
Abstract
Background. Aging is associated with declining mobility, which negatively affects quality of life and incurs substantial economic costs. Techniques to maintain safe mobility in older adults are therefore essential. Rhythmic auditory cueing (RAC) can improve walking patterns in older adults. However, the neural correlates associated with RAC, how they are influenced by repeated exposure and their relationships with gait response, cognitive function, and depressive symptoms are unclear. Objectives. This study aimed to investigate the effects of RAC during walking on cortical activation and the relationship between RAC-related cortical changes and cognitive function, depressive symptoms, and gait response. Methods. Seventeen young adults and eighteen older adults walked on a motorized treadmill for 5 minutes (5 trials with alternating 30-second blocks of usual walking and RAC walking). Changes in oxygenated hemoglobin (HbO2) in the frontal cortex were recorded using functional near-infrared spectroscopy. Cognitive domains were assessed through validated tests. A triaxial accelerometer measured gait parameters. Results. Gait variability decreased and prefrontal HbO2 levels increased during cued walking relative to usual walking. Older adults showed greater HbO2 levels in multiple motor regions during cued walking although the response reduced with repeated exposure. In older adults, lower depression scores, higher cognitive functioning, and reduced gait variability were linked with increased HbO2 levels during RAC walking. Conclusion. These findings suggest that walking improves with RAC in older adults and is achieved through increased activity in multiple cortical areas. The cortical response decline with repeated exposure indicates older adults’ ability to adapt to a new task.
Introduction
Walking deteriorates with age and this is associated with structural and functional changes in the brain and decreased independence and safety.1-3 Age-related gait impairments include reduced walking speed, step length, and swing time, and greater intraindividual gait variability.4,5 Impaired gait is a strong and independent risk factor for falling 6 with fall-related complications leading to diminished quality of life 7 and considerable economic costs. 8 With aging, there is also a substantial increase in the prevalence of multi-morbidity which is associated with greater levels of immobility. 9 Structural and functional changes in the brain occur with aging. 2 together with cognitive impairments affecting particularly memory, attention, planning, and processing speed. 10 Reduced mobility is linked to subclinical changes in cognition and brain structures.2,3 Functionally, age-related changes during imagined locomotion and rhythmic movements include increased activity in, or activation of, additional brain regions,11,12 which may represent a compensatory mechanism due to impaired brain function and/or loss of automatism. Effective therapies to maintain safe mobility in aging are therefore essential to help overcome age related changes in neural pathways controlling gait.
The application of rhythmic auditory cueing (RAC), a relatively simple, low-cost approach, has been used effectively to improve gait in patient populations such as those with Parkinson’s disease.13,14 The limited research on cueing in healthy older adults demonstrates a beneficial effect on gait spatiotemporal measures, including, but not limited to, increase in step length and speed and decrease in temporal variability.15-17 Underlying neural correlates identified from neuroimaging studies investigating upper or lower limb synchronization to a rhythmical cue include motor cortical regions, such as the premotor cortex (PMC) and supplementary motor area (SMA).18-20 There is also evidence that rhythmic stimulation facilitates basal ganglia–PMC interactions18,19 in addition to cerebello-thalamo-cortical pathways. 21 The neural correlates of real cued walking are however not well defined. One study on healthy older adults indicated that stepping in time to rhythmic auditory cues relies on executive/attentional control, implying increased prefrontal cortex (PFC) involvement. 15
Factors affecting response to cueing are not well described in the literature. Clinical observations suggest that not every individual responds positively to cueing, which may occur due to the inability to recruit appropriate brain regions to control cued walking. For example, the effect of cognitive function on responsiveness to cued walking is unknown, with studies showing conflicting gait responses in people with dementia exposed to a cueing stimulus.14,22,23 Additionally, diminished attention is frequently present in older adults, which may result in a decreased response to an auditory cue. Depressive symptoms may also play a role in response to cueing as they limit the capacity to recruit cortical areas associated with cognitive processes. 24 Another important consideration when applying cues to improve mobility is how repeated exposure (or habituation) may affect one’s response to cueing. Although initial exposures to RAC suggest increased attentional control and movement specific reinvestment, 15 repeated exposure may result in adaption and lead to a more automatic control of gait, with fewer demands on cognitive and attentional resources. Recording cortical activity during repeated exposures to RAC is essential to understand this issue better and it has not yet been reported in the literature. The development of a robust body of knowledge about factors affecting the neural response to cueing will contribute to the optimization of interventions.
In this study, we apply RAC during repeated trials of walking to investigate the effect of aging on the response of frontal cortical regions to auditory stimuli. We aim to identify how RAC-related changes in cortical activity relate to subsequent gait response to RAC. Additionally, we explore the relationship between RAC-related changes in cortical activity and measures of cognitive function and depressive symptoms. We hypothesized first, that cued walking will result in more widespread cortical activity in older adults and decreased gait variability. Second, stepping in time to a rhythmical beat will be more attentionally demanding, indicated by increased PFC activity. Our third hypothesis was that repeated exposure to RAC will lead to a more automatic control of gait (expressed as reduction in cortical activity over trials). Finally, our fourth hypothesis was that depressive symptoms and reduced cognitive function will be associated with decreased cortical response to cued walking.
Methods
Participants and Initial Screening
Power analysis using preliminary pilot data from healthy younger and older participants indicated that a sample size of 30 (15 per group) was sufficient to identify changes of 4.3% with a power of 0.8. Fifteen healthy young and 15 older adults therefore participated in this study. Participants were recruited through local advertisements (ie, posters placed in strategic areas of the campus; e-mails circulated to staff and students; a call for older adult participants was made through community-based partnerships). Study procedures were approved by the university ethics committee and participants provided written informed consent prior to participation. Participants were included if they were: able to walk unaided for at least 5 minutes; community dwelling; had adequate hearing and vision (as judged by correct recall on the “whisper test” and a score of 6/12 using the Snellen chart, respectively); and within the age range of 20 to 40 years for young adults and ⩾60 years for older adults. Exclusion criteria included: diagnosis of major gait abnormality; major depressive disorder; clinical diagnosis of dementia or other severe cognitive impairment (defined as Montreal Cognitive Assessment, MoCA; score <21) 25 ; chronic musculoskeletal, unstable cardiovascular, neurological or respiratory disease; and acute lower back or lower extremity pain.
