
Abstract
Background. Motor unit firing frequencies are low during maximal voluntary contractions (MVCs) of human thenar muscles impaired by cervical spinal cord injury (SCI). Objective. This study aimed to examine whether thenar motor unit firing frequencies increase when driven by both maximal voluntary drive and other concurrent inputs compared with an MVC alone. Methods. Motor unit firing rates, force, and surface electromyographic activity (EMG) were compared across 2 contractions: (a) MVC alone and (b) MVC combined with another input (combination contraction). Other inputs (conditions) included vibration, heat, or cold applied to the anterior surface of the forearm, electrical stimulation delivered to the anterior surface of the middle finger, a muscle spasm, or a voluntary contraction of the contralateral thenar muscles against resistance. Results. The maximal firing frequency (n = 68 units), force, and electromyographic activity (n = 92 contraction pairs) were all significantly higher during the combined contractions compared with MVCs alone. There was a 3-way interaction between contraction, condition, and subject for maximal motor unit firing rates, force, and EMG. Thus, combined contraction responses were different for conditions across subjects. Some conditions (eg, a muscle spasm) resulted in more effective and more frequent responses (increases in unit firing frequency, force, EMG in >50% contractions) than others. Recruitment of new units also occurred in combined contractions. Conclusions. Motoneurons are still responsive to additional afferent inputs from various sources when rate modulation from voluntary drive is limited by SCI. Individuals with SCI may be able to combine inputs to control functional tasks they cannot perform with voluntary drive alone.
Keywords
Introduction
After human spinal cord injury (SCI), many descending inputs from higher centers are interrupted. It is unknown whether the affected synaptic sites on motoneurons remain vacant after injury or become occupied by afferent and/or spinal inputs. Yet the end result has the potential to influence muscle function and future repair by axon regeneration. If the source or strength of inputs to motoneurons changes following injury, it may be possible to assist voluntary muscle contractions by activation of unconventional inputs.
During a maximal voluntary contraction (MVC), force is mainly generated by gradation of motor unit firing rate.1-4 In control thenar motor units, for example, stimulation at 5 Hz (recruitment rates) evokes 6% to 51% of maximal unit force. 4 The remaining 49% to 94% of force must be generated by rate modulation. In hand muscles, recruitment is complete by about 50% MVC.5,6 These results emphasize the importance of rate modulation to achieve higher forces. 7
Cervical SCI changes this situation. Motor unit recruitment becomes much more important for force production in hand muscles. First, recruitment occurs over an expanded range of forces (up to 85% MVC).8,9 Second, stimulation at recruitment rates (5 Hz) results in stronger unit forces (14% to 64% maximal), reducing the force contribution from firing rate increases. 10 Third, rate modulation is limited; after recruitment, motor units show little or no increase in firing rate. Consequently, mean MVC firing rates are low (9 ± 3 Hz) compared with those recorded from motor units of uninjured individuals (34 ± 10 Hz).9,11 Whether the low unit firing rates after SCI reflect firing rate saturation is unclear. Two results suggest that thenar unit firing frequencies are submaximal. First, the firing rate of the same motor unit is higher during an involuntary contraction (muscle spasm) than during an MVC. 9 Second, stimulation at 30 to 100 Hz is necessary to evoke maximal forces in thenar motor units.4,10
Here, our aim was to examine whether individuals with cervical SCI can combine voluntary drive with other afferent inputs to enhance maximal thenar motor unit firing rates. Any increase in maximal motor unit firing rates should enhance force, which may improve function.
Methods
Maximal voluntary contractions of thenar muscles were measured in one hand of 8 individuals (7 men, 1 woman, age range, 21-61 years; Table 1) with chronic (>1 year) cervical SCI. Neurological level of injury and injury completeness were defined by the American Spinal Injury Association criteria. 12 All but one subject had some voluntary control of thenar muscles in at least one hand (see Table 1). If the thenar muscles of both hands were under voluntary control, the hand with the weaker muscles was studied. Weaker muscles were assumed to have fewer motor units under voluntary control, making it easier to follow units up to maximal force. Subjects gave written consent to participate in the study, which was approved by the University of Miami Institutional Review Board.
