 minutes, and subsequently 20 µL of each sample was loaded onto 10% sodium dodecyl sulfate polyacrylamide gels for 1 hour at 150 V. The gels were then transferred to polyvinylidene difluoride membranes for 7 minutes. The membranes were blocked using TBST containing a 5% skim milk solution for 1 hour and subsequently washed 6 times every 10 minutes with Tris-buffered saline (20 mM Tris base, 137 mM NaCl, pH 7.6) containing 0.4% Tween 20 (TBST) for 1 hour. The membranes were then incubated for 24 hours at 4°C with specific primary antibodies (1:1000) for 24 hours. The membranes were subsequently washed with TBST every 10 minutes for 1 hour to remove any unbound primary antibodies. Each membrane was then incubated with a secondary antibody (IgG, 1:3000) for 2 hours and washed with TBST every 10 minutes for 1 hour. The protein bands were detected using an ECL kit. The amount of protein expression was quantified and graphed using the imageJ program (NIH, Bethesda, MD, USA).
Statistical Analysis
The results of the experiments were analyzed using Student’s t-test, and p-values of <0.05 (*) or 0.01 (**) were considered statistically significant. The data are presented as mean ± SD of at least 3 independent experiments.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was financially supported by the Ministry of Trade, Industry, and Energy (MOTIE), Korea, under the “Regional Specialized Industry Development Program” (reference number P0006063) supervised by the Korea Institute for Advancement of Technology (KIAT).