Demographic and Cognitive Assessments
The following demographic information was collected: age, height, weight, and years of education. Mood and several cognitive domains were assessed through validated questionnaires. Global cognitive function was measured with the MoCA (score range 0-30; higher score indicates greater cognitive function). 25 Depressive symptoms were assessed using the Beck Depression Inventory (range 0-63; higher score indicates greater depressive severity). 26 Executive function and visuospatial ability were assessed using clock drawing and copying tasks (Royall’s CLOX 1 and 2). 27 Working memory was assessed using forward digit span from the Wechsler Adult Intelligence Scale. 28 Fear of falling was measured using the Falls Efficacy Scale–International (FES-I), 29 which is a multiple-choice self-report questionnaire.
Experimental Design and Equipment
All participants completed a warm up walk permitting familiarization with walking on a treadmill and preferred walking speed was recorded. No participants exhibited any difficulty in walking on the treadmill during this initial walk. Participants walked on a treadmill, at self-selected preferred walking speed, performing 2 different tasks: (a) usual walking and (b) cued walking (RACs were delivered with a digital metronome at preferred stepping frequency). Preferred walking speed on the treadmill was determined by increasing belt speed until it was faster than the participants’ preferred speed, then reducing belt speed until preferred speed was achieved. Participants were asked to walk at selected speed for 1 to 2 minutes so that preferred stepping frequency could be calculated (steps per minute). The 2 tasks were presented in a 5-minute block of alternating tasks (block design). The block consisted of 30 seconds of usual walking followed by 30 seconds of cued walking with 5 trials/repetitions. Participants were instructed to step in time to the beats for the cued walking condition. Preferred cadence has been reported as optimal coupling between gait and metronome beats. 30
A tethered functional near-infrared spectroscopy (fNIRS) system (LABNIRS, Shimadzu, Kyoto, Japan) with continuous wave laser diodes with wavelengths of 780, 805, and 830 nm was used to record changes in oxygenated (HbO2) and deoxygenated (HHb) hemoglobin at a sampling rate of 23.8 Hz. Tethered fNIRS systems commercialized by Shimadzu (eg, LABNIRS, OMM-3000, OMM-2001, OM100-A) have been used in different areas of research (including but not limited to sports medicine, rehabilitation, and cognitive psychology). Noah et al 31 investigated the validity of the Shimadzu OMM-3000 system, and reported that in a dance video game, the correlation between fNIRS and functional magnetic resonance imaging (fMRI) signals was high (r = 0.78; P = .03). Miyai et al 32 assessed the cortical activation during foot movements and gait imagery using both fNIRS (by Shimadzu) and fMRI. The activation patterns of supplementary motor areas and medial primary motor cortices in fMRI were similar to those of fNIRS topography. Dravida et al, 33 using the LABNIRS system, observed high test-retest intraclass correlation (ICC) for HbO2 during manual tasks (overall ICC = 0.76).
A 40-channel arrangement with 25 optodes (5 × 5), consisting of 13 transmitters and 12 detectors, covered the frontal lobe in both right and left hemispheres. Interoptode distance was 30 mm. After the Cz position was determined on participants’ head, a whole-head fiber holder marked with labels of the international 10-10 electroencephalography system (Whole-Head Fiber Holder, Shimadzu, Kyoto, Japan) was placed on the participant’s head. A digitizer (FASTRAK, Polhemus, VT, USA) was used to provide 3-dimensional coordinates of anatomical references (Cz, nasion, and left and right preauricular points) and positions of optodes. The Spatial Registration routine (stand-alone NIRS, using 3-dimensional digitizer) available in the free software package NIRS-SPM (NIRS-SPM, http://www.nitrc.org/projects/nirs_spm) 34 was used to find the correspondence between the scalp location where the fNIRS measurement was performed and its underlying cortical surface where the source signal was located. NIRS-SPM allows registration of fNIRS channel data onto the Montreal Neurological Institute standard space. 35 Cortical regions assessed included PFC (Brodmann areas 8, 9, 10, 45, and 46), PMC (Brodmann area 6, lateral), SMA (Brodmann area 6, medial), and primary motor cortex (M1; Brodmann area 4). Participants wore a triaxial accelerometer-based device (AX3; Axivity, York, UK; dimensions: 23.0 mm × 32.5 mm × 7.6 mm; weight 9 g) at the fifth lumbar vertebral level, which recorded gait data. The accelerometer was attached using double sided tape and Hypafix (BSN Medical Limited, Hull, UK) and programmed to capture with a sampling frequency of 100 Hz (16-bit resolution, range ±8 g).
Data Processing and Analysis
Processing of fNIRS signals followed current recommendations when possible. 36 The fNIRS data were preprocessed using the Time Series Analysis routine available in the NIRS-SPM and included the following steps: (a) filtering—high-frequency noise was reduced using a low-pass filter based on canonical hemodynamic response function 37 ; (b) detrending—a wavelet-minimum description length detrending algorithm was applied to decompose NIRS measurements into global trends, hemodynamic signals and uncorrelated noise components as distinct scales. 38 This reduced unknown global trend due to breathing, cardiac, vasomotion, or other experimental errors (including movement artifacts); (c) baseline correction—set at the initial time. We used HbO2 concentration as a marker for the regional cortical activation, since it is more sensitive to locomotion-related activities than HHb. 39 Preprocessed concentrations of HbO2 were exported to MATLAB (MATLAB and Statistics Toolbox Release R2015a, The MathWorks, Inc, Natick, MA, United States). First, channels were averaged per region of interest (ie, right and left PFC, PMC, SMA, and M1 and normalized (normHbO2) by dividing them by the corresponding signal amplitude of the whole experiment. 40 Each of the 5 trials was divided into 2 phases: (a) a control task period running from 15 to 25 seconds of usual walking and (b) an experimental task period running from 5 to 25 seconds after the start of cued walking. The 10 seconds of usual walking period prior to the task served as the baseline. The last 5 seconds of the usual walking and initial and final 5 seconds of RAC walking were removed to exclude anticipatory changes by the participant (due to the constant 30-second × 30-second design). Additionally, removal of these transitional periods accounted for the 4- to 6-second delay of the hemodynamic response. The normalized HbO2 concentration was averaged over the control task period (usual walking) and experimental task period (cued walking) for each region of interest and each trial. Differences between control task and the experimental task were also calculated for each region of interest and each trial to evaluate the relative change in HbO2 concentration (diffHbO2) during cued walking.