Subjects’ History a
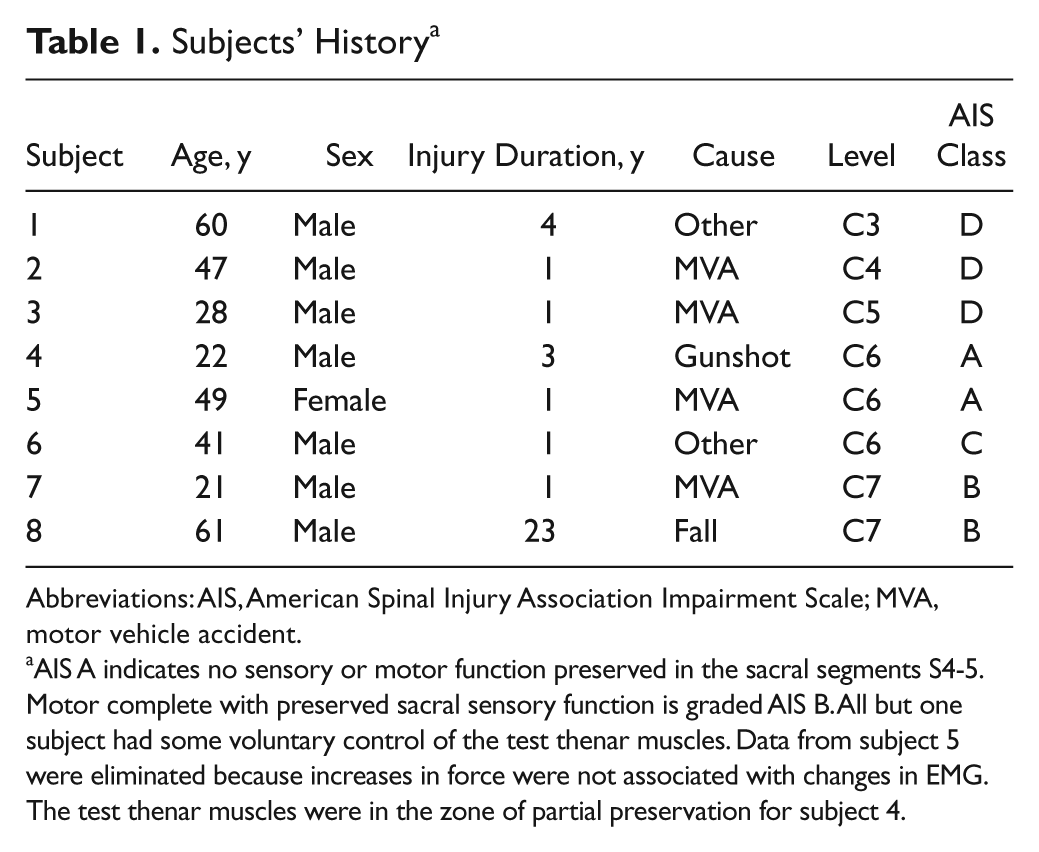
Abbreviations: AIS, American Spinal Injury Association Impairment Scale; MVA, motor vehicle accident.
AIS A indicates no sensory or motor function preserved in the sacral segments S4-5. Motor complete with preserved sacral sensory function is graded AIS B. All but one subject had some voluntary control of the test thenar muscles. Data from subject 5 were eliminated because increases in force were not associated with changes in EMG. The test thenar muscles were in the zone of partial preservation for subject 4.
Experimental Setup
The subject remained seated in his or her wheelchair. As described earlier, 13 the elbow was flexed to 90°. The forearm rested on a platform beside the chair and was stabilized in a vacuum cast. The hand rested palm up between layers of TheraPutty (North Coast Medical, Gilroy, CA). A metal plate was placed on top of the hand and fixed with Velcro straps. The thumb was extended and rested against a custom-made force transducer that registered abduction and flexion forces. Resultant forces were calculated using the following formula: resultant force = √[(abduction force)2 + (flexion force)2]. Surface electromyographic activity (EMG) was recorded from the distal and proximal portions of the thenar muscles. 14 The distal electrode was placed over the interphalangeal joint, whereas the proximal electrode was placed at the base of the thenar eminence. A common electrode lay across the middle of the muscle bellies. The ground electrode was placed across the arm, just proximal to the wrist. Motor unit potentials were recorded intramuscularly with a custom-made tungsten microelectrode. Temperature was monitored from the surface of the skin on the upper arm throughout the experiment.
