L -Cysteine protects intestinal integrity,attenuates intestinal inflammation and oxidant stress,and modulates NF-κB and Nrf2 pathways in weaned piglets after LPS challenge
Free accessResearch articleFirst published online April, 2016
L -Cysteine protects intestinal integrity,attenuates intestinal inflammation and oxidant stress,and modulates NF-κB and Nrf2 pathways in weaned piglets after LPS challenge
In this study we investigated whether L-cysteine (L-cys) could alleviate LPS-induced intestinal disruption and its underlying mechanism. Piglets fed with an L-cys-supplemented diet had higher average daily gain. L-cys alleviated LPS-induced structural and functional disruption of intestine in weanling piglets, as demonstrated by higher villus height, villus height (VH) to crypt depth (CD) ratio, and transepithelial electrical resistance (TER) and lower FITC-dextran 4 (FD4) kDa flux in jejunum and ileum. Supplementation with L-cys up-regulated occludin and claudin-1 expression, reduced caspase-3 activity and enhanced proliferating cell nuclear antigen expression of jejunum and ileum relative to LPS group. Additionally, L-cys suppressed the LPS-induced intestinal inflammation and oxidative stress, as demonstrated by down-regulated TNF-α, IL-6 and IL-8 mRNA levels, increased catalase, superoxide dismutase, glutathione peroxidase activity, glutathione (GSH) contents and the ratio of GSH and oxidized glutathione in jejunum and ileum. Finally, a diet supplemented with L-cys inhibited NF-κB(p65) nuclear translocation and elevated NF erythroid 2-related factor 2 (Nrf2) translocation compared with the LPS group. Collectively, our results indicated the protective function of L-cys on intestinal mucosa barrier may closely associated with its anti-inflammation, antioxidant and regulating effect on the NF-κB and Nrf2 signaling pathways.
The intestinal barrier is important as a selective barrier against endogenous and exogenous noxious antigens and pathogens.1 Disruption of the intestinal barrier promotes luminal antigens to penetrate subepithelial tissues, inducing a mucosal and systemic inflammatory response, which is the major pathogenesis for most of intestinal diseases.2 Multiple factors, including inflammation and oxidative stress, can give rise to intestinal barrier damage.2–4 Over-production of pro-inflammatory cytokines and reactive oxygen species (ROS) can disrupt the intestinal epithelial integrity and function.3–5 Therefore, inhibition of intestinal inflammation and oxidative stress may exert beneficially prevent the intestinal disruption.
Traditionally, L-cysteine (L-cys) is thought to be a non-essential amino acid and mainly participates in protein metabolism. However, many studies have reported its diverse non-nutritional properties.6–10L-cys has been found to suppress effectively inflammation and oxidative stress in diabetic rats and high Glc-treated cells.6,8,11 Moreover, L-cys could alleviate oxidative stress via GSH synthesis in inflammatory bowel disease.3 It has been established that L-cys supplementation ameliorates local inflammation, and improved intestinal barrier restoration in a dextran sulfate sodium (DSS)-induced porcine colitis model.9 These data demonstrated that L-cys was effective in suppressing inflammation and oxidative stress, and suggests L-cys might be a promising nutritional agent for protecting intestinal integrity. However, to our knowledge, little research has been conducted on the protective effect of L-cys on intestinal integrity.
In the current study, the protective effect of dietary L-cys supplementation was evaluated in a LPS-challenged porcine model of intestinal injury. Substantial evidences have demonstrated that LPS stimulates intestinal immune cells to rapid production of pro-inflammatory cytokines and leads to structural and functional injury in intestine.5 The porcine model, with its similar gastrointestinal morphology and physiology to humans, has been recognized as an appropriate experimental model for the study of inflammatory bowel disease (IBD). The LPS-challenged porcine model has been well-established in inducing acute intestinal injury and used in many IBD therapy studies. In the current experiments, we employed this model to elaborate whether L-cys could alleviate LPS-induced intestinal injury through its anti-inflammation, antioxidant and regulating effect on the NF-κB (p65) and NF erythroid 2-related factor 2 (Nrf2) signaling pathway.
Materials and methods
Animals and experiment design
All procedures were approved by the Animal Care and Use Committee, Zhejiang University. Twenty-four crossbred healthy barrows [(Duroc × Landrace × Yorkshire), 6.71 ± 0.56 kg] were weaned at 21 d of age and randomly assigned to four treatment groups (n = 6/group): (1) control group (Con; basal diet and i.p. injection of 0.9% sterile saline); (2) LPS group (LPS; basal diet and i.p. injection of LPS at 100 mg/kg body mass); (3) LPS + 0.25% L-cys group (LPS + 0.25% L-cys; basal diet supplemented with 0.25% L-cys and i.p. injection of LPS); (4) LPS + 0.5% L-cys group (LPS + 0.5% L-cys; basal diet supplemented 0.25% L-cys and i.p. injection of LPS). The basal diet was formulated to meet the requirements of the pigs (Table 1), according to National Research council swine requirements.12L-Cys doses (L-cysteine, purity > 99%; Hao Lai Li Technology Ltd. Co., Wuhan, China) were chosen according to our preliminary study. To obtain isonitrogenous diets, 0.37%, 0.18% and 0% alanine (purity > 99%; Hao Lai Li Technology Ltd. Co.) were added to the basal, 0.25% L-cys and 0.5% L-cys diets, respectively.
Ingredient composition of the basal diet (on an as-fed basis).
Provided per kg of diet: vitamin A, 6000 IU; vitamin D3, 500 IU; vitamin E, 40 IU; pantothenic acid, 20 mg; niacin, 30 mg; riboflavin, 5.0 mg; vitamin K3, 1.5 mg; thiamine, 2.0 mg; pyridoxine, 3.0 mg; biotin, 0.10 mg; folic acid, 0.6 mg; vitamin B12, 0.04 mg; choline, 800 mg; Cu (CuSO4·5H2O), 16 mg; Zn (ZnSO4), 100 mg; Fe (FeSO4), 125 mg; Mn (MnSO4·H2O), 15 mg; I (KI), 0.2 mg; Se (Na2SeO3), 0.3 mg.
Digestible energy was calculated from data provided by Feed Database in China.
After a 19-d feeding period, the challenged groups were received i.p. injections of Escherichia coli LPS (Sigma Chemical, St. Louis, MO, USA) at 100 µg/kg body mass (BM); the Con group was injected with the same volume of saline. The LPS dose was used according to our preliminary experiments and other references.5,13 BM and feed consumption between d 1 and 19 before LPS or saline injection were recorded daily. To avoid the potential effects of feed intake reduction induced by LPS on intestinal variables, all pigs were pair-fed until slaughter 24 h after LPS or saline injection. All piglets were individually housed in 1.3 × 1.2 m2 pens in an environmentally controlled nursery barn. Each treatment had six replicate pens. Each pen contained a feeder and a nipple waterer, to allow piglets ad libitum access to feed and water.
Sample collections
Twenty-four h after LPS or saline solution injection, all piglets were humanely killed by euthanasia with an i.v. injection of sodium pentobarbital (40 mg/kg BM). Two 5-cm-long segments and one 10-cm-long segment were acquired from the middle jejunum and ileum. One 5-cm segment was rinsed with PBS and then placed in 10% neutral buffered formalin until processing. The other 5-cm segments were collected for Ussing chamber experiments. The 10-cm intestinal segments were used for collecting mucosal samples. After harvesting, samples were immediately frozen in liquid nitrogen and stored at −80℃ until analysis.
Intestinal morphology
Segments for morphological study were embedded in paraffin, sectioned (5 µm), and stained with hematoxylin and eosin. Villus height and crypt depth were measured using a light microscope with an image processing and analysis system (Version 1; Leica Imaging Systems, Cambridge, UK). Values are expressed as means from 10 intact, well-oriented crypt-villus units at each slice in triplicate.
