
Abstract
NF-κB, which comprises homo- and hetero-dimers of the five members of the Rel family, plays a crucial role in immunity to infection. The cRel and p50 subunits have been implicated in the development and function of the immune cells, but their in vivo importance remains poorly explored in sepsis. We aimed to study the impact of the combined loss of these two subunits on the innate response to infection in a cecal ligation and puncture model of sepsis. We have explored the possible defects in host defense, including pathogen clearance, bacterial phagocytosis and cytokine plasma release. We also performed gene profiling of cRel−/−p50−/− and wild-type LPS-stimulated peritoneal macrophages. Deficiency of cRel and p50 led to enhanced mortality to sepsis that was associated with defective macrophages phagocytosis, decreased bacterial clearance and moderate cytokine response. Transcription profile analysis revealed a common inflammatory response but a significant down-regulated transcription of genes encoding for pathogen recognition receptors and antimicrobial molecules, supporting the in vivo findings in mice. In conclusion, the cRel and p50 subunits of NF-κB play an important combined role in the innate response and are crucial for survival and pathogen clearance in polymicrobial sepsis.
Introduction
Upon infection, the host initiates an innate inflammatory response mediated through an early recognition of pathogen-associated molecular patterns (PAMPs) of the invading microorganisms by mobilized phagocytes, such as macrophages. 1 Those cells detect invading pathogens by different pathogen recognition receptors (PRRs) and are engulfed by phagocytosis and killed.2–4
PAMP stimulation of innate immune cells during sepsis engages the classical NF-κB pathway, an early step necessary for host defense, as well as inflammation and the induction of adaptive immunity.5,6 NF-κB factors consist of homo- and heterodimers of the five members of the Rel protein family: RelA (p65), RelB, cRel, p50 and p52. 7 Among the 15 potential dimer associations, 12 appear to be transcriptionally active, but only 3 have been extensively studied. 8 Amongst these, the heterodimer RelA:p50 is a key factor for pro-inflammatory gene transcription and the regulation of apoptosis.7,9 In contrast, the p50:p50 homodimers appear not only to repress the transcription of genes encoding pro-inflammatory molecules such as TNF-α and IL-12, but can also stimulate the transcription of the gene for the anti-inflammatory cytokine IL-10.10,11 Finally, the cRel:p50 heterodimers seem to be important in the development and control of adaptive immunity mediated through the regulation of co-stimulatory molecule expression and the control of dendritic cell survival.6,7,9,12
The relative importance of NF-κB subunits in sepsis has been highlighted mainly in knockout mice studies. 13 Despite an apparent normal development, cRel−/− and p50−/− mice exhibit several immunological defects in lymphocyte proliferation and humoral immunity.14,15 Both strains are susceptible to various single microbial infections,15–18 while an increased susceptibility of the cRel−/− mice to polymicrobial sepsis has recently been reported. 19 As expected, the combined loss of cRel and p50 [double knockout (DKO)] not only leads to an increased severity of defects seen in the individual knockout mice, but also the emergence of novel phenotypes, such as a reduced number of plasmacytoid dendritic cells,20,21 a decreased survival of conventional dendritic cells and impaired IL-12 production. 22 The DKO mice also exhibit decreased T cell receptor-induced T cell survival and impaired T cell responses, as well as low numbers of effectors/memory and regulatory CD4+ T cells. 23 However, the combined importance of cRel and p50 in innate immunity is poorly studied. In particular, the in vivo importance of cRel/p50 subunits in a polymicrobial model of sepsis is still unknown.
Here, we report that DKO mice have decreased bacterial clearance linked to impaired phagocytosis and an increased rate of mortality in polymicrobial sepsis. Gene profiling of LPS-stimulated peritoneal macrophages revealed dramatic diminished expression of PRR transcripts in DKO cells, supporting the in vivo findings in mice in the cecal ligature and puncture (CLP) model of sepsis and highlighting the important role of cRel and p50 subunits in innate immunity.
Materials and methods
Mice
C57BL6/J female mice, 8–12 wk old, were purchased from Charles River Laboratories (Orleans, France).
p50−/−cRel−/− (DKO) mice with a C57BL6/J background were kindly provided by A. Beg (Moffitt Cancer Center, Tampa, FL, USA). Experiments were conducted in accordance with Cochin Institute guidelines in compliance with European animal welfare regulation.
CLP model of sublethal polymicrobial sepsis
We used a sublethal model of CLP described previously. 19 Briefly, mice were anesthetized by an intraperitoneal (IP) injection of ketamine and xylazine. After a midline incision (<1 cm), the cecum was exposed, ligatured at its external third and punctured through and through with a 21-gauge needle. Incision was sutured in layers and animals were resuscitated with an IP injection of 1 ml saline. Controls were sham-operated mice undergoing abdominal surgery with exposition of cecum without CLP. Six h following surgery and then every 12 h over 3 d, mice received an IP injection of antibiotics (Imipenem cilastatine, Tienam®, Merck Sharp & Dohme (Clermont-Ferrand, France): 25 mg/kg in 0.5 ml saline). Survival was monitored every 12 h for up to 8 d after CLP.
Assessment of bacterial load in liver, spleen and blood
At d 1, 2 and 3 after surgery, the mice were humanely killed and a blood sample was obtained by cardiac puncture. Bacterial dissemination was assessed through quantitative spleen, liver and blood bacterial cultures to quantify the number of CFUs. Briefly, spleen and liver were removed and mechanically homogenized under sterile conditions. Blood, spleen and liver homogenates were subjected to serial 10-fold dilutions. Bacteria were quantified in tryptic soy agar (for spleen and liver homogenates) or in TH agar (for blood) after 24 h.
Escherichia coli phagocytosis experiments
Adherent macrophages were generated by plating the collected peritoneal cells from DKO and wild-type (WT) mice on 12-mm coverslips in 24-well plastic trays at 2 × 105 cells per well during 16 h at 37°C in 5% CO2. Non-adherent cells were removed by washing. GFP-expressing Escherichia coli Xl1blue were grown overnight at 37°C; then, the bacterial suspension absorbance was measured at 600 nm. The bacteria were fixed at 22°C during 15 min with 4% paraformaldehyde and re-suspended in RPMI medium without serum at a multiplicity of infection of 10 before allowing contact with cells for 5 or 60 min at 37°C in 5% CO2. Non-adherent E. coli were then washed and cells were fixed and labeled with rabbit anti-E. coli; all surface antigens were followed by Cy3-anti-rabbit IgG to distinguish internal (GFP-positive, Cy3-negative) and external (GFP- and Cy3-positive) bacteria, and analyzed by a wide-field microscope (Leica DMB, Houston, TX, USA) equipped with an oil immersion objective (100 PL APO HCX, 1.4 NA) and a cooled CCD camera (MicroMAX Princeton Instruments, Acton, MA, USA). Z stacks of fluorescent images were acquired using a piezo with a 0.2-mm increment. Phase contrast images of the same cells were acquired to identify phagosomes. The efficiencies of bacterial association and phagocytosis were calculated for 50 DKO and 50 WT cells randomly chosen on the coverslips. Results are expressed as indexes of association and phagocytosis, i.e. the mean numbers of associated or phagocytosed bacteria per cell.
Determination of cytokine levels in mice sera
At d 0, 1 and 3 after surgery, concentrations of IL-1β, IL-6, TNF-α, monocyte chemotactic protein (MCP)-1, IFN-γ and IL-10 were quantified in mice sera using quantitative multiplex assays (Bio-Plex cytokines; BioRad, Marne la Coquette, France) according to the manufacturer’s instructions.