Protocol
Initially, the subject was instructed to completely relax and stop any thenar motor unit activity. This was only possible for some units. Subjects then performed a series of MVCs, in pairs, to determine whether motor units could be driven more effectively by voluntary drive (MVC), or by voluntary drive in combination with other inputs (combined contraction). For an MVC alone, subjects were instructed to contract their thenar muscles voluntarily from rest to maximum force in 5 seconds, maintain force at maximum for 5 seconds, then decrease the force to rest over 5 seconds. For a combined contraction, subjects were asked to generate an MVC as described before, while another input was applied simultaneously throughout the voluntary contraction. The other inputs (conditions) included (a) vibration (130 Hz) applied to the middle of the anterior surface of the forearm, (b) a heat pad applied to the same area, (c) an ice pack applied to the same area, (d) electrical stimulation (100 Hz, pulse duration 1 ms) delivered to a metal ring around the middle finger, (e) an investigator lifting the knee or pushing the shoulder backward to trigger a spasm that activated thenar muscles, and (f) a voluntary contraction of the contralateral thenar muscles against resistance. One minute of rest was provided between each contraction. The order of the 2 contractions was reversed between conditions to avoid order effects.
Application of other inputs alone failed to elicit activation of the test thenar muscles with one exception. In one subject, involuntary contractions of leg muscles produced both EMG and force in thenar muscles but data were obtained for only 3 motor units. These data were not included in the results.
Data Collection and Analysis
Intramuscular EMG, surface EMG, and force were amplified, filtered (30 Hz to 10 kHz, 30 Hz to 3kHz, DC-100 Hz, respectively), sampled online using a CED 1401 data-acquisition interface (20 kHz, 2 kHz, 0.5 kHz, respectively) and analyzed offline using Spike 2 software (Cambridge Electronic Design Limited, Cambridge, UK).
Point-by-point running averages (over 500 ms) of the rectified surface EMG and resultant force were plotted against each other. Force increases had to be accompanied by increases in EMG to indicate that the force increase was produced by a general increase in excitatory drive to the thenar motor pools. Data from one subject were eliminated because increases in force were not associated with changes in EMG.
Potentials belonging to the same motor unit were identified using templates based on amplitude and shape. To ensure that the same motor unit was followed, the waveforms for each unit were overlaid for each contraction and compared across contractions (Figure 1). Two researchers verified the accuracy of triggering individual motor unit potentials. The surface EMG signals time-locked to the intramuscular potentials were also averaged and overlaid. In some cases, a small motor unit potential exhibited sustained firing after most contractions (Figure 1B), suggesting that the position of the recording electrode was stable across contractions. Recruitment of a new unit could also occur without any change in the shape of the potentials or firing behavior of an already active unit (Figure 1C).