Ussing chamber experiments
Segments for Ussing chamber experiment were stripped from seromuscular layer in oxygenated (95% O2/5% CO2) Ringer’s solution. Then, samples were mounted in an Easy Mount Ussing chamber system (model VCC MC6; Physiologic Instruments, San Diego, CA, USA) and connected to Acquire and Analyze software (Physiologic Instruments) for automatic data collection. After a 30-min equilibration period on Ussing chambers, TER (Ω·cm2) was recorded at 15-min intervals over a 2-h period and the average TER value for each given pig was calculated. FD4 (Sigma-Aldrich, St. Louis, MO, USA) was added to the mucosal side. Mucosal-to-serosal flux of FD4 (µg/cm2/h) was measured by sampling 100 μl solution from the serosal side at 30-min intervals for 120 min and same amount of fresh Ringer’s solution were supplemented to maintain the solution volume constant. The concentrations of FD4 in the serosal side were measured by fluorescence microplate reader (FLx800; Bio-Tek Instruments Inc., Winooski, VT, USA). FD4 flux over the 2-h period were calculated.
Caspase-3 activity assay
The caspase-3 activity in jejunum and ileum mucosa was determined using the caspase-3 activity kit following the manufacturer’s protocols (Beyotime Institute of Biotechnology, Jiangsu, China). Caspase activities were expressed as the percentage of the Con group.
Redox status
Intestinal mucosal supernatants were used for the measurement of catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), malondialdehyde (MDA), glutathione (GSH) and oxidized glutathione (GSSG), using an ELISA kit specific for porcine models, following the manufacturer’s protocol (Nanjing Jiancheng Bioengineering Institute, Jiangsu, China).
Real-time PCR
RNA was isolated by AxyPrep total RNA miniprep kit (Axygen Scientific Inc., Union City, CA, USA) from mucosal samples, according to the manufacturer’s guidelines. RNA concentration was measured using a Nano Drop spectrophotometer (ND-2000; NanoDrop Technologies, Wilmington, DE, USA). The cDNA synthesis was performed by PrimeScripte RT reagent kit (TaKaRa Biotechnology, Dalian, China), following the manufacturer’s guidelines. Quantitative real-time PCR was performed with a Step One Plus real-time PCR system (Applied Biosystems, Foster City, CA, USA) using a SYBR premix EX Taq (TaKaRa Biotechnology), following the standard protocols. The primers used are presented in Table 2. Primer sequences were as follows: F 5’-ATGGTGAAGGTCGGAGTGAAC-3’ and R 5’-CTCGCTCCTGGAAGATGGT-3’ for GAPDH; F 5’-CATCGCCGTCTCCTACCA-3’ and R 5’-CCCAGATTCAGCAAAGTCCA-3’ for TNF-α; F 5’-CCTGTCCACTGGGCACATAAC-3’ and R 5’-CAAGAAACACCTGGCTCTGAAAC-3’ for IL-6; F 5’- TGGCAGTTTTCCTGCTTTCT-3’ and R 5’-CAGTGGGGTCCACTCTCAAT-3’ for IL-8. The relative changes in each target gene expression were analyzed using the 2−△△Ct method.5 All samples were run in triplicate.
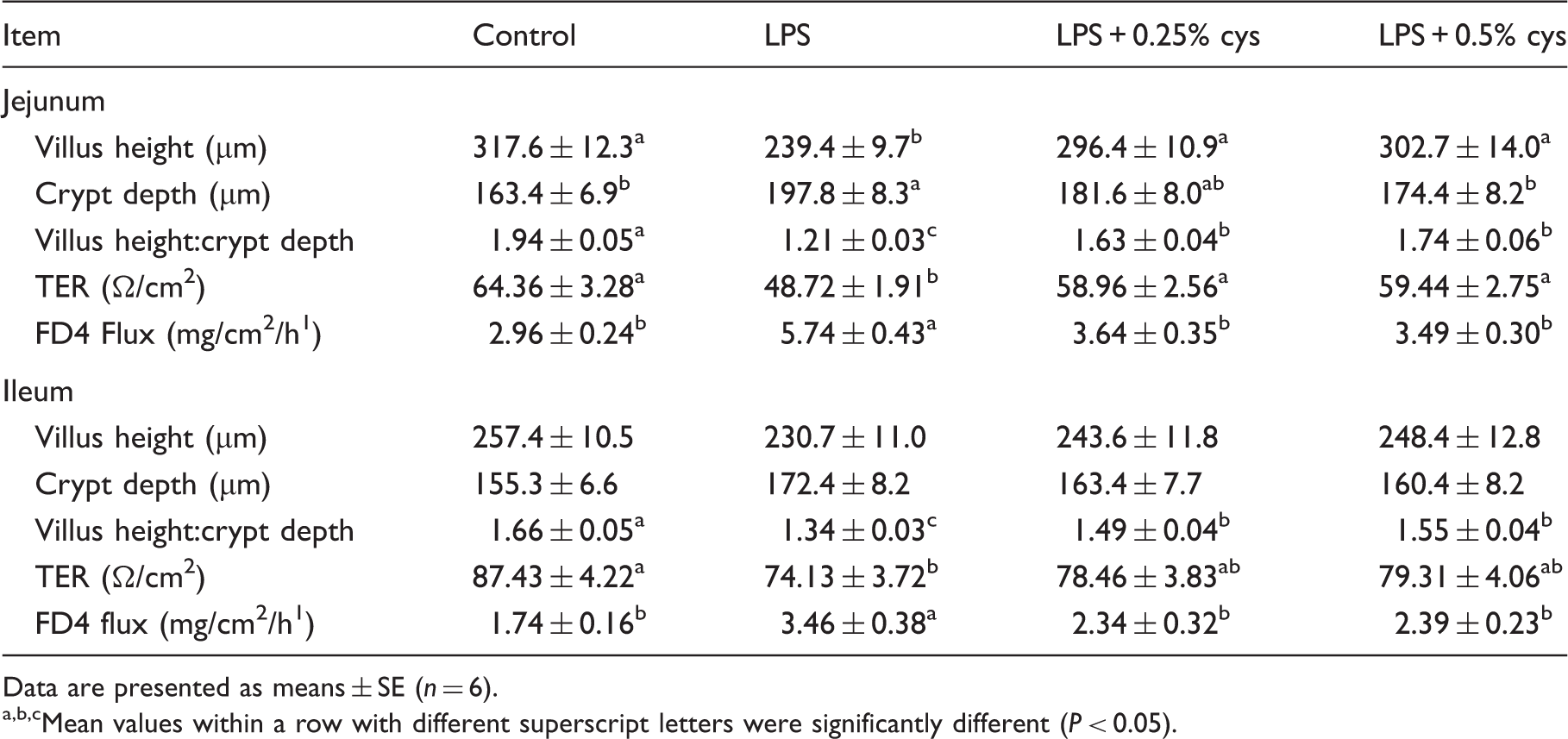
Effects of dietary L-cys supplementation on intestinal morphology and barrier function of weaned piglets.1
Mean values within a row with different superscript letters were significantly different (P < 0.05).