Spatiotemporal gait parameters were extracted from the accelerometer data according to published methods developed by our research group and described in previous publications.41,42 Gait variables included step time, swing time, step length, and gait variability (ie, step time, stance time, swing time, step length, and step velocity).
Statistical Analysis
Differences in the levels of normHbO2 between groups (ie, young vs older adults), tasks (ie, usual walking vs RAC) and trials (ie, 1, 2, 3, 4, and 5) were analyzed using linear mixed models in SPSS (v21, IBM, Armonk, NY, USA), P < .05, while controlling for treadmill speed. Bonferroni corrections for multiple comparisons (P value / number of comparisons) were applied to the post hoc analyses. Similar procedures were used to assess changes across trials in the diffHbO2 levels (ie, RAC HbO2 − usual walking HbO2), including group and trial as fixed factors. Linear regression analyses were performed to determine the individual slope of cortical activation (diffHbO2) across trials; independent samples t-tests were used to compare groups and 1-sample t tests were used to assess the attenuation or increase in cortical activation across trials. Gait measures were also analyzed using the linear mixed model approach. The association between cortical activation and cognitive or gait measures was explored using Spearman and Pearson correlation coefficients (according to data type and distribution).
Results
Participant characteristics are presented in Table 1. Older adults walked with a slower self-selected treadmill speed than young adults (P < .05). In addition, older adults showed decreased executive visuospatial ability (Royals CLOX 2 score) and less years of formal education than young adults (P < .05).
Demographic and Cognitive Characteristics.
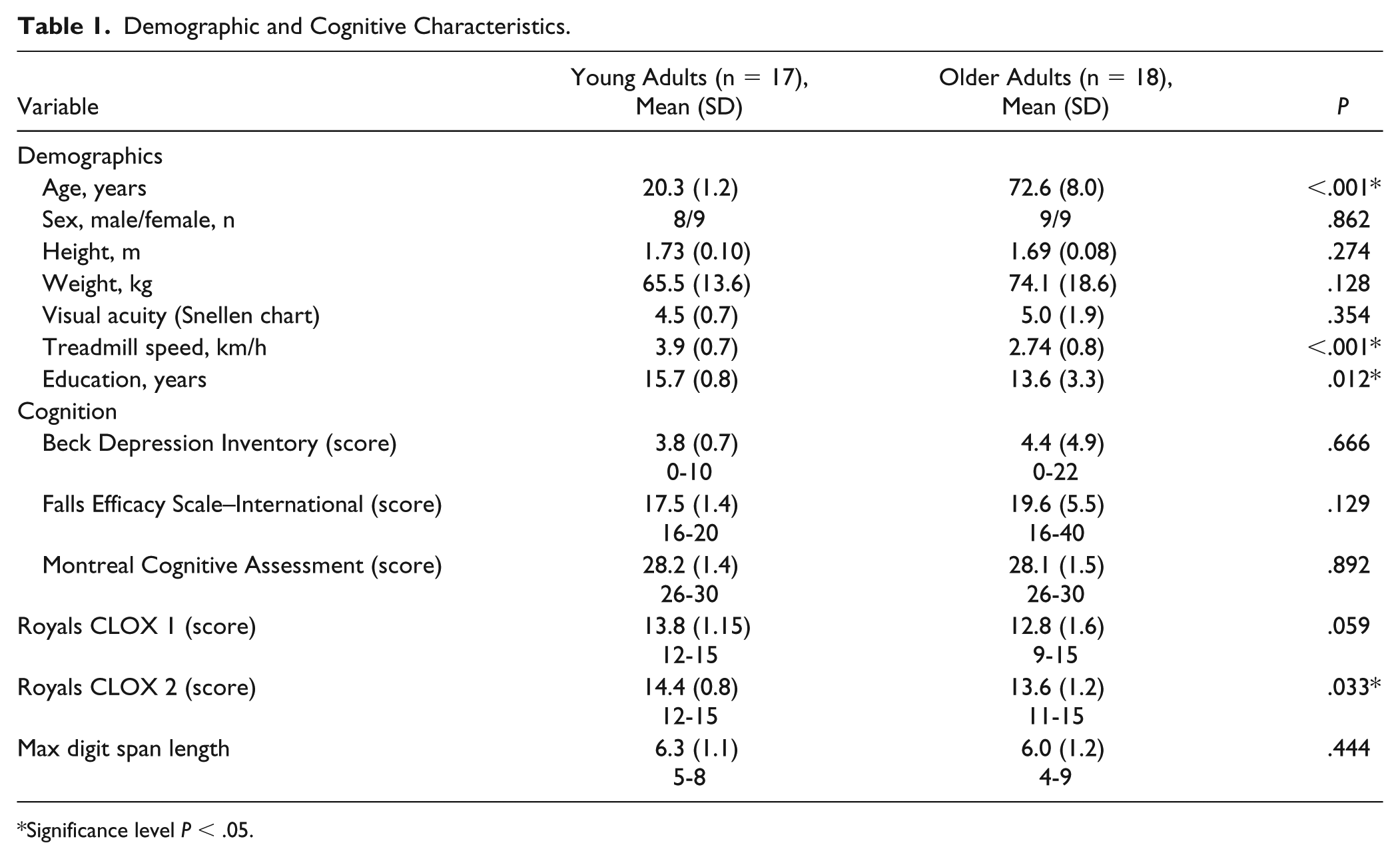
Significance level P < .05.
HbO2 Levels
Response to Rhythmic Auditory Cues Across Trials
Significant 2-way interactions (group*task) were observed for normHbO2 levels in the left PMC (F1, 297 = 9.527, P = .002), right PMC (F1, 297 = 16.965, P < .001), and left SMA (F1, 297 = 37.267, P < .001). Post hoc comparison tests revealed that older adults increased normHbO2 levels in the 3 motor regions (left PMC, effect size = 0.18; right PMC, effect size = 0.24; left SMA, effect size = 0.35) whereas younger adults did not increase motor cortical activity in response to RACs (Figure 1). Both groups increased bilateral PFC normHbO2 levels in response to RACs (left PFC-young: F1, 297 = 21.297, P < .001, effect size = 0.27; right PFC-young: F1, 297 = 19.531, P < .001, effect size = 0.26; left PFC-older: F1, 297 = 10.702, P = .001, effect size = 0.19; right PFC-older: F1, 297 = 11.029, P = .001, effect size = 0.19) (Figure 1).