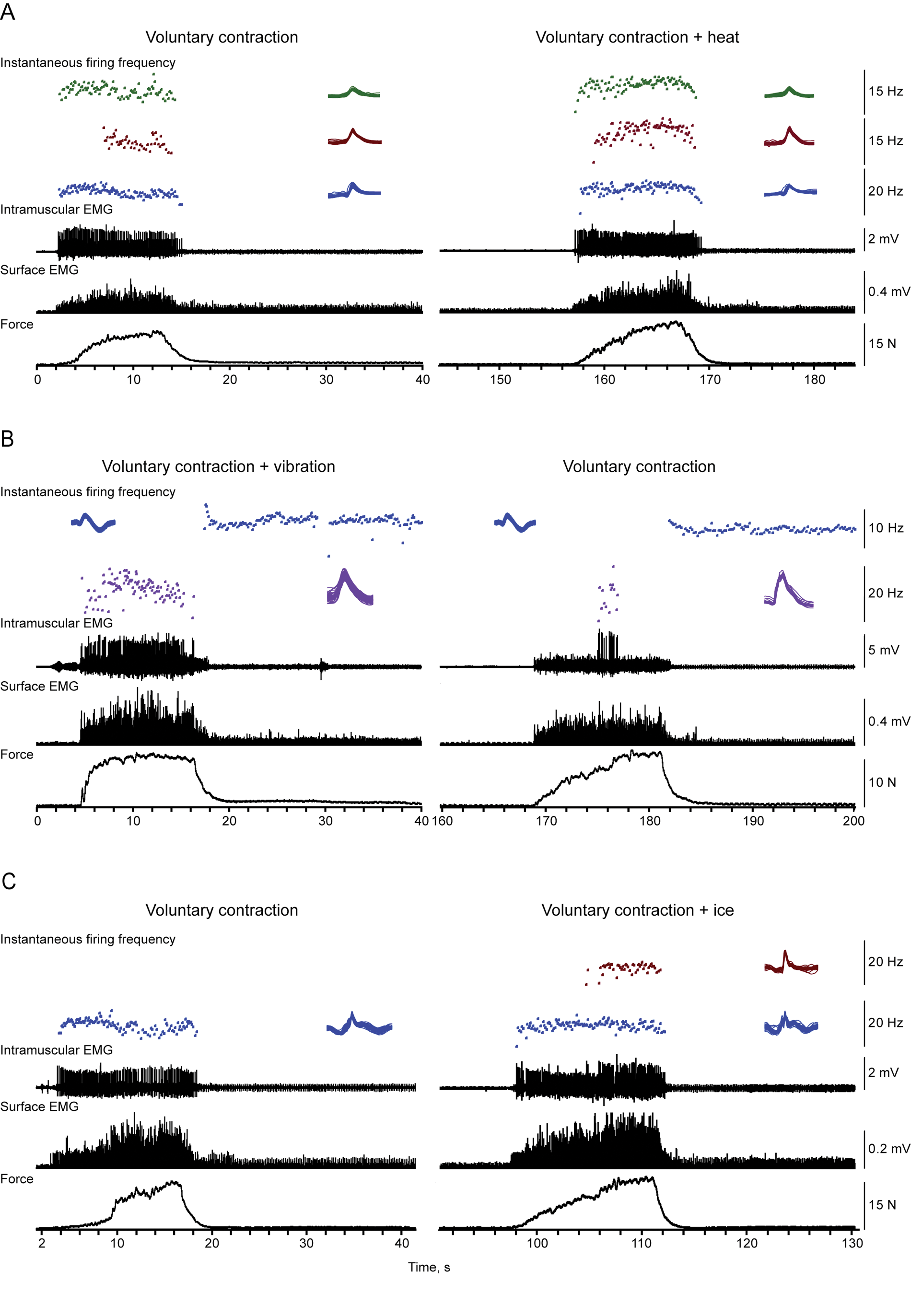
Motor unit firing behavior, electromyographic activity (EMG), and force across contractions. Instantaneous motor unit firing rates, intramuscular EMG, distal surface EMG, and force (top to bottom) recorded during 3 pairs of contractions. Maximal voluntary contractions (MVCs) were paired with combined contractions involving an MVC and heat (A), an MVC and vibration (B), or an MVC and ice (C). Contraction order varied across the pairs. Overlays of the potentials for motor units followed throughout a pair of contractions verify that the unit identification was accurate. Some motor units continued to fire after the contractions but were not always identifiable at stronger forces. Motor unit recruitment occurred with ice.
For each contraction, maximal surface EMG (maximal rectified value over 500 ms), maximal force (maximal value over 500 ms), and maximal motor unit firing frequency (mean frequency over 3 consecutive interspike intervals) were determined. Each parameter was considered to increase for the combined contraction if the value exceeded the MVC alone data by 5%, to decrease if the reduction exceeded 5% of the MVC alone value, or to show no change if the value changed by ≤5% in either direction (within 95% and 105% MVC alone).
Statistical Analysis
Maximal motor unit firing rate, surface EMG, and force were compared for the 2 contractions (MVC alone, combined contraction) by expressing data for the combined contraction as a percentage of the MVC alone nearest in time. Data were analyzed with repeated-measures analysis of variance with contraction type as within-subject parameter (2 levels: MVC only, combined contraction), and condition (6 levels: vibration, bilateral, ice, heat, stimulation, spasm) and subject (7 levels) as between-subject parameters. Data are expressed as mean ± standard deviation (mean ± SD). Values were considered to be significantly different at P < .05.
Results
For each motor unit, the MVC firing rate was always compared with that recorded when another input was combined with an MVC. For example, during an MVC and heat (Figure 1A), 3 units fired at higher maximal rates for longer times than during an MVC alone; a change associated with greater force, better force retention, and an increase in surface EMG. An MVC with vibration resulted in an increase in maximal unit firing rate and surface EMG compared with an MVC alone, as well as a more sustained maximal firing rate and force (Figure 1B). In contrast, the maximal firing rate of one unit declined during an MVC and ice versus an MVC alone (Figure 1C). However, another unit was recruited only during the combined MVC, resulting in increases in both force and EMG. Together, these examples show an increase in muscle output when inputs are combined, as well as successful maintenance of higher forces.