Western blot
The mucosal scrapings were thawed at 4℃ and then lysed on ice using chilled lysis buffer. This mixture was homogenized on ice and centrifuged to harvest the supernatants. The nuclear extract was extracted from the homogenized mucosa sample according to the manufacturer (Vazyme Biotech Co., Ltd., Nanjing, China). Protein concentration of extract was measured by an enhanced BCA protein assay kit (Beyotime Institute of Biotech). Next, tissue extract (equal amounts of protein) was boiled at 95℃ for 5 min with a quarter-volume of 5 × SDS-PAGE sample buffer. Lysates were separated by a SDS polyacrylamide gel. Afterwards, the proteins were transferred to polyvinylidene difluoride membrane (Millipore, Bedford, MA, USA). The membranes were incubated with primary Ab at 4℃ for 12 h, then with the secondary Ab for 60 min at room temperature (24℃). The occludin and β-actin Ab were purchased from Abcam Inc. (Cambridge, UK), and proliferating cell nuclear antigen (PCNA), zonula occluden-1 (ZO-1), claduin-1 NF-κB (p65), Nrf2 and nucleolin Ab were all purchased from Santa Cruz Technology (Santa Cruz, MA, USA). The HRP-conjugated secondary Ab was from Beyotime. The visualization of protein on the membrane were developed with an ECL detection kit (Beyotime Institute of Biotechnology), photographed by a ChemiScope 3400 (Clinx Science Instruments, Shanghai, China). The Western blotting result was evaluated using Quantity One software (Bio-Rad Laboratories, Berkley, CA, USA). β-Actin was used as an internal control. The relative changes of each target protein was expressed as fold changes to the Con group.
Statistical analysis
Data were analyzed using the SAS statistical package (SAS Institute, Cary, NC, USA),14 with each animal considered an experimental unit. Differences between means were tested using Duncan’s multiple range tests. Differences were considered significant at P < 0.05.
Results
Growth performance
During the 19-d feeding trial before challenge, differences were not observed in average daily gain (267 ± 9.53, 270 ± 10.46, 282 ± 10.15, 287 ± 11.45), daily feed intake (396 ± 15.92, 401 ± 14.36, 408 ±16.73, 412 ± 17.98) and feed to gain ratio (1.48 ± 0.04, 1.48 ± 0.04, 1.45 ± 0.04, 1.44 ± 0.05) among Con, LPS, LPS + 0.25% L-cys and LPS + 0.5% L-cys, respectively.
Intestinal morphology and barrier function
As shown in Table 2, LPS challenge reduced VH, VH/CD and TER, and increased CD and FD4 flux in jejunum, and reduced VH/CD and TER and increased FD4 flux in ileum compared with the Con group (P < 0.05). Piglets in the LPS + 0.25% L-cys group had higher VH, VH:CD and TER, and lower FD4 flux in jejunum and higher VH:CD and lower FD4 flux in ileum (P < 0.05) relative to the LPS group. Piglets in LPS + 0.5% L-cys had higher VH, VH:CD and TER, and lower FD4 flux in jejunum and higher VH:CD and lower FD4 flux in ileum (P < 0.05) compared with the LPS group.
Tight junction (TJ) expression
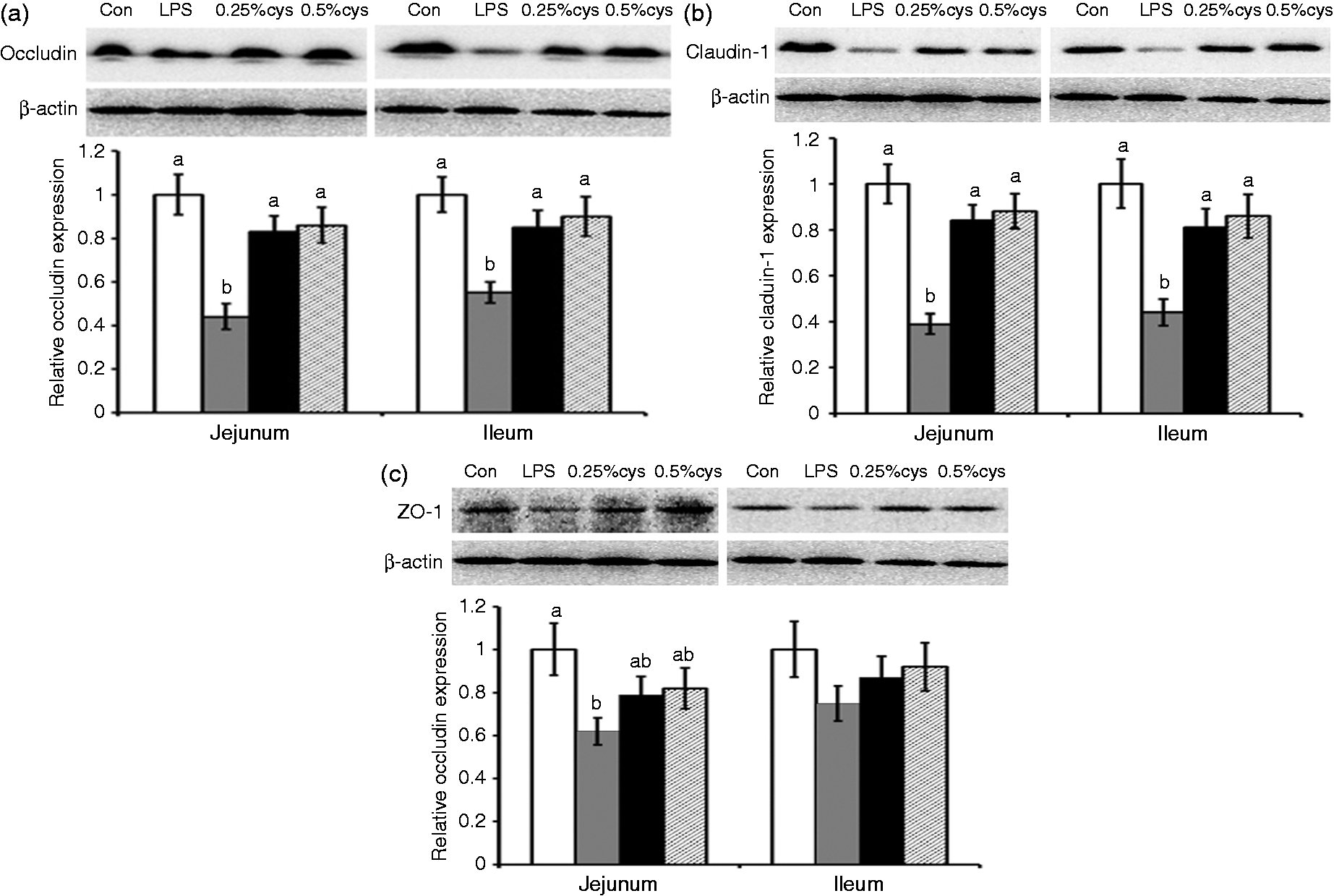
TJ expression in jejunum and ileum of piglets is presented in Figure 1. Relative to the Con group, LPS challenge reduced occludin, claudin-1 and ZO-1 expression in jejunum, and occludin and claduin-1 in ileum (P < 0.05). Supplementation with L-cys (0.25% or 0.5%) increased the protein expression of occludin and claudin-1, while it did not have significant effect on ZO-1 expression in jejunum and ileum compared with piglets in the LPS group (P < 0.05).
Effects of L-cys supplementation on the expression of (A) occludin, (B) claudin-1 and (C) ZO-1 in jejunum and ileum mucosa of weaned piglets. Values are means and SE represented by vertical bars. a,bMeans with different letters differ significantly (P < 0.05). Control group (Con; white bars) was fed with basal diet and injected with 0.9% sterile saline; LPS group (LPS; dark gray bars) was fed with basal diet and injected with LPS; LPS + 0.25% L-cys group (0.25% cys; black bars) was fed with a 0.25% L-cys-supplemented diet and injected with LPS; LPS + 0.5% L-cys group (0.5% cys; hatched bars) was fed with a 0.5% L-cys-supplemented diet and injected with LPS. The control sample was used as the reference sample. The protein expression of all samples was expressed as fold changes, calculated relative to Con group.