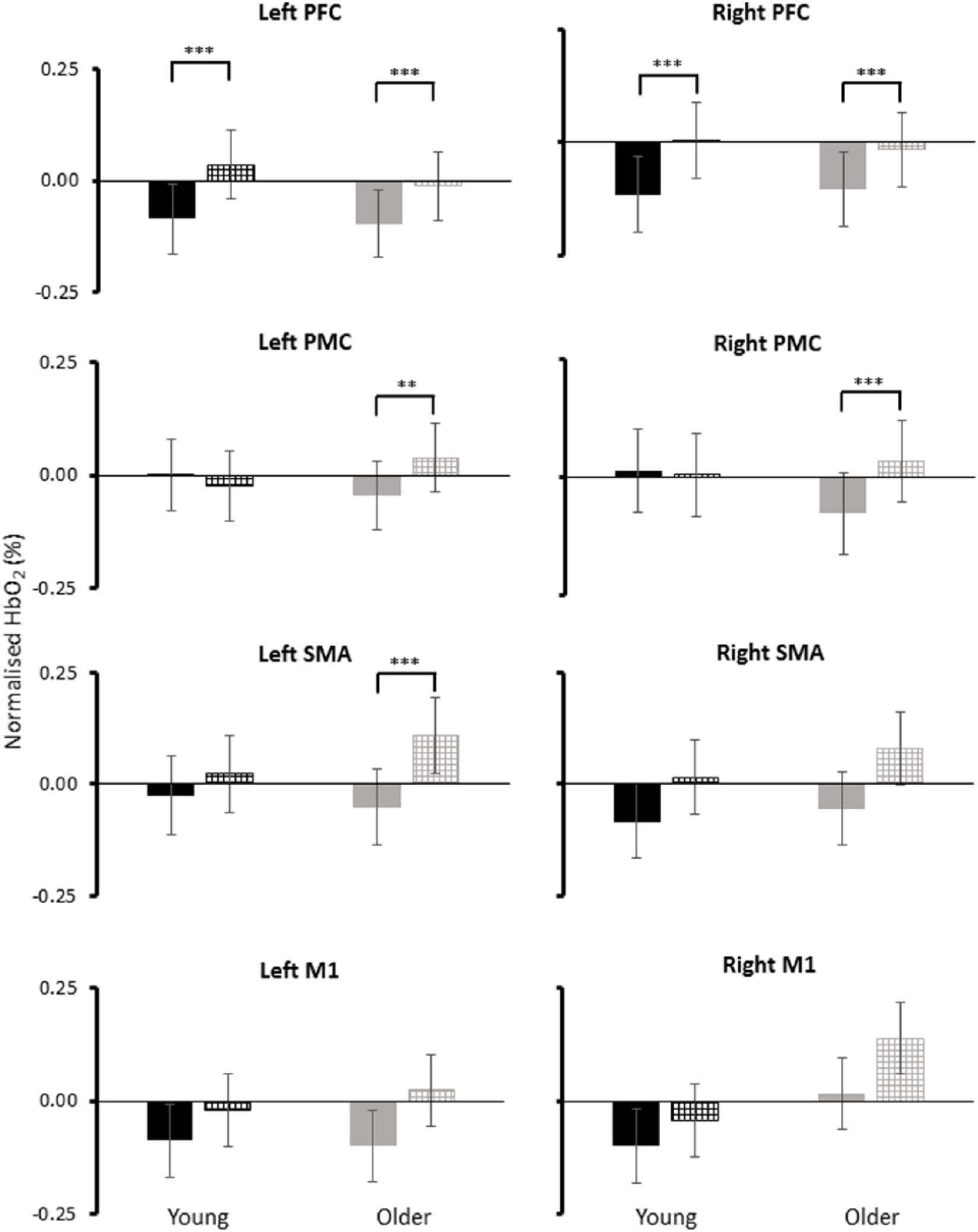
Normalized oxygenated hemoglobin (HbO2) levels for usual walking (solid black, young adults; solid gray, older adults) and walking with a rhythmic auditory cue (checkered black, young adults; checkered gray, older adults). PFC, prefrontal cortex; PMC, premotor cortex; SMA, supplementary motor area; M1, primary motor cortex.
Response to Rhythmic Auditory Cues Between Trials
Significant 3-way interactions (group * task * trial) were observed for normHbO2 levels in the SMA, PMC, and M1 (Table 2). Post hoc analysis revealed that in young adults, normHbO2 levels did not change during RAC, as compared with usual walking, in any of the 5 trials performed. In contrast, older adults increased normHbO2 levels during RAC, as compared with usual walking, in the first one or two trials performed (Table 2). Accordingly, older adults showed higher levels of diffHbO2 than young adults in the first and/or second trial (Figure 2). Further analysis revealed a trial effect for the motor regions with significant differences between trials for older adults only. The first and/or second trial displayed greater diffHbO2 levels compared with the fourth and/or the fifth trial (Figure 2).
Results for 3-Way Interactions (Group * Task * Trial) for Normalized Oxygenated Hemoglobin (HbO2) and Post Hoc Test Results for Older Adults for Usual Walking and Cued Walking.