Microarrays
Resident macrophages were collected from the peritoneal cavity of naive WT and DKO mice with RPMI medium. Peritoneal macrophages (5 × 105 WT and DKO) cultured in RPMI (10% fetal calf serum (FCS), 1% penicillin/streptomycin, 1% HEPES, 1% glutamine) were stimulated for 2, 6 or 18 h with 1 µg/ml LPS (E. coli K-12 ultra-pure LPS, Invivogen, Toulouse, France). RNA extraction was performed with TRIzol® Plus RNA Purification Kit (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol and analyzed using an Agilent 2100 Bioanalyser (Agilent Technologies, Palo Alto, CA, USA). RNA was used to make biotin labeled cRNA using SuperScript® double-stranded cDNA synthesis kit (Invitrogen) and BioArray high yield RNA transcript labeling kit (Enzo, Villeurbanne, France). cDNA were hybridized to GeneChip Mouse Gene 1.0 ST Array (Affymetrix, Santa Clara, CA, USA) with the GeneChip WT Terminal Labeling Kit. The arrays were read by a confocal scanner and were imported into two analysis softwares: MicroArray Suite 5.0 Gene Expression analysis program (Affymetrix) and R Bioconductor. Gene expression levels were calculated using the RMA algorithm in Expression Console and flags were computed using a custom algorithm within R. To limit potentially biased measurement (background or saturating), all probes with normalized and intensity measures outside of confidence interval (CI) were flagged. The CI was two fold SDs from the mean intensities of each chip. Three probe lists have been used for each comparison according to flagged measurement in the relevant chips. The ‘PP’ list was made of probes only flagged as ‘Present’ for all chips involved in the comparison. The ‘P50%’ list was created from filtering probes flagged as ‘Present’ for at least half of the chips. The ‘All’ list consisted of all probes without any filter. Independent samples were compared by computing fold ratios and filtered at twofold. Cluster analysis was performed by hierarchical clustering using the Spearman correlation similarity measure and average linkage algorithm. Data were subsequently submitted to Ingenuity Pathway Analysis (IPA; http://www.ingenuity.com) to model relationships among genes and proteins and to construct putative pathways and relevant biological processes. Micro array data were deposited into ArrayExpress under accession number: E-MEXP-3446.
Quantitative real-time PCR
The cDNA templates were obtained after reverse transcription of peritoneal macrophages RNA using the RevertAid™ H Minus M-MuLV Reverse Transcriptase (Fermentas, Villebon sur Yvette, France). We used the Murine RealTime ready probes (Roche Diagnostics, Meylan, France) of several PRRs, including Tlr1, Marco and Lcn2. Quantitative real-time PCR (qPCR) was performed in triplicate with a 96-well plate Light Cycler480 instrument (Roche Diagnostics) using Roche's Probes Mastermix and 10 ng cDNA per well. Reported fold changes were normalized to hypoxanthine-guanine phosphoribosyltransferase (HPRT) in each sample.
Bone marrow-derived macrophage generation and Western blot
Bone marrow-derived macrophages (BMDMs) were generated upon ex vivo differentiation of mouse bone marrow precursors flushed from femurs and tibias. Bone marrow cells were cultured in six-well plates at 1 × 106 cells/ml in DMEM medium in the presence of 30% supernatant from L929 cells containing macrophage-CSF. Generated macrophages were 95% CD11b+Gr-1−. Protein extracts were made from untreated or 24 h LPS-stimulated WT and DKO BMDMs. Comparative expression of macrophage receptor with collagenous structure (MARCO) between WT and DKO cells was analyzed by Western blot using the specific Ab ED31 for MARCO (Southern Biotech Associates, Birmingham, AL, USA). 24
Statistical analysis
Survival curves were analyzed using the Kaplan-Meier method and compared using the log-rank test. Continuous variables were expressed as median and inter-quartile ranges, displayed using box plots or scatter plots. They were represented as mean ± SD and compared using the Wilcoxon rank sum test. Categorical variables were compared using the Fisher exact test. P values lower than 0.05 indicated statistically significant differences. For microarray analysis, independent samples were compared by computing fold ratios and filtered at twofold. Cluster analysis was performed by hierarchical clustering, using the Spearman correlation similarity measure and average linkage algorithm.
Results
Sublethal polymicrobial sepsis induces early over mortality of cRel−/−p50−/− mice
To investigate the consequences of the combined loss of cRel and p50 subunits of NF-κB on mice outcome to infection, a sublethal polymicrobial sepsis was induced by CLP in WT and DKO mice. Whereas the survival rate of WT mice was 77% (14/18), DKO mice showed an accelerated and statistically higher mortality rate, with only 9% (2/23) of CLP-operated DKO mice surviving after D6 (P < 0.001) (Figure 1). These results clearly indicate that the absence of both cRel and p50 subunits impaired the host survival after a polymicrobial sepsis.
Survival rates of DKO and WT mice in polymicrobial sepsis. Sublethal polymicrobial sepsis was induced by CLP in WT and DKO mice. Survival of WT (continuous line, n = 18) or DKO (doted line, n = 23) post-CLP mice was monitored for 8 d (log rank test P < 0.001).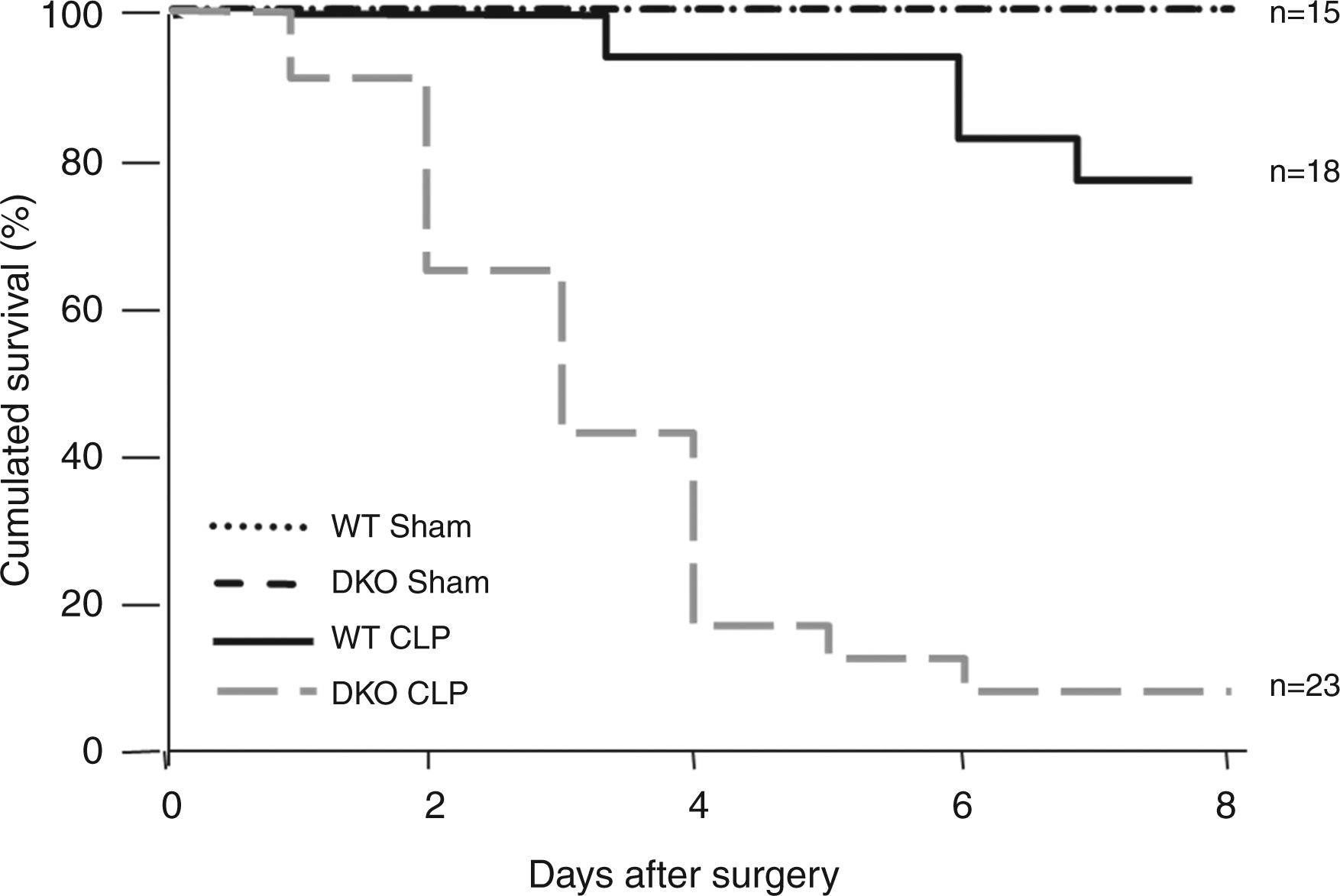
DKO mice exhibit a pathogen clearance defect upon induction of polymicrobial sepsis
Rapid control of pathogen dissemination is essential for host survival to sepsis. To investigate the importance of cRel/p50 subunits on bacterial clearance during sepsis, we evaluated the bacterial burden during the first 3 d following CLP. As shown in Figure 2, at d 1 DKO mice showed a dramatic increase in the amount of bacteria in spleen and liver. While this difference was still pronounced at d 2, by d 3 bacterial numbers in the spleen and liver were equivalent in WT and DKO mice. However, the bacterial burden was significantly higher and sustained in the blood of DKO throughout the three days of infection when compared with WT controls. Therefore, the combined loss of cRel and p50 NF-κB subunits is strongly associated with a defect in bacterial clearance during polymicrobial sepsis.