Apoptosis
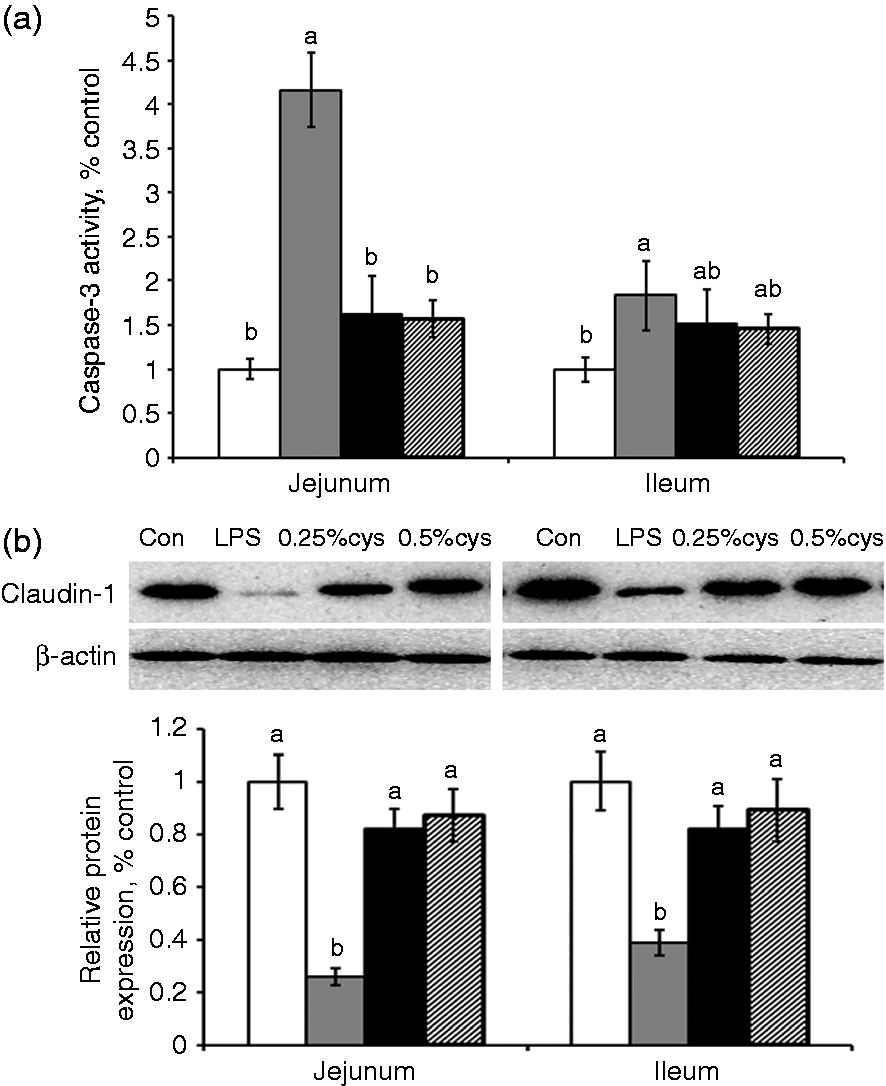
As shown in Figure 2, the LPS group had higher caspase-3 activity and lower PCNA expression in jejunum and ileum compared with the Con group (P < 0.05). L-Cys supplementation (0.25% or 0.5%) decreased caspase-3 activity in jejunum and improved PCNA expression in jejunum and ileum (P < 0.05) compared with the LPS group.
Effects of L-cys supplementation on (A) caspase-3 activity and (B) PCNA expression in weaned piglets in jejunum and ileum mucosa of weaned piglets. Values are means and SE represented by vertical bars. a,bMeans with different letters differ significantly (P < 0.05). Control group (Con; white bars) was fed with basal diet and injected 0.9% sterile saline; LPS group (LPS; dark gray bars) was fed with basal diet and injected with LPS; LPS + 0.25% L-cys group (0.25% cys; black bars) was fed with a 0.25% L-cys-supplemented diet and injected with LPS; LPS + 0.5% L-cys group (0.5% cys; hatched bsars) was fed with a 0.5% L-cys-supplemented diet and injected with LPS. The control sample was used as the reference sample. The protein expression of all samples was expressed as fold changes, calculated relative to Con group.
Inflammation
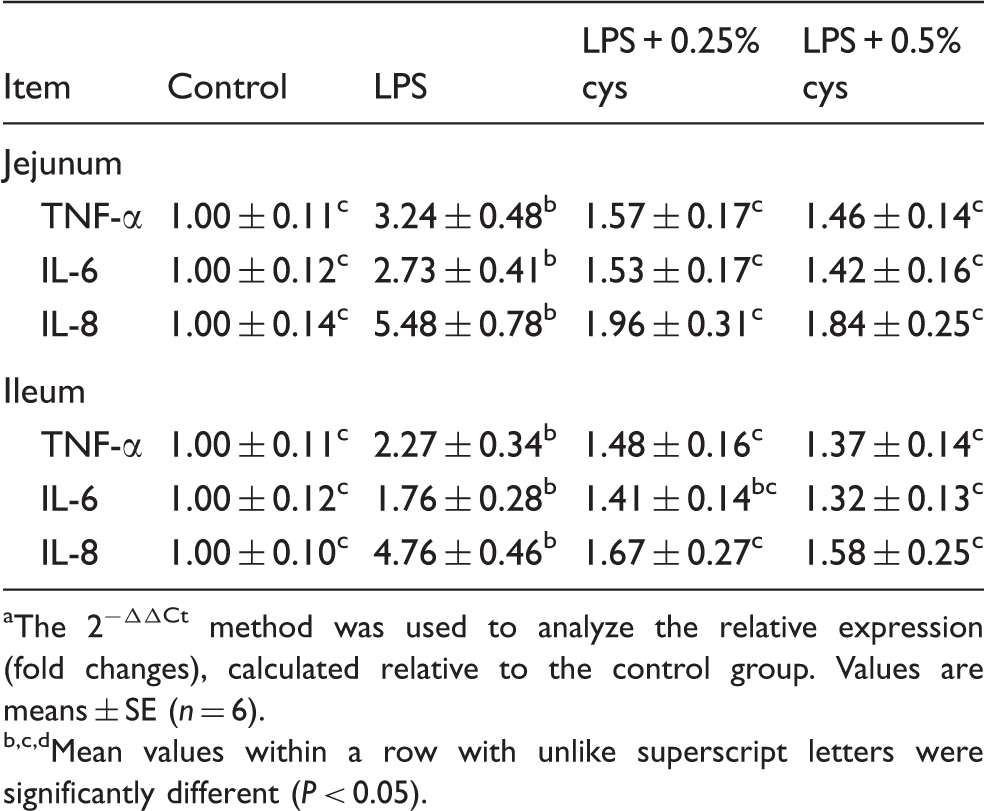
The effect of L-cys supplementation on inflammatory cytokine expression in the jejunum and ileum of weaned piglets are presented in Table 3. Relative to the Con group, LPS challenge improved TNF-α, IL-6 and IL-8 expression in jejunum and ileum of piglets (P < 0.05). Supplementation with 0.25% L-cys reduced TNF-α, IL-6 and IL-8 expression in jejunum, and TNF-α and IL-8 expression in ileum compared with the LPS group (P < 0.05). Supplementation with 0.5% L-cys reduced TNF-α, IL-6 and IL-8 expression in jejunum and ileum (P < 0.05).