Motor Unit Firing Behavior
An increase in maximal force and EMG can occur both by recruitment of inactive motor units and by increasing the firing frequency of already active units. We were able to follow 68 units throughout both an MVC and then a combined contraction (or vice versa). Overlays of the potentials for each contraction in Figure 1 show matching shapes for a given unit, indicative of accurate unit identification across contractions. Other units were only active during the voluntary contraction or during combined contractions. In some cases, differences in unit activity across contractions reflected recruitment of new units. For example, potentials belonging to a new unit(s) appeared in the intramuscular record and were accompanied by increases in surface EMG (Figure 1C). In other cases, additional unit recruitment resulted in force increases and unitary potentials in the surface EMG. This recruitment occurred without affecting the shapes of the potentials of already active units.
Figure 2A shows the distribution of units with an increase, no change, or a decrease in firing frequency during combined contractions. Increases in motor unit firing frequency occurred in more than half of the units when an MVC was combined with a spasm or vibration, whereas frequency decreased in the ice condition for all of the units tested.
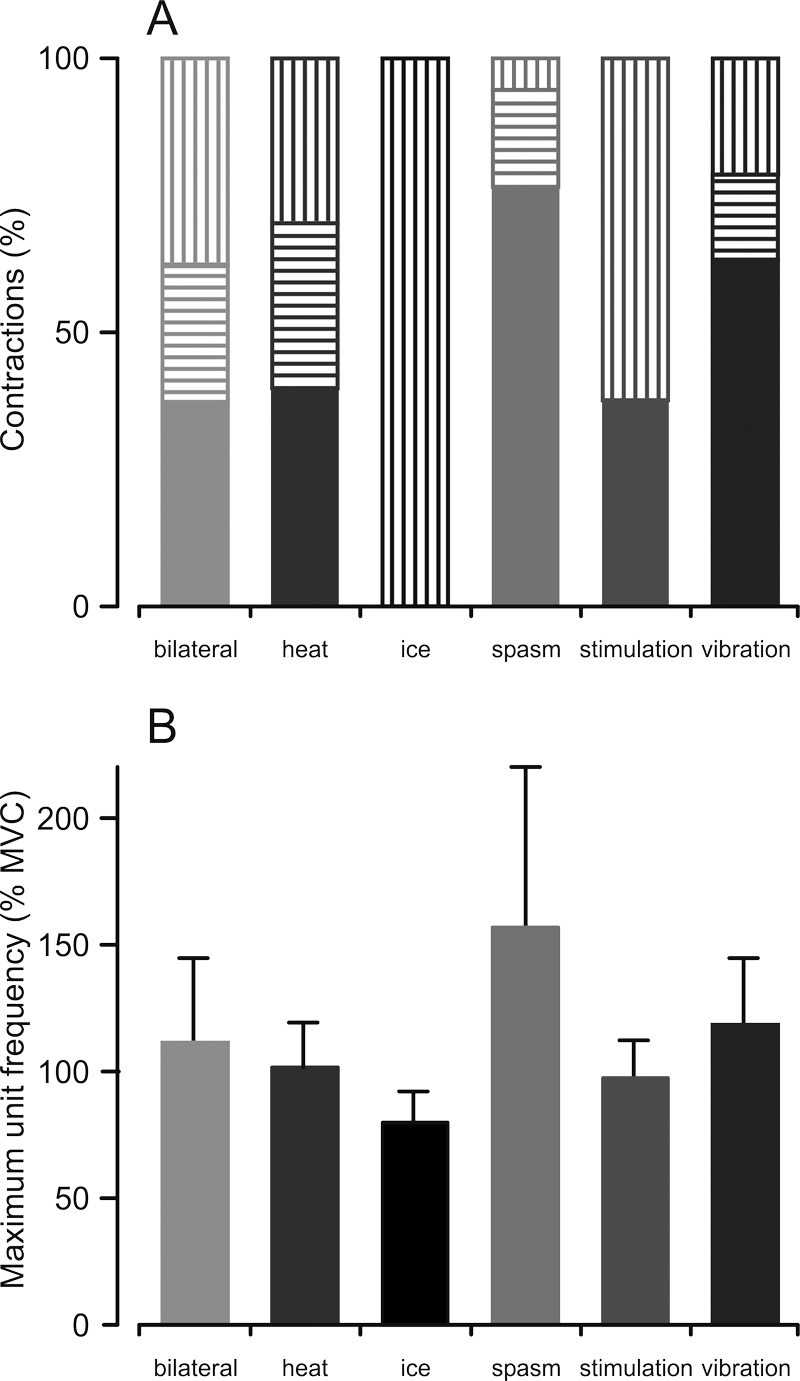
Maximal motor unit firing frequency. (A) Percentage of motor units that show an increase (solid), no change (horizontal stripes), or a decrease (vertical stripes) in maximal firing frequency during the combined contraction compared with maximal voluntary contraction (MVC) alone. (B) Mean (+SD) maximal motor unit firing frequency when an MVC was combined with another input (% frequency during MVC only). A significant 3-way interaction between contraction, condition, and subject was observed (F(9, 49) = 3.180, P = .004).