Assessment of bacterial load in mice upon induction of polymicrobial sepsis. The bacterial count was determined at days 1, 2 and 3 after CLP in DKO (in white) and WT mice (in black). Bacterial dissemination was assessed through quantitative blood, spleen and liver bacterial culture. Differences were determined to be statistically significant by Wilcoxon rank sum test.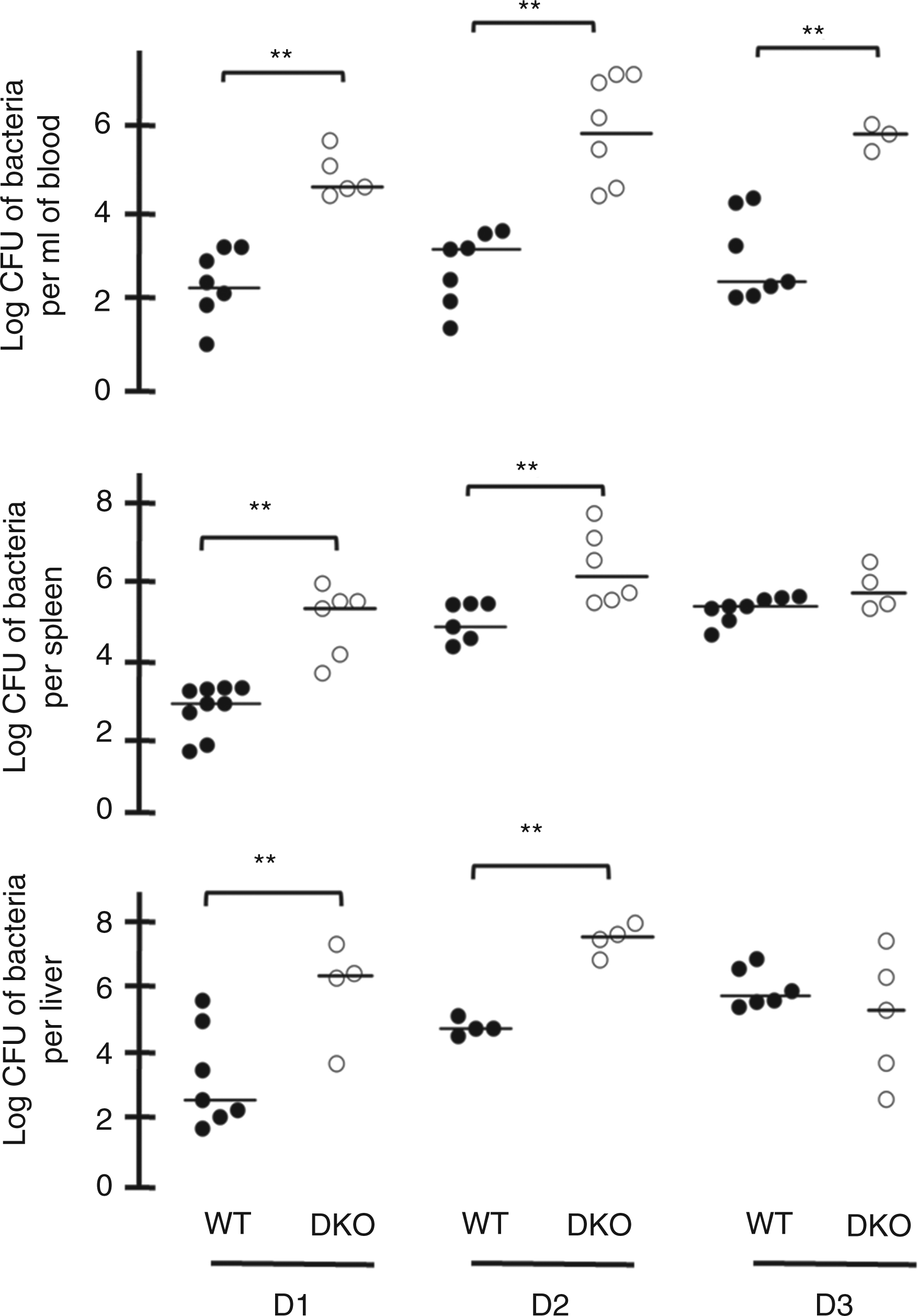
Altered bacterial phagocytosis by cRel−/−p50−/− peritoneal macrophages
During peritonitis, peritoneal macrophages represent the first line of defense in controlling the spread of pathogens. As phagocytosis represents a key bacterial clearance mechanism, we assessed the comparative ability of WT and DKO peritoneal macrophages to perform phagocytosis of non-opsonized, fixed, GFP-expressing E. coli. As illustrated in Figure 3, DKO peritoneal macrophages exhibited a significant defect of both association (Figure 3A) and phagocytosis (Figure 3B) of E. coli when compared with WT cells. These results suggest that the impaired bacterial clearance in DKO mice upon polymicrobial sepsis may be partially explained by the altered macrophage phagocytic ability.