Effects of L-cys supplementation on the expression of TNF-α, IL-6 and IL-8 in weaned piglets.a
Item
Control
LPS
LPS + 0.25% cys
LPS + 0.5% cys
Jejunum
TNF-α
1.00 ± 0.11c
3.24 ± 0.48b
1.57 ± 0.17c
1.46 ± 0.14c
IL-6
1.00 ± 0.12c
2.73 ± 0.41b
1.53 ± 0.17c
1.42 ± 0.16c
IL-8
1.00 ± 0.14c
5.48 ± 0.78b
1.96 ± 0.31c
1.84 ± 0.25c
Ileum
TNF-α
1.00 ± 0.11c
2.27 ± 0.34b
1.48 ± 0.16c
1.37 ± 0.14c
IL-6
1.00 ± 0.12c
1.76 ± 0.28b
1.41 ± 0.14bc
1.32 ± 0.13c
IL-8
1.00 ± 0.10c
4.76 ± 0.46b
1.67 ± 0.27c
1.58 ± 0.25c
The 2−ΔΔCt method was used to analyze the relative expression (fold changes), calculated relative to the control group. Values are means ± SE (n = 6).
Mean values within a row with unlike superscript letters were significantly different (P < 0.05).
Oxidative stress
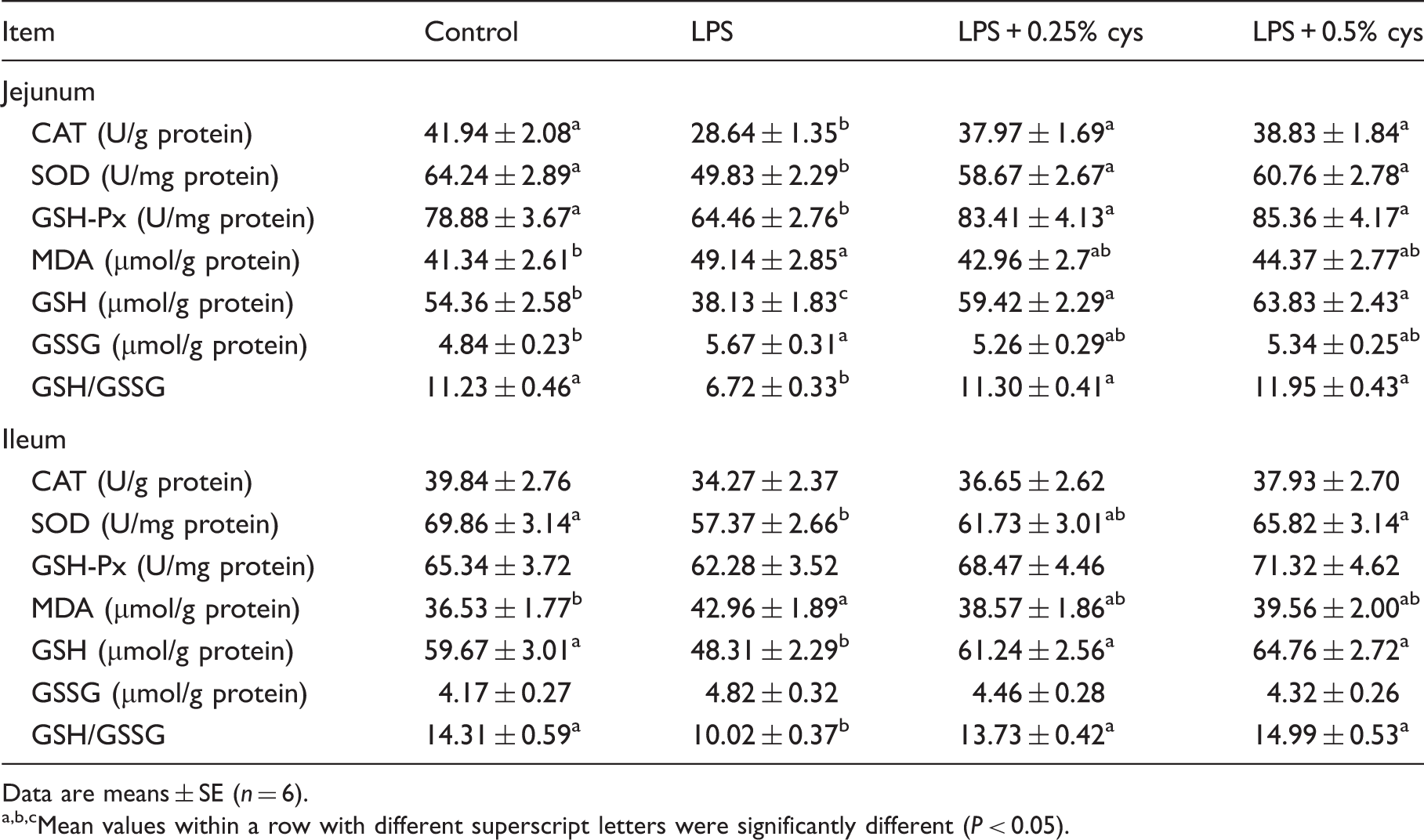
As shown in Table 4, compared with the Con group, LPS challenge increased MDA and GSSG content; decreased CAT, SOD and GSH-Px activity, and GSH contents and GSH/GSSG ratio in jejunum; increased MDA activity; and decreased SOD activity, GSH and GSH/GSSG ratio in ileum (P < 0.05). Relative to the LPS group, piglets in the LPS + 0.25% L-cys group had higher CAT, SOD and GSH-Px activity, GSH concentration and GSH/GSSG ratio in jejunum, and higher GSH concentration and GSH/GSSG in ileum (P < 0.05). Piglets in the LPS + 0.5% L-cys group had higher CAT, SOD and GSH-Px activity, GSH concentration and GSH/GSSG in jejunum, and higher SOD activity, GSH concentration and GSH/GSSG in ileum (P < 0.05).
Effect of L-cys supplementation on antioxidant enzymes, GSH, GSSG and MDA of weaned pigs in jejunum and ileum.
Item
Control
LPS
LPS + 0.25% cys
LPS + 0.5% cys
Jejunum
CAT (U/g protein)
41.94 ± 2.08a
28.64 ± 1.35b
37.97 ± 1.69a
38.83 ± 1.84a
SOD (U/mg protein)
64.24 ± 2.89a
49.83 ± 2.29b
58.67 ± 2.67a
60.76 ± 2.78a
GSH-Px (U/mg protein)
78.88 ± 3.67a
64.46 ± 2.76b
83.41 ± 4.13a
85.36 ± 4.17a
MDA (µmol/g protein)
41.34 ± 2.61b
49.14 ± 2.85a
42.96 ± 2.7ab
44.37 ± 2.77ab
GSH (µmol/g protein)
54.36 ± 2.58b
38.13 ± 1.83c
59.42 ± 2.29a
63.83 ± 2.43a
GSSG (µmol/g protein)
4.84 ± 0.23b
5.67 ± 0.31a
5.26 ± 0.29ab
5.34 ± 0.25ab
GSH/GSSG
11.23 ± 0.46a
6.72 ± 0.33b
11.30 ± 0.41a
11.95 ± 0.43a
Ileum
CAT (U/g protein)
39.84 ± 2.76
34.27 ± 2.37
36.65 ± 2.62
37.93 ± 2.70
SOD (U/mg protein)
69.86 ± 3.14a
57.37 ± 2.66b
61.73 ± 3.01ab
65.82 ± 3.14a
GSH-Px (U/mg protein)
65.34 ± 3.72
62.28 ± 3.52
68.47 ± 4.46
71.32 ± 4.62
MDA (µmol/g protein)
36.53 ± 1.77b
42.96 ± 1.89a
38.57 ± 1.86ab
39.56 ± 2.00ab
GSH (µmol/g protein)
59.67 ± 3.01a
48.31 ± 2.29b
61.24 ± 2.56a
64.76 ± 2.72a
GSSG (µmol/g protein)
4.17 ± 0.27
4.82 ± 0.32
4.46 ± 0.28
4.32 ± 0.26
GSH/GSSG
14.31 ± 0.59a
10.02 ± 0.37b
13.73 ± 0.42a
14.99 ± 0.53a
Data are means ± SE (n = 6).