The maximal unit firing frequency during an MVC alone (11.7 ± 4.0 Hz), was significantly lower than the firing frequency during the combined contractions (13.4 ± 5.2 Hz; main effect of contraction: F (1, 47) = 11.529, P = .001). The magnitude of the increase in maximal unit frequency differed across conditions (F(5, 47) = 4.196, P = .003) and subjects (F(4, 47) = 8.228, P < .001). Additionally, there was a 3-way interaction between contraction, condition, and subject (F(9, 49) = 3.180, P = .004), indicating that the effect of the various conditions differed among subjects. The largest mean increases in unit firing frequency occurred when an MVC was combined with a spasm or vibration, whereas a decrease in frequency occurred when an MVC was combined with ice (Figure 2B).
Force and Surface EMG
The maximal force that subjects could produce voluntarily (MVC) ranged from 5.1 to 31.8 N. In the combined contractions, maximal force ranged from 5.5 to 41.9 N. In 6 of the 7 subjects, the maximal force in the combined contractions increased beyond that produced by an MVC alone (>20% in 2 subjects; >5% in 4 subjects). Maximal force and EMG were measured for 92 pairs of contractions. The combined contractions resulted in stronger force (112.1% ± 32.4% MVC; main effect of contraction: F(1, 53) = 14.367, P < .001) and EMG (118.0% ± 42.7% MVC; F(1, 52) = 18.438, P < .001).
In individual trials, the force produced during the combined contraction could increase, remain similar, or decrease in comparison with the MVC alone (Figure 3A). Combining an MVC with a spasm or ice increased the force in more than 50% of the contractions. The other conditions were less conclusive because the effects were similarly distributed between increase, no change, and decrease (~30% each).
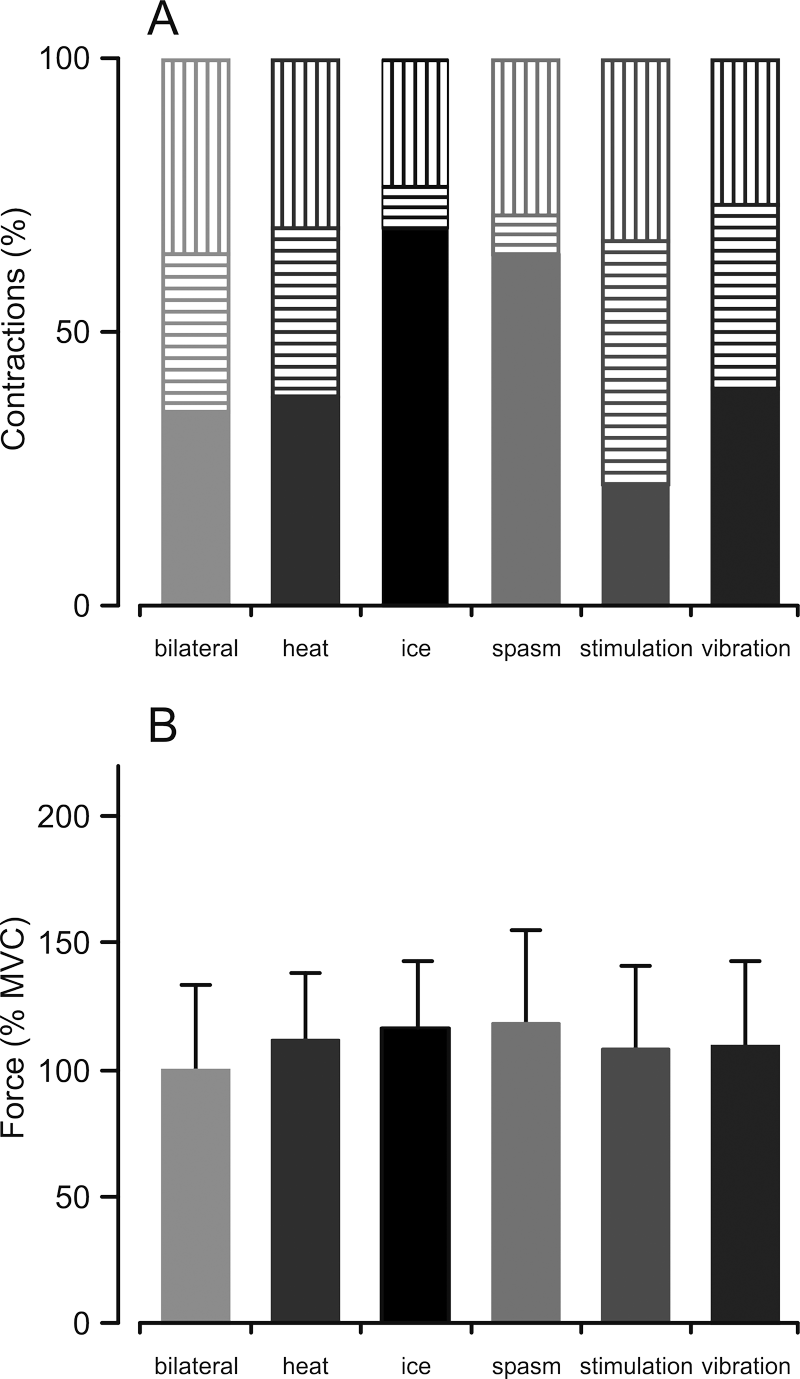