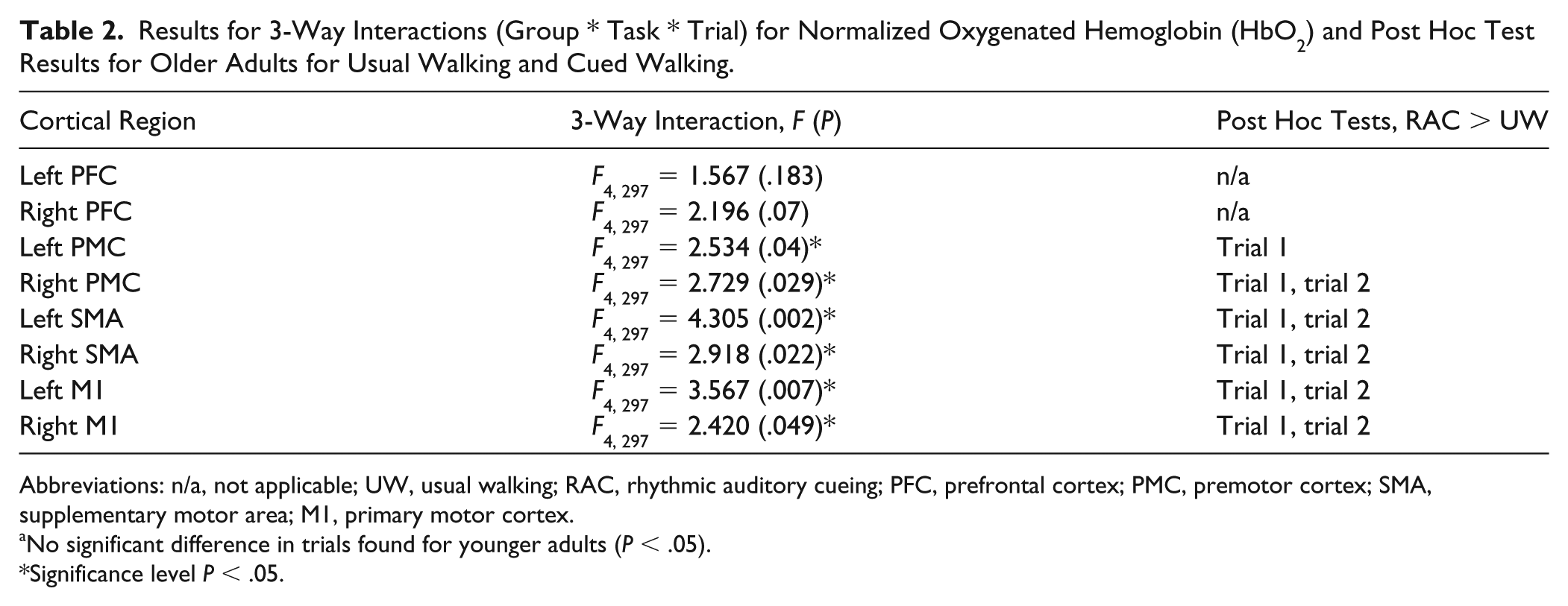
Abbreviations: n/a, not applicable; UW, usual walking; RAC, rhythmic auditory cueing; PFC, prefrontal cortex; PMC, premotor cortex; SMA, supplementary motor area; M1, primary motor cortex.
No significant difference in trials found for younger adults (P < .05).
Significance level P < .05.
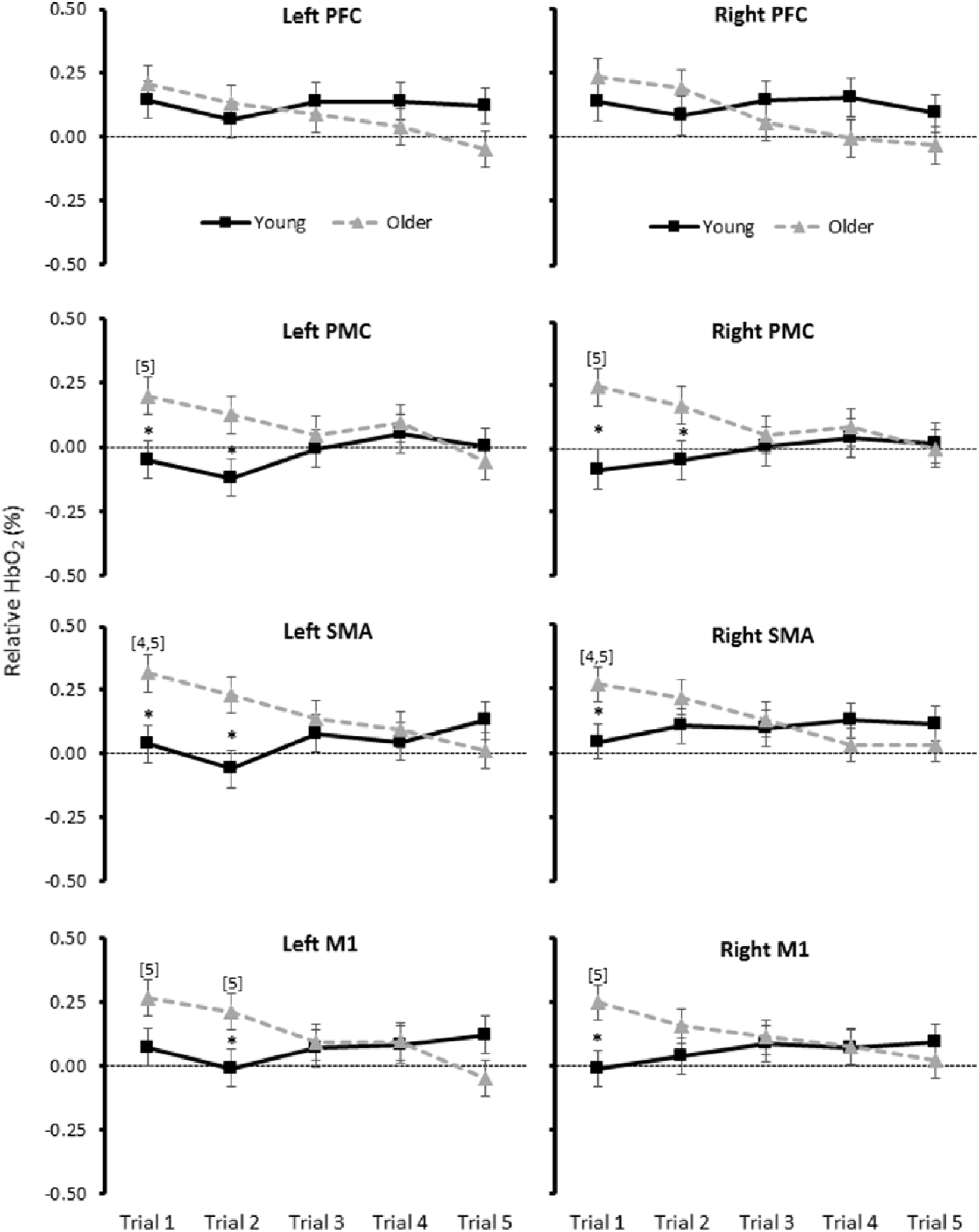
Relative oxygenated hemoglobin (HbO2; mean ± SE) concentrations for each group and trial. *Significant between group difference in the specified trial. [4] Significant difference from trial 4 for older adults only. [5] Significant difference from trial 5 for older adults only. PFC, prefrontal cortex; PMC, premotor cortex; SMA, supplementary motor area; M1, primary motor cortex.