Mean values within a row with different superscript letters were significantly different (P < 0.05).
NF-kB and Nrf2
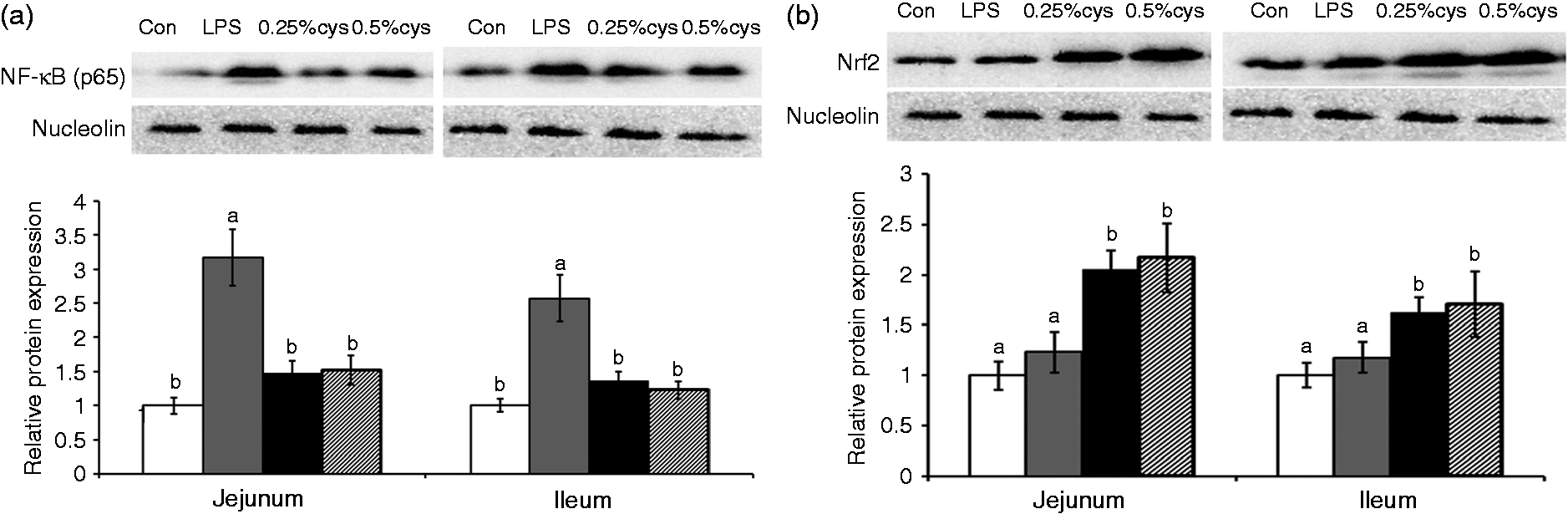
The nuclear translocation of transcription factor NF-κB and Nrf2 is presented in Figure 3. Relative to the Con group, the LPS group had higher NF-κB nuclear translocation in jejunum and ileum (P < 0.05) but no significant difference on Nrf2 nuclear translocation (P > 0.05). Dietary addition of L-cys (0.25% or 0.5%) suppressed (P < 0.05) the NF-κB nuclear translocation and elevated (P < 0.05) the Nrf2 translocation compared with jejunum and ileum in the LPS group.
Effects of L-cys supplementation on the nuclear translocation of transcription factors (A) NF-κB (p65) and (B) Nrf2 in jejunum and ileum mucosa of weaned piglets. Values are means and SE represented by vertical bars. a,bMeans with different letters differ significantly (P < 0.05). Control group (Con; white bars) was fed with basal diet and injected 0.9% sterile saline; LPS group (LPS; dark gray bars) was fed with basal diet and injected with LPS; LPS + 0.25% L-cys group (0.25% cys; bars) was fed with a 0.25% L-cys-supplemented diet and injected with LPS; LPS + 0.5% L-cys group (0.5% cys; hatched bsars) was fed with a 0.5% L-cys-supplemented diet and injected with LPS. The control sample was used as the reference sample. The protein expression of all samples was expressed as fold changes, calculated relative to Con group.
Discussion
In the present experiment, dietary addition of 0.25% and 0.5% L-cys did not affect the growth performance of weaned piglets before LPS challenge. The Cys used in this study was L-cys. Different from our results, Kim et al. reported that infusion 0.21% L-cys significantly improved mass gain to feed intake ratio compared with DSS-challenged piglets.9 Hou et al. reported that dietary addition of N-acetyl cysteine (NAC) attenuates LPS-induced mass loss in weaned piglets.13 However, Lee et al. found that dietary supplemented with 2% L-cys decreased the food intake and BM of rats receiving a normal diet (contained 0.3% L-cys).10 Dilger et al. reported that excess Cys consumption (40 g/d) severely depressed the weight gain of young pigs.17 The reason for this discrepancy may result from different factors, such as animal species, the L-cys level supplemented and the cysteine source (L-cys and NAC).The range of L-cys concentration in the regular diet of weaned piglets is 0.35–0.47%, according to National Research Council swine requirements. L-cys is the free form of cysteine, and NAC is the acetylated variant of L-cys. In the animal body, NAC is converted to cysteine via a deacetylation reaction carried out by several aminoacylases. NAC, as the precursor of L-cys, is now widely used in human and animals as an antioxidant. However, the efficacy of NAC may be limited because not all tissues contain the deacetylases required to convert NAC to cysteine. Therefore, L-cys has recently been explored as an alternative to NAC. It was used as its metabolism and absorption presents a minimal amount of cellular stress, and does not require digestion or ATP expenditure prior to being absorbed. Thus, cys would be rapidly absorbed and not exert additional strain on the already compromised cells.
The integrity of intestine plays an essential role in nutrition absorption, gut homeostasis and animal growth and health.1,2,18 VH, CD and VH:CD ratio are the common indices for evaluating intestinal morphology.5 In our present study, LPS challenge reduced VH, VH/CD in jejunum and ileum, and increased CD in jejunum. These results demonstrated that LPS caused acute intestinal damage as after the injury there was an elongation of the crypt to repair the mucosal damage.219 In current experiment, piglets fed with L-cys supplemented diet showed higher VH and VH/CD ratio in jejunum and ileum and lower CD in jejunum, which indicated that L-cys effectively attenuated LPS-induced intestinal damage. Consistent with the intestinal morphology results, piglets fed with L-cys supplemented diet showed higher TER and lower FD4 flux in jejunum and ileum, which indicated an improvement in intestinal barrier function. Similarly, Kim et al. found piglets fed with L-cys had a lower plasma D-mannitol concentration and improved restoration of the intestinal barrier integrity.9 Hou et al. found that dietary supplemented with NAC decreased crypt depth, increased VH:CD ratio and decreased diamine oxidase activity in plasma of LPS-challenged weanling piglets.13 In the present study, dietary L-cys supplementation was effective in protecting intestinal integrity.
Intestinal epithelial barrier primarily consists of epithelial cells and the tight junctions between epithelial cells.20 Tight junction proteins mainly include the integral transmembrane proteins, such as the occludin and claudin families, and intracellular linker proteins, like zonula occludens.21 These proteins construct a complex formation and seal the intercellular space between epithelial cells, working as a selective permeable barrier.22 Disruption of tight junctions would increase the intestinal permeability, which has been recognized as the major etiology of many intestinal diseases.23,24 In the present experiments, L-cys improved the expression of occludin and claudin-1 in jejunum and ileum. Numerous studies have reported that occludin and claudin-1 are key component of tight junction complexes and play important roles in intestinal permeability.25,26 Similarly to our results, it has been reported that dietary supplementation with NAC increased the expression of claduin-1 and occludin in jejunum of LPS-challenged pigs and increased claudin-1 expression in colon of acetic acid-challenged piglets.13,25 These results suggest that L-cys may alleviate the LPS-induced intestinal damage via regulating expression of tight junction proteins.