Analysis of E. coli association and phagocytosis by peritoneal macrophages. DKO or WT peritoneal macrophages were incubated for 5 or 60 min with GFP-E. coli at 37°C then fixed and labeled with rabbit anti-E. coli on all surface antigens followed by Cy3-anti-rabbit IgG to distinguish internal (GFP-positive, Cy3-negative) and external (GFP- and Cy3-positive) bacteria. Cells were analyzed by wide-field fluorescence microscopy and phase contrast. The efficiencies of association and phagocytosis were calculated for 50 DKO and 50 WT cells. Results are expressed as an index of association and phagocytosis. The means ± SEM of indices obtained for 100 cells in two independent experiments were plotted.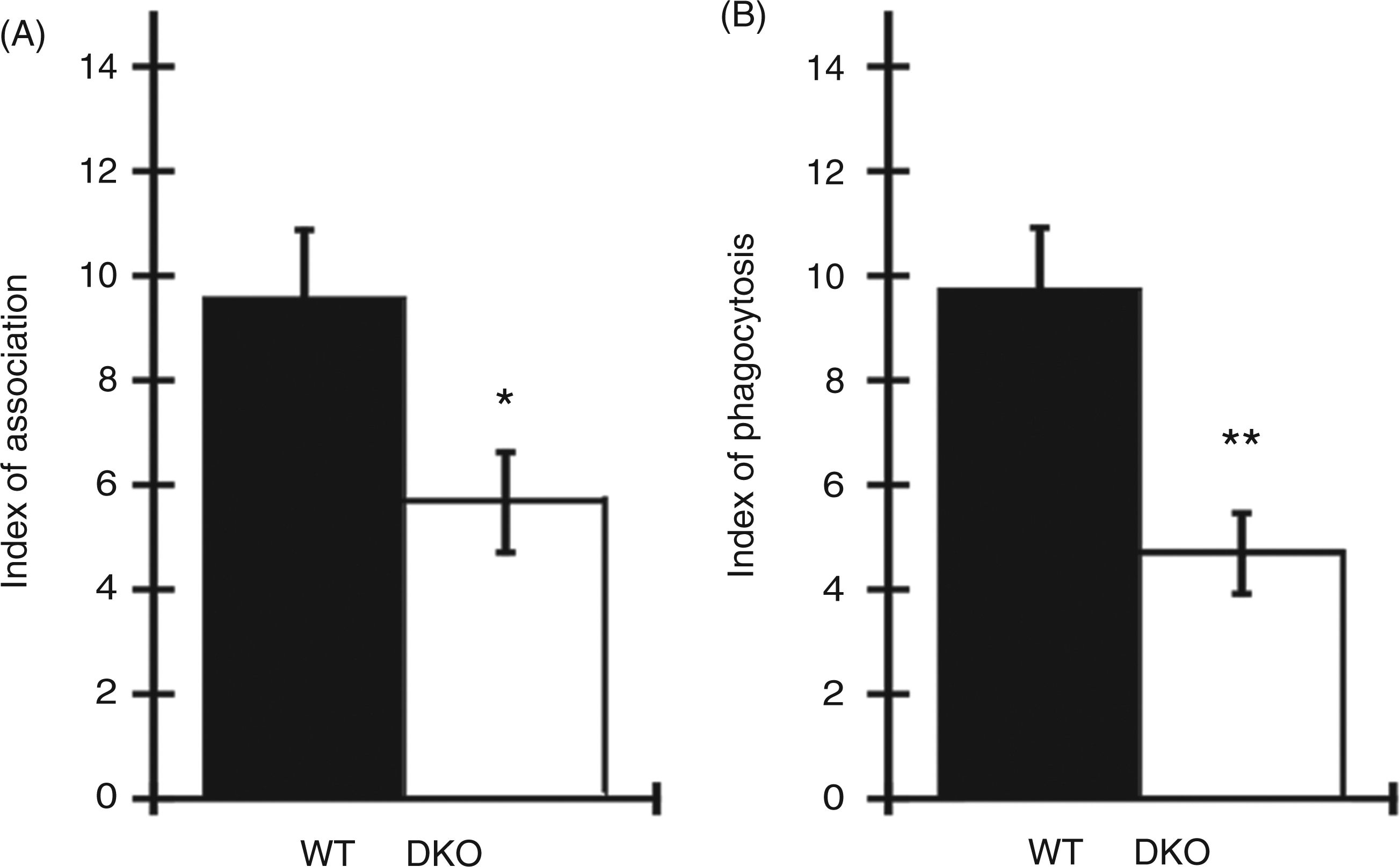
The combined deficiency of cRel and p50 leads to an alteration of the systemic inflammatory response to polymicrobial infection
To explore eventual links between the increased mortality of DKO mice upon polymicrobial sepsis and cytokine release in the plasma, we evaluated the systemic inflammatory response in WT and DKO mice after CLP. Prior to infection, WT and DKO mice exhibited similar plasma levels of cytokines and chemokines (Figure 4). At d 1, DKO mice exhibited a moderate increase in the secretion of IL-1β and TNF-α compared with WT animals, which was counterbalanced by enhanced levels of IL-10. At the same time, IL-6 levels were significantly elevated in the WT mice only. By d 3, the DKO mice displayed an anti-inflammatory profile with a trend towards low levels of pro-inflammatory cytokines, such as IL-1β, TNF-α and IFN-γ, and concomitant high levels of IL-10 and IL-6 compared with WT mice. MCP-1 levels remained unchanged in the WT mice over the course of sepsis but increased significantly at d 3 in the DKO mice.
Assessment of the systemic cytokine response in mice sera after induction of polymicrobial sepsis. Sublethal polymicrobial sepsis was induced by CLP in WT and DKO mice. Blood samples were collected at d 0, 1, and 3 after CLP. The concentration of IL-1β, IL-6, TNF-α, MCP-1, IL-10 and IFN-γ in post-CLP mouse sera was determined by Multiplex. Each measurement value was realized in duplicate; means were obtained from 4–9 mice *: P < 0.05, **: P < 0.01.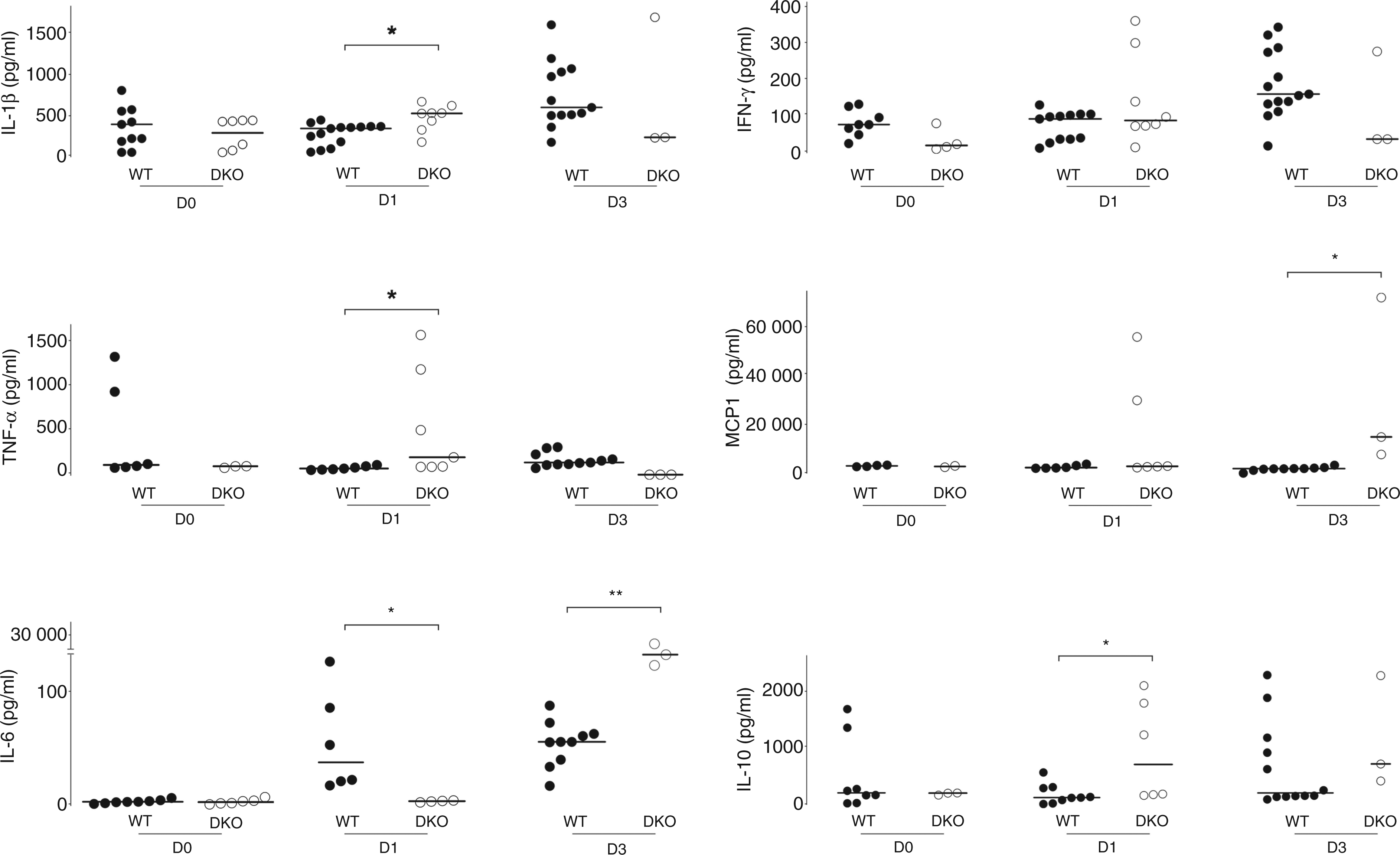