Change in diffHbO2 Over 5 Trials
Linear regression analyses showed that cortical activation in older adults significantly attenuated across the 5 trials in all areas assessed. Young adults, by contrast showed no significant change over time (Supplementary Table S1).
Gait Measures
As expected, there was a significant difference between groups with older adults having a significantly shorter step length and increased gait variability compared to young adults (see Supplementary Table S2). The preferred cadence was 99.6 steps per minute for young adults and 97.5 steps per minute for older adults. RAC resulted in a significant increase in swing time and decreased step length variability and step velocity variability in both older and young adults, averaged across trials (see Supplementary Table S2). No significant interactions were observed for gait measures (P > .05). Linear regression analyses for gait confirmed that response to cueing did not change over time in both groups (P > .05).
Association Between RAC-Related Changes in Cortical Activity and Cognition, Depressive Symptoms, and Gait
In older adults, there was a significant negative association between the Beck Depression Inventory score and diffHbO2 in the bilateral PFC, right PMC, and right M1. Additionally, older adults demonstrated a positive association between the MoCA score and diffHbO2 in the right PMC (ρ = 0.547, P = .019) and left SMA (ρ = 0.472, P = .048) (Table 3). In contrast, there was no significant association between cognitive measures and diffHbO2 in young adults.
Correlations Between Relative Oxygenated Hemoglobin (HbO2) and Cognitive and Gait Measures for Older Adults.
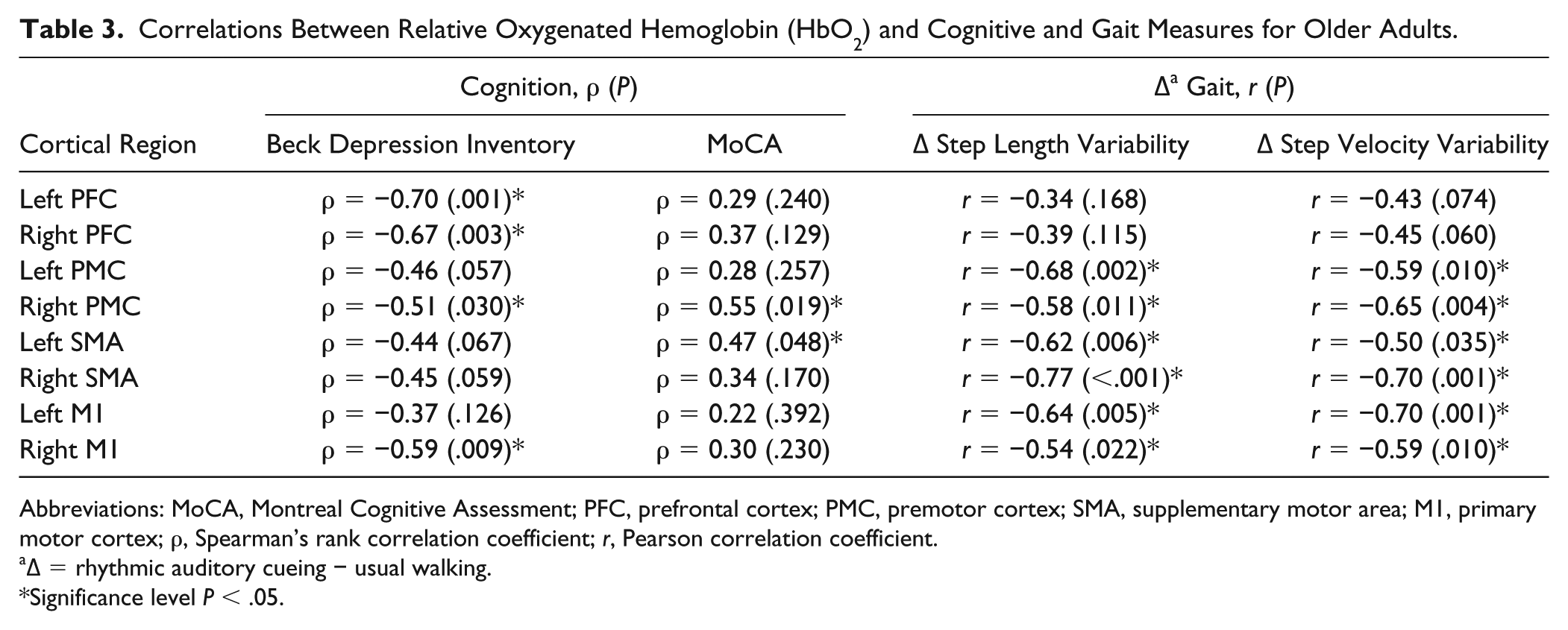
Abbreviations: MoCA, Montreal Cognitive Assessment; PFC, prefrontal cortex; PMC, premotor cortex; SMA, supplementary motor area; M1, primary motor cortex; ρ, Spearman’s rank correlation coefficient; r, Pearson correlation coefficient.
∆ = rhythmic auditory cueing − usual walking.
Significance level P < .05.
The diffHbO2 was also related to changes in gait in both young and older adults; however, there was a different pattern depending on group. In young adults, greater step length variability was associated with greater diffHbO2 in the left PMC and M1 (Figure 3). In contrast, in older adults, greater step length and step velocity variability were associated with diminished diffHbO2 in all motor regions (Figure 3; Table 3).