Epithelial turnover, including cell proliferation and apoptosis, is another critical factor determining intestinal integrity.9 In inflamed intestinal mucosa of IBD patients, normal intestinal epithelial cell turnover is disrupted.27 Epithelial cell apoptosis increase and proliferation decrease has been thought to be the main mechanism responsible for intestinal mucosal damage. Caspase-3 and PCNA are common markers to assess cell apoptosis and proliferation. In the current study, we found that LPS challenge enhanced caspase-3 activity and reduced PCNA expression in jejunal and ileal mucosa of weaned piglets. Our results were similar to those of Leng et al.28 In our current study, supplementation with L-cys reduced caspase-3 activity and improved PCNA expression. In support of our results, Salman et al. found that L-cys administration reduced caspase-3 concentrations in the liver of streptozotocin-induced type 2 diabetes rats.6 Zhu et al. reported that NAC decreased enterocyte apoptosis index and inhibited Fas, caspase-3 and caspase-8 gene expression in weanling piglets.29 These results suggested that L-cys may protect intestinal integrity by regulating epithelial cell apoptosis and proliferation, although the mechanism of cys regulation of cell proliferation and apoptosis has not been fully elucidated. Shoveller et al. reported that cys regulates epithelial cell proliferation via modulation of redox status.7 Increased oxidant stress and redox imbalance suppress cell proliferation and induce apoptosis. Cysteine may regulate cellular redox by its thiol (-SH) and as a precursor of GSH, a major cellular antioxidant.7
To further explore the mechanism of L-cys protection of the intestinal barrier integrity, we investigated its effects on intestinal inflammation. Accumulating evidence has demonstrated that inflammation play a critical role in many intestinal diseases.5 Several relatively pro-inflammatory cytokines including TNF-α and IL-1β, have been found to disrupt the tight junction complex and lead to epithelial cell apoptosis.23,24 In our experiment, LPS challenged improved TNF-α, IL-6, and IL-8 concentration in jejunum and ileum of piglets. LPS-induced model of gut inflammation is a most common used and well-established intestinal injury model. When binding with TLR4/MD-2/CD14 receptor complex, LPS stimulates immune cells, such as macrophages, to produce a variety of pro-inflammatory cytokines, such as TNF-α, IL-6 and IL-8. Piglets fed with L-cys-supplemented diets had lower TNF-α, IL-6, IL-8 concentration in jejunum and ileum. Similar with our results, Kim et al. showed that L-cys administration suppressed intestinal inflammation, decreased TNF-α, IL-1β and IL-6 expression in the intestine of a DSS-induced porcine colitis model.9L-cys was also found to reduce inflammatory markers MCP-1and CRP levels in serum in diabetes rats.6In vitro studies showed that supplementation with L-cys reduced TNF-α, IL-8 and IL-1β expression in human U937 monocytes treated with high Glc.30 Therefore, in the present study, L-cys may protect the intestinal integrity by inhibiting intestinal inflammation.
Oxidative stress is another critical factor that involved in the intestinal barrier disruption in intestinal diseases.18 ROS are not only byproducts of the inflammatory process; they also are capable of causing damage to epithelial cell integrity and closely linked to apoptosis in a variety of cell types.3 Evidences with intestinal epithelial cells has proved that elevated oxidant stress could inhibit cell proliferation and induce apoptosis.3,31 In animals, there exist many anti-oxidative enzymes (e. g. CAT, SOD, GSH-Px) and non-enzymatic antioxidants (e.g. GSH) to protect the organism from oxidative damage.32 MDA is produced by lipid peroxidation and often regard as the indices for oxidative injury. In the current study, LPS challenge elevated MDA and GSSG contents and decreased CAT, SOD, GSH-Px activity, GSH contents and GSH/GSSG. Similarly, Hou et al. reported that NAC attenuated LPS-induced oxidative stress in intestine as indicated by improved CAT, SOD and GSH-Px activity, GSH contents and GSH/GSSG in small intestine of weaned piglets.13 Lee et al. reported that L-cys supplementation improved catalase and GSH reductase activity in rats.10 Jain et al. found that L-cys administration attenuates oxidative stress in Zucker diabetic rat liver.11In vitroL-cys reduced high Glc- and ketone-induced ROS production in human umbilical vein endothelial cells and THP-1 monocytes.8 These results indicate that L-cys may protect intestinal integrity via attenuation of oxidative stress.
According to our results, L-cys alleviated the LPS-induced intestinal disturbance via its effects on suppressing inflammation and oxidant stress. To elucidate fully the mechanism of L-cys protect the intestinal barrier, we examined two critical signaling pathway that are involved in anti-inflammation and antioxidant, NF-κB and Nrf2 signaling pathways. NF-κB and Nrf2 pathways form complicated signaling nets to regulate the intestinal inflammation and redox status.18 Once stimulated, NF-kB up-regulates various pro-inflammatory cytokines expression, including TNF-α, IL-1β, IL-6 and IL-8,33 while Nrf2 improves the expression of antioxidant genes and inhibits the expression of pro-inflammatory cytokines.34 In our current experiments, dietary addition of L-cys inhibited NF-κB nuclear translocation and elevated Nrf2 translocation. Consistent with our results, Manna and Jain reported that supplementation with L-cys reduced NF-kB phosphorylation in high Glc-treated human U937 monocytes.30 Satsu et al. reported that supplementation with cys improved Nrf2 protein nucleus translocation in human intestinal epithelial LS180 cells.34 Yang et al. reported that L-cys up-regulated Nrf2 and downstream NQO1 and HO-1 mRNA expression in rats.35 Many studies have shown that L-cys can regulate NF-κB and Nrf2 signaling pathway in multiple ways. Several studies reported that L-cys may regulate the NF-κB and Nrf2 signaling pathway by affecting cellular thiols.36–38 Intracellular thiols could regulate NF-KB signaling pathway at one or more points. For instance, high intracellular thiol levels could influence protein folding or enzyme activation and thus inhibit the activation of the protein kinases that phosphorylate the IKB/NF-κB complex and liberate activated NF-κB. Alternatively, high thiols could interfere directly with IKB phosphorylation or with the transport of activated NF-κB into the nucleus. Qanungo et al. demonstrated that NAC could prevent NF-κB activation via S-glutathionylation of p65-NFκB.39 Moreover, L-cys also has been reported to block NF-κB activation by affecting the upstream NF-kB signaling pathway. Wang et al. reported that NAC down-regulated TLR4 and type I transmembrane receptors that inhibited NF-kB activation.125 Finally, L-cys can regulate the NF-κB and Nrf2 signaling pathway simply by scavenging oxidants, which can also directly activate NF-κB and Nrf2 signaling pathway. Thus, in the present study, the beneficial effects of L-cys on intestinal integrity may be related to alleviating intestinal inflammation and oxidative stress via regulating NF-κB and Nrf2 signaling pathway.
In conclusion, L-cys supplementation exhibits beneficial effects on improving intestinal integrity. Its protective effects on the intestine may be closely related to improving tight junction protein expression, inhibiting apoptosis and improving proliferation of epithelial cell, alleviating intestinal inflammation and oxidative stress via inhibiting the NF-κB signaling pathway and activating the Nrf2 signaling pathway.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the National Natural Science Foundation of China (31472103), the Special Fund for Agroscientific Research in the Public Interest (201403047).
References
1.
WijttenPJMeulenJVVerstegenMW. Intestinal barrier function and absorption in pigs after weaning: a review. Br J Nutr2011; 105: 967–981.