Gene profiling of LPS-stimulated WT and DKO peritoneal macrophages
To further explore the underlying mechanisms of the defective anti-microbial response of the DKO mice, we performed gene profiling of LPS-stimulated peritoneal macrophages, which are crucial cells for innate defense. In WT cells, LPS stimulation led to global expression changes for a total of 1880 probe sets (1583 genes) comprising 882 up-regulated and 998 down-regulated probe sets (Figure 5A). The absence of p50 and cRel led to a higher number of LPS-induced gene expression changes, (2245 probe sets corresponding to 1920 genes), including 1085 up-regulated and 1160 down-regulated probe sets (Figure 5B). However, the increased number of changes in gene expression in DKO cells was not uniform during the time-course of LPS stimulation. At 2 h, endotoxin challenge modified the expression of more genes in the WT cells than in the DKO cells. For both WT and DKO macrophages, the peak of the gene expression changes occurred 6 h after LPS stimulation, with 1309 probe set expression modifications for WT and 1701 variations for DKO cells. These results clearly indicate that quantitatively the absence of both cRel and p50 induces dramatic and complex changes in the transcriptome upon the innate response.
Quantitative analysis of gene modifications induced upon LPS response of WT and DKO peritoneal macrophages at different times of stimulation. The figure represents the number of genes differently expressed (up-regulated named ‘Up’ and down-regulated named ‘Down’, as indicated inside the circles) between the three probe lists in WT macrophages (A) (WT LPS 2 h vs UNT, WT LPS 6 h vs UNT, WT LPS 18 h vs UNT) and in DKO cells (B) (DKO LPS 2 h vs UNT, DKO LPS 6 h vs UNT, DKO LPS 18 h vs UNT).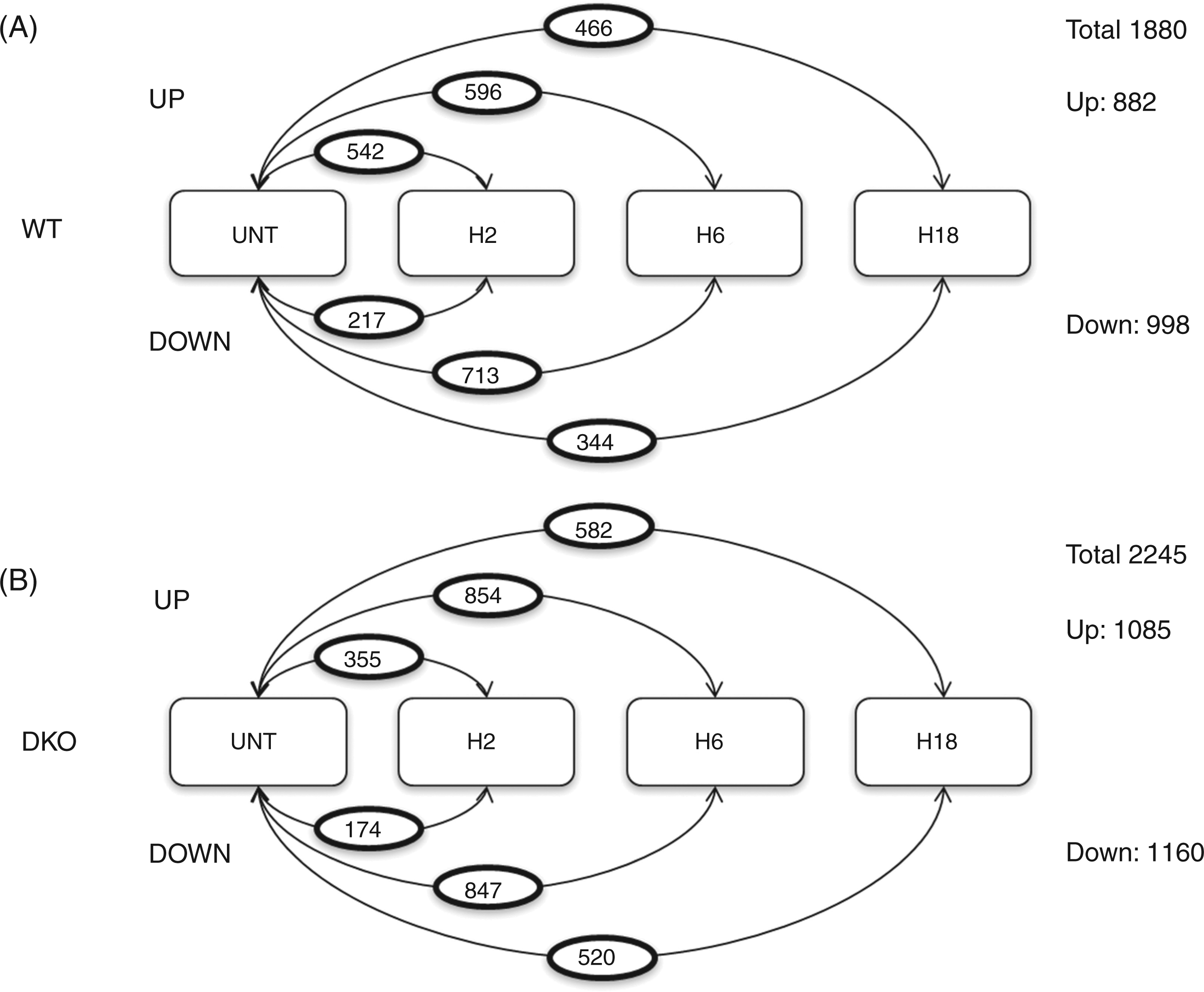
Functional clustering of genes modulated by LPS stimulation in WT and cRel−/−P50−/− peritoneal macrophages
Down-modulation of PRRs and anti-microbial proteins transcripts in DKO LPS-stimulated peritoneal macrophages.