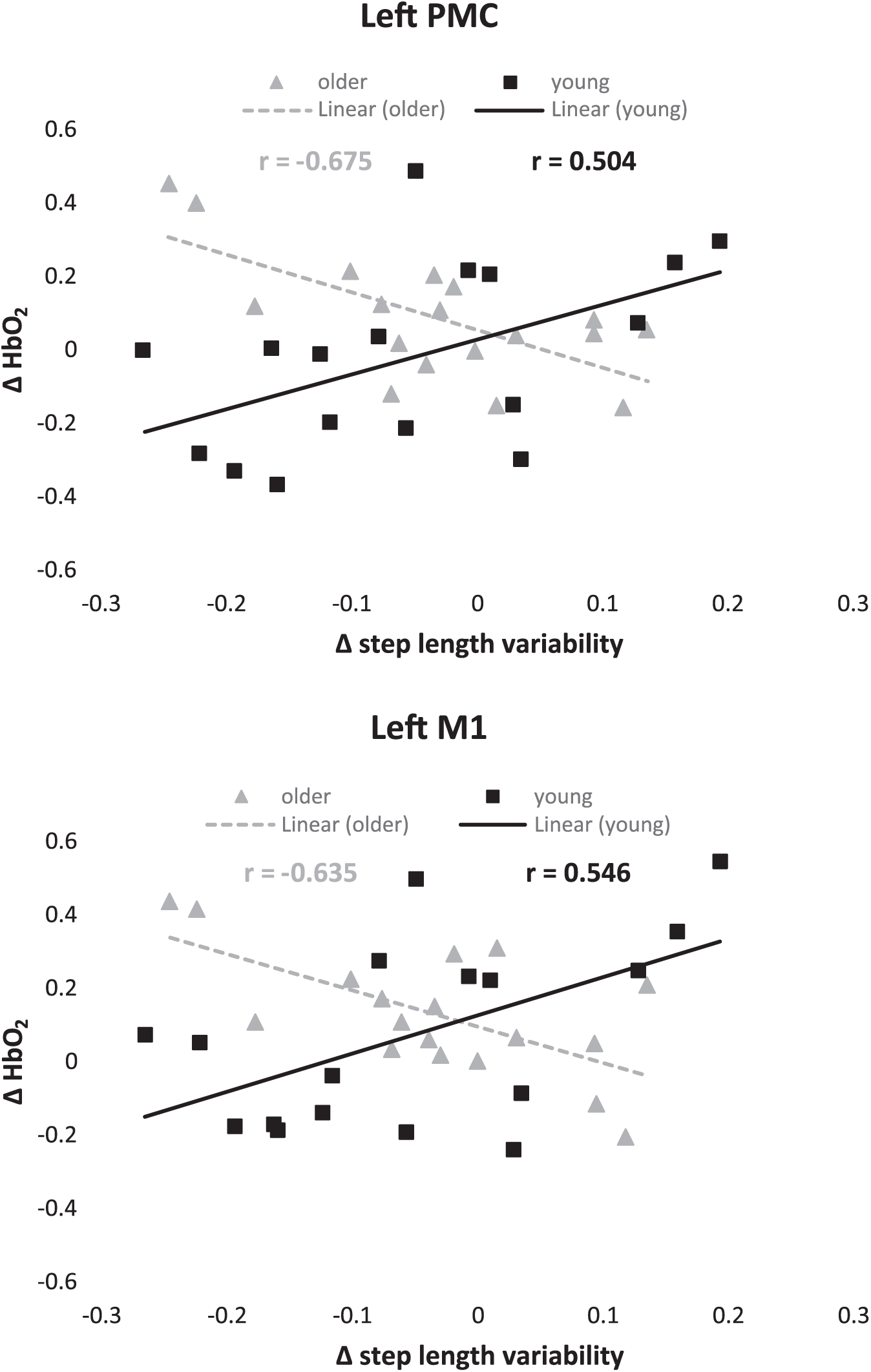
Correlations between relative oxygenated hemoglobin (HbO2) and changes in gait measures (Δ = rhythmic auditory cueing − usual walking). PMC, premotor cortex; M1, primary motor cortex.
Discussion
This study investigated the effects of aging on cortical activation in response to repeated trials of RAC during walking and explored the relationship between cortical changes and measures of cognitive function, depressive symptoms, and gait response. We demonstrated that aging leads to activation of additional motor regions, confirming our first hypothesis. Here we show for the first time that RAC walking increases PFC activity in both young and older adults, supporting our second hypothesis. The increased cortical activity measured in the first trial declined with repeated exposure to cueing in older adults, indicating an adaption process in healthy older adults; this finding confirmed our third hypothesis. Finally, corroborating our fourth hypothesis, we found significant correlations in older adults between RAC-related changes in cortical activity and measures of gait (response to RAC), cognitive function and depressive symptoms, suggesting strong interactions between motor and cognitive networks.
Differences in Motor Cortical Activity in Response to Cued Walking: The Effects of Age
Aging leads to activation of more motor cortical regions (PMC, SMA, and M1) when walking with RACs (Figures 1 and 2; Table 2), particularly during initial trials. Our findings are consistent with neuroimaging studies demonstrating older adults recruit additional motor areas during imagined walking.11,12 The neural compensation hypothesis 43 and the cognitive resource theories of aging 44 propose that as a given task becomes more demanding, older adults recruit additional cortical regions to manage the increased relative demands. One explanation for this theory is that age-related structural and/or functional changes in the brain impairs previously used brain networks. Consequently, the older adults recruited additional cortical regions (ie, PMC, SMA, and M1) while walking to RACs, demonstrating compensatory recruitment 45 . Increased cortical recruitment may also be explained by reduced cortical reciprocal inhibition reported in older adults. 46 The changes in cortical activity cannot be accounted for by differences in walking velocity, as this remained similar between walking tasks, in contrast to findings from a previous study. 11 Increased activity of the left SMA may be related to the lateralization that has been reported. The SMA is directly connected to Broca’s area in the left hemisphere in right sided dominant individuals through the frontal aslant tract. 47 The frontal aslant tract forms part of the posterodorsal auditory pathway involved in sensorimotor integration and may play an important role in cued motor responses.
Cortical Activation Patterns in the PFC in Response to Rhythmic Auditory Cueing of Gait
RAC intensifies the usage of PFC cognitive resources (ie, attention and executive functions) during walking in both young and older adults. This concurs with one study demonstrating the attentional and/or executive demands of walking to rhythmic auditory cues in healthy older adults. 15 The process of synchronizing each step to a beat increases executive and/or attentional control as supported by greater PFC activity irrespective of age.
Effect on Cortical Activity of Repeated Exposure to Rhythmic Auditory Cue
After two trials, the increased activation of cortical motor regions was no longer observed. Cued treadmill walking represents a novel task requiring greater precision and motor control and may present a higher task demand for older adults. The decreased motor cortical response with repeated exposure suggests incorporation of external temporal information signaling stepping frequency (provided by the RAC), into subcortical locomotor pathways, reducing task demand. Linear regression analysis of cortical activation showed older adults also decreased PFC activation with repeated exposure. A possible explanation is that older adults may have a limited capacity to concentrate on RACs over prolonged periods. Alternatively, repetitions of the task may result in adaption and a reduced need for attentional control as subcortical networks become more active. Analysis of gait data indicated that gait variability did not change with repetitive task exposure, supporting the second explanation.