Maximal force. (A) Percentage of contractions that showed an increase (solid), no change (horizontal stripes), or a decrease (vertical stripes) in maximal force when a maximal voluntary contraction (MVC) was combined with another input compared with an MVC alone. (B) Average (+SD) force during combined contractions (%MVC alone). A significant 3-way interaction between contraction, condition, and subject was observed (F(27, 53) = 2.554, P = .002).
The force differed across conditions (contraction × condition, F (5, 53) = 3.314, P = .011; Figure 3B) and subjects (interaction effect: contraction × subject, F(6, 53) = 14.222, P = .042). Moreover, there was a differential effect of condition among subjects (interaction effect: contraction × condition × subject, F(27, 53) = 2.554, P = .002). Thus, certain inputs increased force in some subjects more than other inputs.
For surface EMG, the distribution of effects across conditions (increase, no change, decrease) was similar to the distribution for the maximal forces (Figure 4A). Inputs from a spasm or ice, as well as vibration, increased EMG in more than 50% of the contractions, whereas there were no clear trends for the other conditions. For EMG, there was an effect for contraction (F(1, 52) = 18.438, P < .001), condition (contraction × condition, F(5, 52) = 4.463, P = .002), and subject (interaction effect: contraction × subject, F(6, 52) = 2.967, P = .015). A 3-way interaction between contraction, condition, and subject was also found (interaction effect: contraction × condition × subject, F (26, 52) = 2.524, P = .002). Overall, combining an MVC with a spasm or ice was most effective for increasing EMG (Figure 4B) and force.
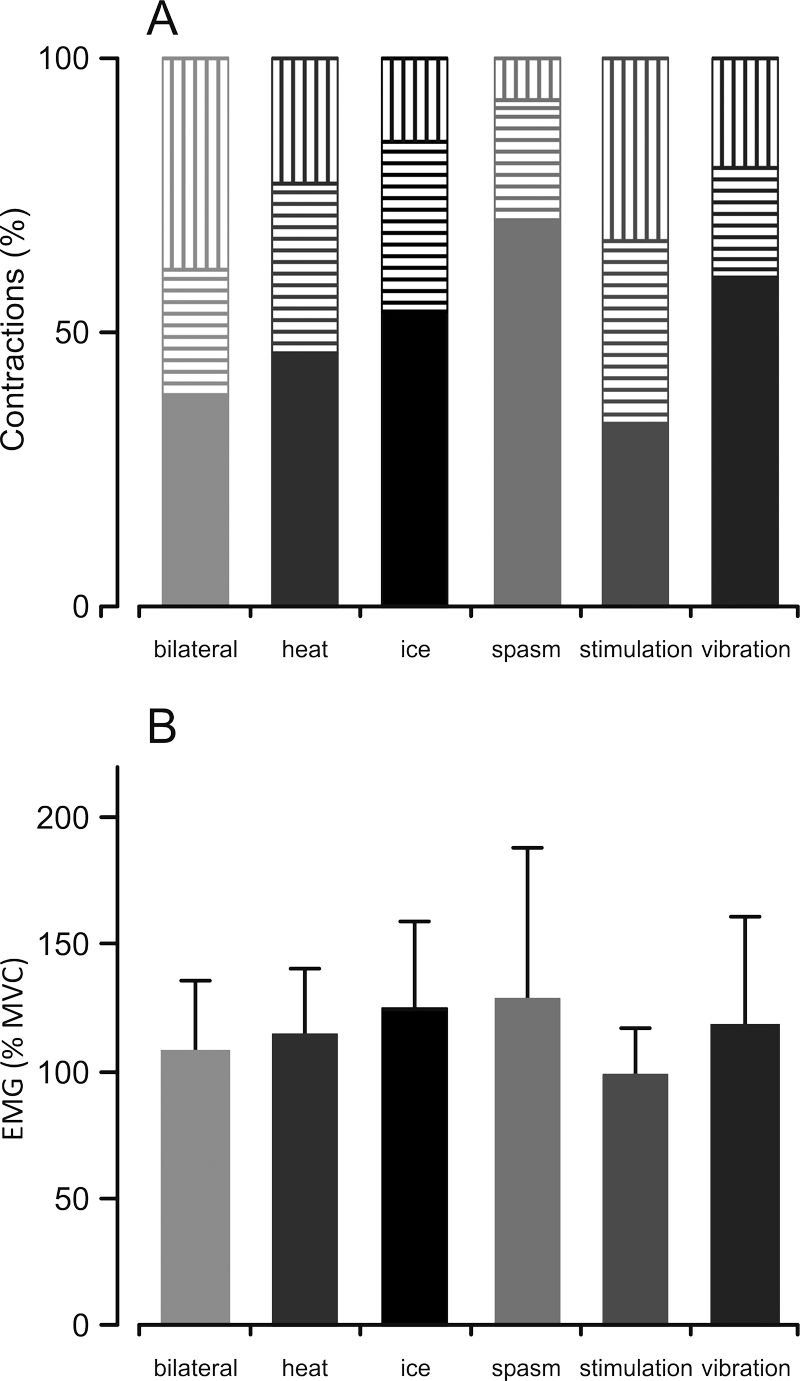
Maximal electromyography activity (EMG). (A) Percentage of contractions that showed an increase (solid), no change (horizontal stripes), or a decrease (vertical stripes) in maximal EMG when a maximal voluntary contraction (MVC) was combined with another input compared with an MVC alone. (B) Average (+SD) EMG during combined contractions (%MVC alone). A significant 3-way interaction between contraction, condition, and subject was observed (F(26, 53) = 2.524, P = .002).
Discussion