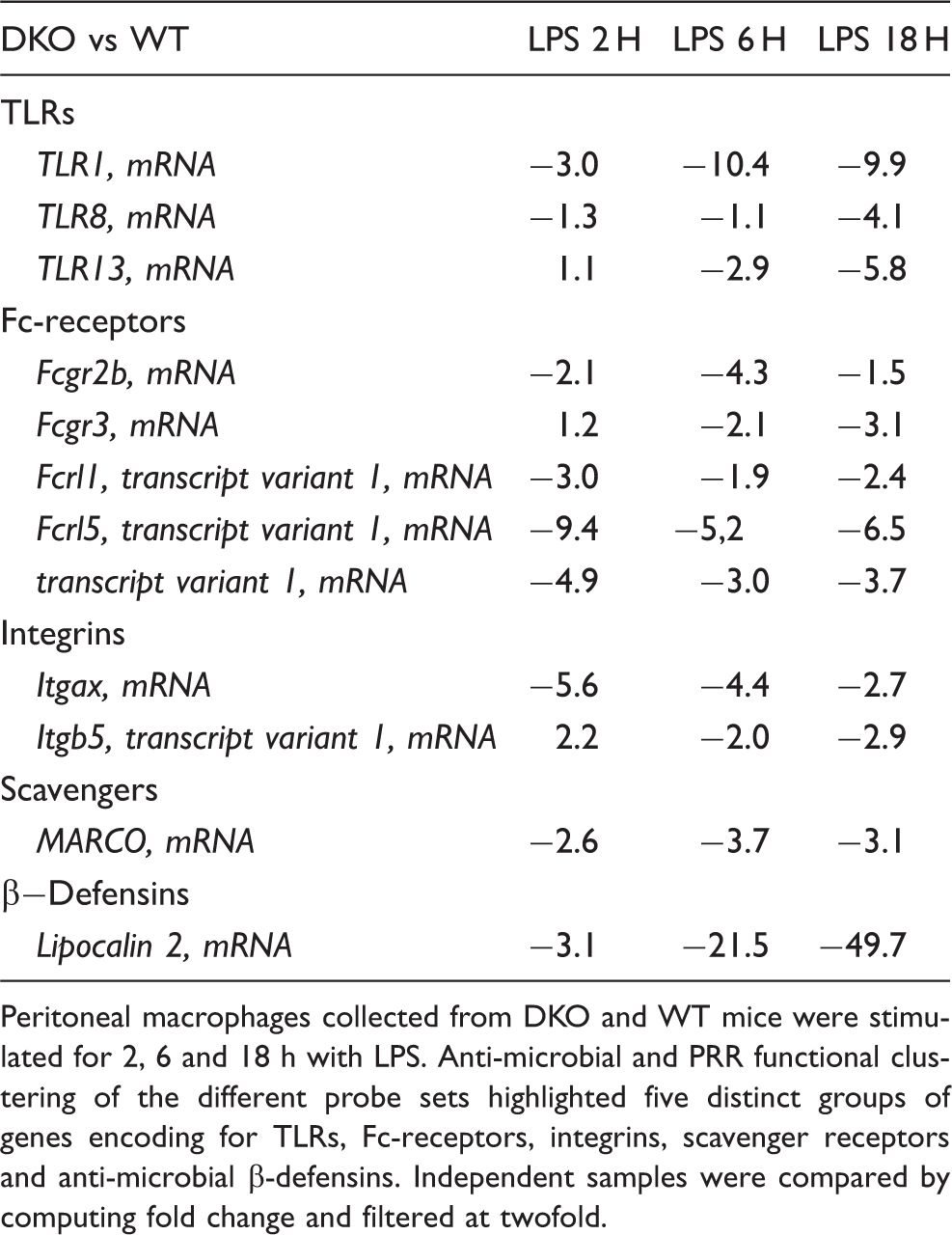
Peritoneal macrophages collected from DKO and WT mice were stimulated for 2, 6 and 18 h with LPS. Anti-microbial and PRR functional clustering of the different probe sets highlighted five distinct groups of genes encoding for TLRs, Fc-receptors, integrins, scavenger receptors and anti-microbial β-defensins. Independent samples were compared by computing fold change and filtered at twofold.
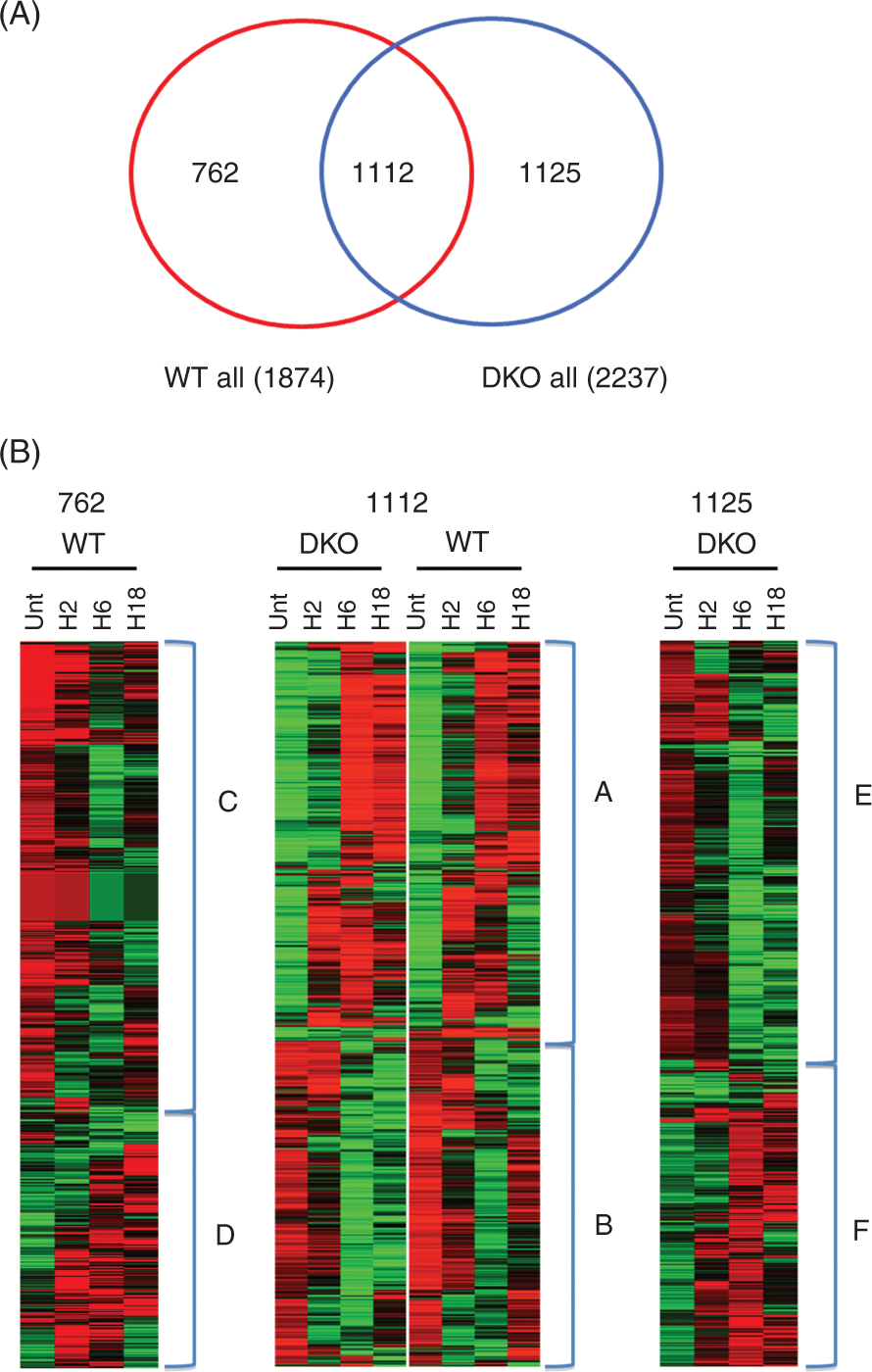
Heat map analysis and functional annotation of gene clusters differently modulated in DKO and WT LPS-stimulated peritoneal macrophages. Peritoneal macrophages collected from DKO and WT mice were stimulated for 2, 6 and 18 h with LPS. (A) Venn-diagram of probes expressed differentially in WT and in DKO peritoneal macrophages. Number in the overlapping region of the Venn diagram represents common regulated genes. Number of WT-specific genes is shown inside the right circle (red). Number of DKO-specific genes is shown inside the left circle (blue). (B) Heat map depicting the impact of cRel and p50 loss on the mRNA abundance of specific clusters. Heat map of the 2999 probe sets were classified in DKO-modified probe sets (n = 1125, left panel), common probe sets (n = 1112, middle panel), and WT-modified probe sets (n = 762, right panel). Probes are represented as rows and samples as columns. Probes were classified by similarity of expression profile among the samples. Colors indicate the normalized expression values below (green) and above (red) the row median expression.