2.
BlikslagerATMoeserAJGookinJL. Restoration of barrier function in injured intestinal mucosa. Physiol Rev2007; 87: 545–564.
3.
OzHSChenTSNagasawaH. Comparative efficacies of 2 cysteine prodrugs and a glutathione delivery agent in a colitis model. Transl Res2007; 150: 122–129.
4.
OzHSChenTSMcClainCJ. Antioxidants as novel therapy in a murine model of colitis. J Nutr Biochem2005; 16: 297–304.
5.
LiuYChenFOdleJ. Fish oil enhances intestinal integrity and inhibits TLR4 and NOD2 signaling pathways in weaned pigs after LPS challenge. J Nutr2012; 142: 2017–2024.
6.
SalmanZKRefaatRSelimaE. The combined effect of metformin and l-cysteine on inflammation, oxidative stress and insulin resistance in streptozotocin-induced type 2 diabetes in rats. Eur J Pharmacol2013; 714: 448–455.
7.
ShovellerAKStollBBallROBurrinDG. Nutritional and functional importance of intestinal sulfur amino acid metabolism. J Nutr2002; 132: 1609–1611.
8.
Kanikarla-MariePJainSK. l-Cysteine supplementation reduces high-glucose and ketone-induced adhesion of monocytes to endothelial cells by inhibiting ROS. Mol Cell Biochem2014; 391: 251–256.
9.
KimCJKovacs-NolanJYangC. L-cysteine supplementation attenuates local inflammation and restores gut homeostasis in a porcine model of colitis. Biochem Biophys Acta2009; 1790: 1161–1169.
10.
LeeSHanKNakamuraY. Dietary L-cysteine improves the antioxidative potential and lipid metabolism in rats fed a normal diet. Biosci Biotech Biochem2014; 77: 1430–1134.
11.
JainSKVelusamyTCroadJL. L-Cysteine supplementation lowers blood glucose, glycated hemoglobin, CRP, MCP-1, and oxidative stress and inhibits NF-κB activation in the livers of Zucker diabetic rats. Free Radical Bio Med2009; 46: 1633–1638.
12.
National Research Council. Nutrient requirements of swine, 10th ed. Washington, DC: National Academy Press, 1998.
13.
HouYWangLZhangW. Protective effects of N-acetylcysteine on intestinal functions of piglets challenged with lipopolysaccharide. Amino Acids2012; 43: 1233–1242.
14.
SAS. SAS/STAT user’s guide, Version 8.1, Cary, NC: SAS Institute Inc., 2000.
15.
BakerDH. Comparative species utilization and toxicity of sulfur amino acids. J Nutr2006; 136: 1633S–1635S.
16.
PasiniEAquilaniRDioguardiFS. Amino acids: chemistry and metabolism in normal and hypercatabolic states. Am J Cardiol2004; 93(Suppl.): 3A–5A.
17.
DilgerRNToueSKimuraT. Excess dietary L-cysteine, but not L-cystine, is lethal for chicks but not for rats or pigs. J Nutr2007; 137: 331–338.
18.
CircuMLAwTY. Intestinal redox biology and oxidative stress. Semin Cell Dev Biol2012; 23: 729–737.
19.
WilliamsKLFullerCRDielemanLA. Enhanced survival and mucosal repair after dextran sodium sulfate-induced colitis in transgenic mice that overexpress growth hormone. Gastroenterology2001; 120: 925–937.
20.
HuCHXiaoKLuanZSSongJ. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J Anim Sci2013; 91: 1094–1101.
21.
HuCSongJLiY. Diosmectite-zinc oxide composite improves intestinal barrier function, modulates expression of pro-inflammatory cytokines and tight junction protein in early weaned pigs. Br J Nutr2013; 110: 681–688.
22.
LiXAkhtarSChoudhryMA. Alteration in intestine tight junction protein phosphorylation and apoptosis is associated with increase in IL-18 levels following alcohol intoxication and burn injury. Biochim Biophys Acta2012; 1822: 196–203.
23.
Al-SadiRMMaTY. IL-1beta causes an increase in intestinal epithelial tight junction permeability. J Immunol2007; 178: 4641–4649.
24.
MaTYBolvinMAYeD. Mechanism of TNF-α modulation of Caco-2 intestinal epithelial tight junction barrier: role of myosin light-chain kinase protein expression. Am J Physiol Gastrointest Liver Physiol2004; 288: G422–G430.25.
25.
WangQHouYYiD. Protective effects of N-acetylcysteine on acetic acid-induced colitis in a porcine model. BMC Gastroenterol2013; 13: 133–133.
26.
Van ItallieCMFanningASHolmesJAndersonJM. Occludin is required for cytokine-induced regulation of tight junction barriers. J Cell Sci2010; 123: 2844–2852.
27.
PastorelliLDe SalvoCMercadoJR. Central role of the gut epithelial barrier in the pathogenesis of chronic intestinal inflammation: lessons learned from animal models and human genetics. Front Immunol2013; 4: 280–280.
28.
LengWLiuYShiH. Aspartate alleviates liver injury and regulates mRNA expressions of TLR4 and NOD signaling-related genes in weaned pigs after lipopolysaccharide challenge. J Nutr Biochem2014; 25: 592–599.
29.
ZhuLCaiXGuoQ. Effect of N-acetyl cysteine on enterocyte apoptosis and intracellular signalling pathways' response to oxidative stress in weaned piglets. Br J Nutr2013; 110: 1938–1947.
30.
MannaPJainSK. L-cysteine and hydrogen sulfide increase PIP3 and AMPK/PPARγ expression and decrease ROS and vascular inflammation markers in high glucose treated human U937 monocytes. J Cell Biochem2013; 114: 2334–2345.
31.
JonasCRZieglerTRGuLHJonesDP. Original contribution extracellular thiol/disulfide reds proliferation rate in a human colon carcinoma (Caco2) cell line. Free Radical Bio Med2002; 33: 1499–1506.
32.
YinJLiuMRenW. Effects of dietary supplementation with glutamate and aspartate on diquat-Induced oxidative stress in piglets. PLoS One2015; 10: e122893–e122893.
33.
HuCHSongZHXiaoK. Zinc oxide influences intestinal integrity, the expressions of genes associated with inflammation and TLR4-myeloid differentiation factor 88 signaling pathways in weanling pigs. Innate Immun2014; 20: 478–486.
34.
SatsuHChidachiEHiuraY. Induction of NAD(P)H:quinone oxidoreductase 1 expression by cysteine via Nrf2 activation in human intestinal epithelial LS180 cell. Amino Acids2012; 43: 1547–1555.
35.
YangLShenJHeS. L-cysteine administration attenuates pancreatic fibrosis induced by TNBS in rats by inhibiting the activation of pancreatic stellate cell. PLoS One2012; 7: e31807–e31807.
36.
YangLJShenJQHeSS. L-Cysteine administration attenuates pancreatic fibrosis induced by TNBS in rats by inhibiting the activation of pancreatic stellate cell. PLoS One2012; 7: e31807–e31807.
37.
AlessandraFRitaCAnnaC. Molecules altering the intracellular thiol content modulate NF-kB and STAT-1/IRF-1 signaling pathways and IL-12 p40 and IL-27 p28 production in murine macrophages. PLoS One2013; 8: e57866.–e57866..
38.
FrankJMarioRLeonardAHLeonoreAH. Intracellular thiols regulate activation of nuclear factor KCB and transcription of human immunodeficiency virus. Proc Natl Acad Sci U S A1990; 87: 9943–9947.
39.
QanungoSJoachimDManevichY. N-acetyl-l-cysteine sensitizes pancreatic cancers to gemcitabine by targeting the NFκB pathway. Biomed Pharmacother2014; 68: 855–864.