Relationship Between Gait Characteristics, Cognitive Function, and Cortical Activation
In older adults, we observed consistent negative correlations between relative HbO2 and changes in gait variability in all motor regions (see Table 3). Individuals with the largest decrease in temporal and spatial gait variability showed the largest increase in activation of cortical motor regions similar to a previous study involving cued foot movements. 11 In contrast, the opposite was observed in young adults (Figure 3). Three possible explanations are as follows: (a) the temporal demands of synchronizing stepping movements to the beat come at a higher motor cost in older adults requiring greater cortical motor activation to synchronize stepping movements to the beat (and therefore reduced variability). (b) Older adults employ different motor control strategies with greater muscle coactivation than younger adults during usual walking 48 resulting in increased joint stiffness. Agonist muscles must contract more strongly to achieve the desired movement, which is associated with greater cortical activity. 49 The increased joint stiffness, both active and passive, together with increased damping (dissipation of energy) will partly determine gait variability. In the absence of sudden or high frequency perturbations, 50 increased joint stiffness may be associated with increased stability and decreased gait variability. (c) The explanation for the positive correlation between gait variability and cortical activity in younger adults may be associated with control of walking relying more on automatic pathways. Higher cortical activity in young individuals may be an indication of dysfunctional automatic pathways and consequentially greater gait variability.
Depressive symptoms and global cognitive status influence the cortical response to RACs during walking in older adults but not young adults. Our results showed that less severe depressive symptoms and better cognitive function were associated with increased cortical activation during RAC walking. These findings are consistent with studies showing reduced activation of the PFC and/or SMA during cognitive tasks in patients with major depressive disorder 24 and cognitive impairments. 51 As the older adults enrolled in the current study were high-functioning, independent and cognitively preserved (based on MoCA score), the observed correlations might be an indication of early functional changes in cortical activation in those with very mild cognitive declines.
Study Limitations
Limitations apply while interpreting the current findings. First, treadmill walking may not represent overground walking. The treadmill acts as a pace maker, dictating the rhythm of walking. However, the fact that we observed changes in gait variability and cortical changes, given the constraints of treadmill walking, strengthens the association. A second limitation is that our sample included high-functioning older adults, which limits the generalization of current findings. Additionally, the test we selected for global cognition, the MoCA, is semiquantitative and has a ceiling effect. The moderate correlations we reported between cognition and cortical activity may be higher if we selected a test where scores were normally distributed or recruited participants with a wider range of MoCA scores. A third limitation is that short separation channels were not used, which may have reduced scalp blood flow artifact. 52 Short separation channels have been recently suggested to correct the fNIRS signal for hemodynamic changes in superficial tissue layers. However, the impact on our findings should be minimal as the treadmill speed was kept constant for both walking conditions. Additionally, as older adults walked slower than young adults, we used a statistical model with treadmill speed as a covariate. Finally, the differential path length factor, which is a correction factor that must be incorporated when the Lambert-Beer law is applied to biological tissues, is not known for adults older than 50 years. Consequently, the same differential path length factor is commonly used for those older than 50 years. This limitation applies for all fNIRS studies involving older adults. Despite these limitations, this study advances the current knowledge about the effects of aging on the activation of cortical regions during walking to RAC.
Clinical Implications
This study demonstrates that in older adults RAC results in increased activity across multiple cortical areas and decreased gait variability. Although a novel task such as cued treadmill walking is attentionally demanding as supported by increased PFC activity, the increase attenuates to levels similar to younger adults in a fairly short time period, demonstrating adaption. Clinicians applying RACs to aid older adult gait should conduct multiple trials to ensure that the cue is integrated into automatic gait control. The cognitive and depressive status of the older adult should be assessed prior to administering cueing therapy, as these may adversely affect the response to cues.
Conclusions
In conclusion, this novel study demonstrated positive compensation strategies employed by older adults when walking to an RAC resulting in improved gait performance (reduced gait variability). Additionally, results confirmed the involvement of the PFC in the control of walking to rhythmic auditory cues in young and older adults, with a significant reduction in activity following repeated exposure indicating a training effect. Finally, this study highlights the interaction between cognitive capacity, increased cortical activity and changes in gait parameters, stressing the need to take a holistic multisystem approach when managing gait in the older adult.
Supplemental Material
Supplementary_Table_1_Slope_cortical_areas – Supplemental material for Reduced Gait Variability and Enhanced Brain Activity in Older Adults With Auditory Cues: A Functional Near-Infrared Spectroscopy Study
Supplemental material, Supplementary_Table_1_Slope_cortical_areas for Reduced Gait Variability and Enhanced Brain Activity in Older Adults With Auditory Cues: A Functional Near-Infrared Spectroscopy Study by Rodrigo Vitorio, Samuel Stuart, Lilian T. B. Gobbi, Lynn Rochester, Lisa Alcock and Annette Pantall in Neurorehabilitation and Neural Repair
Supplemental Material
Supplementary_Table_2_Gait – Supplemental material for Reduced Gait Variability and Enhanced Brain Activity in Older Adults With Auditory Cues: A Functional Near-Infrared Spectroscopy Study
Supplemental material, Supplementary_Table_2_Gait for Reduced Gait Variability and Enhanced Brain Activity in Older Adults With Auditory Cues: A Functional Near-Infrared Spectroscopy Study by Rodrigo Vitorio, Samuel Stuart, Lilian T. B. Gobbi, Lynn Rochester, Lisa Alcock and Annette Pantall in Neurorehabilitation and Neural Repair
Footnotes
Acknowledgements
Authors acknowledge the contribution of Ellen Lirani-Silva, MSc, with data collection and Dr Silvia Del Din with analysis of accelerometer data.
Authors’ Note
The views expressed are those of the authors and not necessarily those of the National Health Service, the National Institute for Health Research, or the Department of Health.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institute for Health Research (NIHR) Newcastle Biomedical Research Unit (BRU) and centre (BRC) based at Newcastle upon Tyne Hospitals NHS Foundation Trust and Newcastle University. The research was also supported by NIHR Newcastle CRF Infrastructure funding and Sao Paulo Research Foundation (FAPESP 2016/00518-8; postdoctoral fellowship to Rodrigo Vitorio).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.