Down-modulation of transcripts encoding pathogen recognition receptors and antimicrobial proteins in DKO peritoneal macrophages
As the detection of pathogens is the initial step in the first line of immune defense, the gene ontology analysis was focused on pathogen recognition receptors and antimicrobial molecules. This revealed that the combined lack of cRel and p50 led to the decreased LPS-induced transcription of several genes for PRRs, such as tlr1, the macrophage scavenger receptor MARCO and also genes encoding important antimicrobial products such as lcn2 (Table 1). These results were confirmed by RT-PCR (Figure 7A) and validated at the protein level by Western blotting for MARCO, a scavenger receptor directly implicated in the phagocytosis of non-opsonized E. coli by macrophages (Figure 7B).
25
Collectively, these results indicate that the cRel and p50 NF-κB subunits together control the gene expression of a subset of PRRs and antimicrobial molecules upon host infection.
Analysis of PRRs transcripts and antimicrobial proteins in DKO peritoneal macrophages. (A) Real-time PCR analysis of MARCO, TLR1 and LCN2 gene expression. RNA was isolated from untreated or LPS-stimulated WT and DKO peritoneal macrophages. RT-PCR for MARCO, TLR1 and LCN2 was performed in triplicate. Reported fold changes were normalized to HPRT in each sample. (B) Western Blot analysis of MARCO expression in DKO and WT macrophages. Protein extracts were made from untreated or 24 h LPS-stimulated WT and DKO BMDMs. Comparative expression of MARCO between WT and DKO cells was analyzed by Western blot using the specific Ab ED31 for MARCO. Protein loading was controlled by immunobloting with the anti-actin Ab.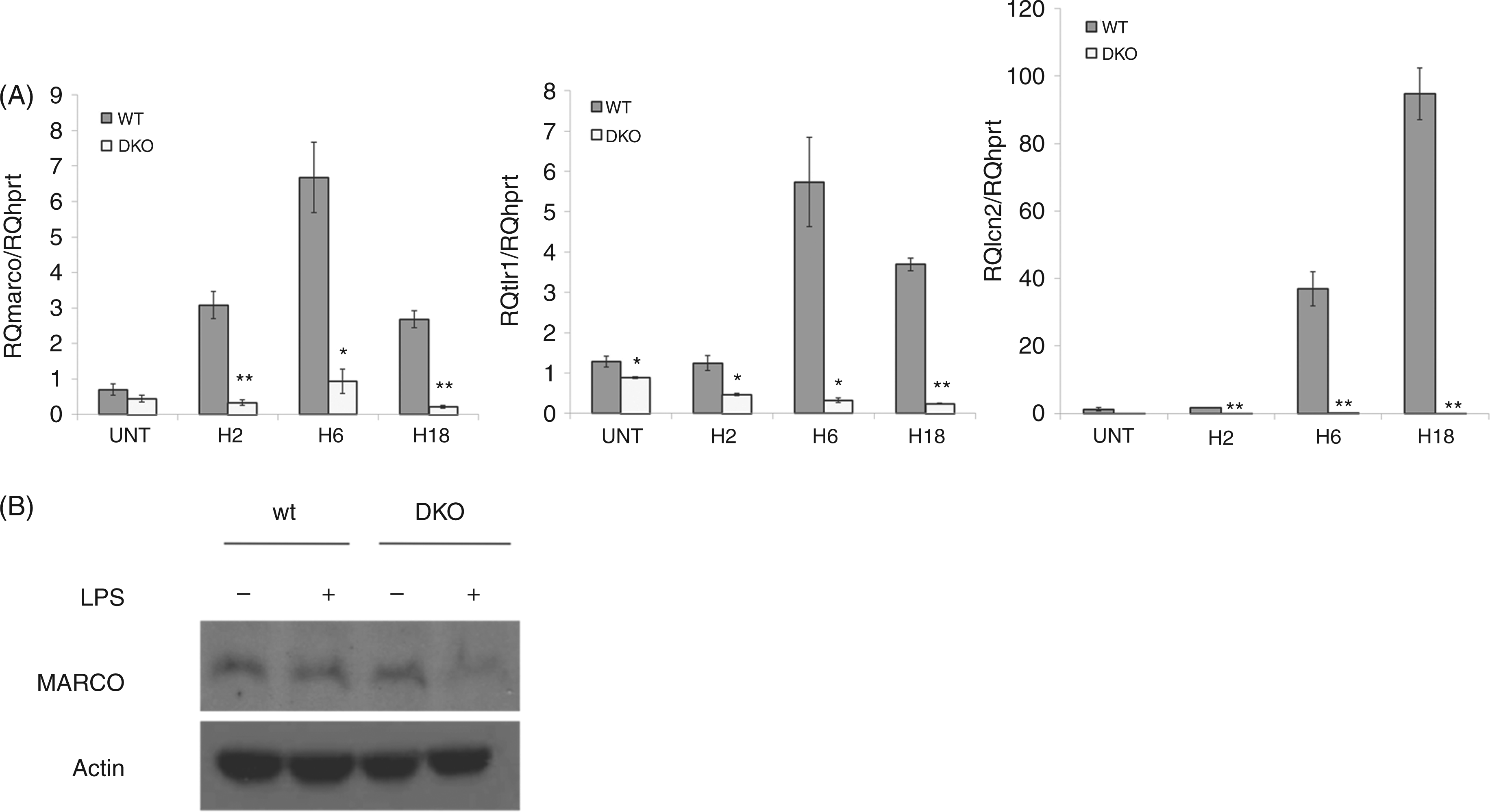
Discussion
In this study, we report that mice deficient for cRel and p50 subunits of NF-κB have increased mortality rate, impaired phagocytosis and decreased bacterial clearance following polymicrobial sepsis. Furthermore, gene profiling of LPS-stimulated peritoneal macrophages revealed dramatic differences between WT and cRel−/−p50−/− (DKO) cells that included diminished expression of PRRs and antimicrobial molecule transcripts, suggesting that the combined activity of the cRel and p50 subunits are essential for expression of innate immune proteins involved in pathogen clearance, supporting the in vivo mice findings in the CLP model of sepsis.
To highlight the combined role of cRel and p50 subunits in innate defense, the consequences of the DKO have been evaluated in vivo by analyzing the impact on survival and pathogen clearance in polymicrobial sepsis. Although the exact mechanism responsible for the elevated mortality of the DKO mice is not clear, reduced pathogen clearance observed at d 1, 2 and 3 in DKO mice is likely to contribute to this phenotype (Figure 2). Indeed, in animals, the control of microorganism dissemination is closely associated with mortality. 26 Similarly in humans, early initiation of adequate antimicrobial therapy and source control has been shown to improve the overall survival of septic shock. 27 Hence, bacterial dissemination results from either the influence of the bacterial load or the defects in leukocyte recruitment to the site of infection, pathogen capture, the killing process or in cytokine release.2,5 Despite the normal recruitment of peritoneal phagocytic Gr1+CD11b+ cells (data not shown), DKO mice exhibit severe defects in pathogen recognition and/or phagocytosis. Decreased expression of several PRRs in LPS-stimulated DKO peritoneal macrophages may provide a potential explanation of this phenomenon. One such example—the macrophage class A scavenger receptor MARCO—is known to be directly involved in phagocytosis of non-opsonized E. coli. 25 Interestingly, MARCO-deficient mice have been recently reported to be defective in the bacterial clearance of an acute systemic infection. 28 Furthermore, our microarray analysis also revealed diminished expression of lcn2, which encodes lipocalin-2, a β-defensin that limits bacterial growth by sequestering iron. This finding is also consistent with the increased susceptibility of lcn2−/− mice to bacterial sepsis.29,30
The family of NF-κB transcription factors consists of 12 potential combinatory dimers driving multiple distinct biological functions.7,8 The transcriptional and biological consequences arising from a double deficiency of cRel and p50 could result directly from the absence of those NF-κB dimers containing cRel and/or p50, or the replacement of these dimers by remaining dimers of the NF-κB family (RelA-RelA, RelA-p52, p52-p52 or RelB-p52).
Sepsis results in a potent inflammatory response that needs to be counterbalanced by a compensatory anti-inflammatory response. 31 As NF-κB is crucial for the regulation of both processes, we expected to observe dramatic differences in cytokine release in the absence of two important Rel subunits. Surprisingly, the DKO mice displayed an almost similar inflammatory response to WT mice. The functional genomics analysis confirmed that inflammation is preserved in DKO macrophages. The cytokine data are consistent both for gene expression in macrophages (Figure 6B, cluster A; Supplemental Table 1) and at the plasma level illustrated in mice (Figure 4). Hence, despite the differences in model (macrophage versus mice), stimulus (LPS versus CLP) and in the timing, the absence of cRel/p50 does not seem to affect dramatically the main cytokine response. It is well known that full and optimal cytokine gene expression requires the involvement of several transcription factors, including NF-κB. For NF-κB, the cytokine expression findings could be explained by both dimer absence and dimer exchange. 8 Indeed, the loss of these two subunits of NF-κB will lead directly to the absence of generation of homodimers, such as p50/p50 that is known to be a strong repressor for pro-inflammatory cytokines. In parallel, activating heterodimers such as RelA/p50 will also be absent, and the final picture might be equilibrated. At the same time, the absent dimers could be replaced by either activator dimers such as RelA/RelA or by p52/RelB which is a poor activating transcriptional factor. However, this is a very restrictive approach and the importance of other transcriptional factors in the context of less NF-κB activity is probably underestimated.
To highlight the potential mechanisms underlying the differential outcome between WT and the DKO mice in sepsis, genome-wide analysis was performed in LPS-stimulated macrophages. As expected, LPS-stimulation of WT macrophages led to dramatic changes in the expression of 5.4% of the whole genome (1583/28,944 genes), consistent with the genomic findings in whole blood after endotoxin injection in human volunteers. 32 In comparison, the absence of two subunits of NF-κB led to an overall increased number of LPS-induced gene expression changes. A functional global gene expression analysis revealed that the inflammatory response was preserved in DKO macrophages, being the most significant function in the common core response (P = 2.9 × 10−34, Supplemental Table 1), suggesting that absence of cRel and p50 did not affect expression of genes involved in one major aspect of the host response to LPS (such as TNF, IL-1 b, IL-6, nos2 that were confirmed by RT-PCR, data not shown).
The dimer exchange hypothesis might partly explain these results through RelA recruitment. Hence, the presence of RelA, which is known as a powerful activator for the transcription of many pro-inflammatory cytokines,7,12 may compensate the combined loss of cRel and p50 by promoting generation of RelA-containing NF-κB dimers. These results are consistent with a previous study that showed that in contrast to RelA, cRel and p50 subunits were not found to be crucial for pro-inflammatory gene expression but were required for T cell co-stimulatory molecule expression. 12
An analysis of these gene expression profiles also revealed a modulation of a large network of different clusters of genes controlling various biological processes (Supplemental Tables 1–3) that include the cell cycle (P = 7.01 × 10−11), immune cell trafficking (P = 5.65 × 10−10), cell death (P = 2.55 × 10−10), lipid metabolism (P = 3.81 × 10−8) or organismal survival (P = 7.89 × 10−7). Macrophages are key immune cells, critical for both bacterial clearance and the inflammatory response. Interestingly, the combined loss of both cRel and p50 led to down-modulated transcription of genes encoding for PRRs and antimicrobial molecules (Table 1). Two clusters of genes involved in ‘immune cell trafficking’ and ‘organism survival’ were specifically up-regulated in WT mice (Supplemental Table 2), suggesting that an efficient immune cell function is associated with the improved survival of WT animals to polymicrobial sepsis. 33 Among them, several genes necessary for Th17 cell functions are activated, including STAT3, IL-17, and IL-22. 34 In parallel, gene ontology highlighted two subgroups of genes involved in ‘cell death’ and ‘inflammatory response’, respectively, which are up-regulated in the DKO cells. These genes included genes that have been associated with mortality, such as FAS, caspase 1 or caspase 8 (Supplemental Table 3B). 35 These results are consistent with the association of immune cell apoptosis and sepsis mortality.33,35 Indeed, DKO mice exhibit dramatic survival defect of conventional and plasmacytoid dendritic cells21,22 and also of T lymphocytes, 23 which are key cells in mounting an adequate immune defense against infection.
There is a limitation in the transcriptome study. This analysis, which was initially designed as ‘an explorative approach’ to understand the potential mechanisms of mortality, was not realized with the highest standard of experiments. However, it is interesting to notice that for several genes, such as those for PRRs and cytokines, the data were verified by qPCR (Figure 7 and data not shown).
Surprisingly, the increased mortality of the DKO mice does not seem to be associated with a dramatic alteration in cytokine release in the plasma as these animals displayed similar inflammatory systemic responses to those of WT mice, despite defective pathogen clearance. Recently, we have reported that cRel−/− mice exhibited an increased mortality in the CLP model. 19 Interestingly, despite a normal bacterial clearance these single KO mice show an enhanced inflammatory systemic response, confirming that sepsis-induced death may result from complex and distinct mechanisms in both animal models.
Previous studies have shown that cRel and p50 subunits promote cell survival, IL-12 production and co-stimulatory molecule expression in dendritic cells—all crucial for the development of an efficient adaptive immunity.12,22 Here, we report for the first time that these two subunits also play a role in innate defense by controlling both bacterial phagocytosis and pathogen clearance by macrophages. Taken together, these findings suggest that cRel and p50 subunits of NF-κB exhibit a dual function in distinct antigen-presenting cells (APCs) and in consequence regulate both innate and adaptive immune responses during infection.
In conclusion, the present study highlights the transcriptional roles of cRel and p50 in PRR expression and the combined loss of these two subunits led to impaired bacterial clearance and decreased survival in polymicrobial sepsis. Moreover, we demonstrated that the lack of these two subunits of NF-κB does not significantly impact the inflammatory response. These findings should be integrated in the development of NF-κB-targeted immunomodulation strategies in severe infections.
Footnotes
Funding
Cochin Association for Research on Inflammation Sepsis and Molecular Advances (CARISMA) supported this work. The funders had no role in study design, data collection and analysis, decision to publish or the preparation of the article. JM was supported by a doctoral fellowship from ANRS.
Acknowledgements
We would like to thank members of the transcriptomic (Franck Letourneur), flow cytometry (Brigitte Chanaud, Laurence Stouvenel), and animal facilities (Véronique Fauveau), at the Cochin Institute for their helpful assistance.
Conflicts of interest
The authors had no financial conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.