Motor unit rate modulation is limited during voluntary contractions of thenar muscles that have been partially paralyzed by cervical SCI. Low maximal firing frequencies were found for all units irrespective of whether they were recruited at either low or high forces. 9 We show that these low motor unit firing frequencies do not reflect firing rate saturation. Motor unit firing frequencies, force, and surface EMG all increased significantly when an MVC was combined with additional inputs from various sorts of afferents. These inputs also induced additional motor unit recruitment.
Small Increases in Unit Firing Rates Enhance Muscle Force
The mean increase in maximal motor unit firing rate for all of the combined contractions was 2 Hz. Although this mean change is small, the increase in force may be considerable. The low firing rates of these units (13.4 Hz) indicates that they are operating on the steep part of the force–frequency curve (from 5 to 20 Hz) where small frequency changes induce large relative force changes. Estimates from paralyzed and control thenar motor units show that force will increase by an average of 3% and 4% maximal for every 1 Hz change in frequency, respectively.4,10 Furthermore, people with cervical SCI who can use triceps brachii against gravity generate an average of 8% of control (uninjured) maximal force. 15 Since most tasks do not require maximal force, a 6% force increase in thenar muscles may improve function. Using a smaller percentage of maximal force may also make a task less fatiguing, which would help the subject.
Deficits in Descending Inputs Contribute to Low Thenar Motor Unit Firing Rates After Cervical SCI
Insight into the factors that contribute to low motor unit firing rates in thenar muscles after SCI comes from comparing the maximal firing rate of a unit to its rate when it continues to fire after a contraction (Figure 5A). Such sustained motor unit firing is often seen in hand muscles after SCI9,16,17 and is usually attributed to persistent inward currents.18-21 These currents are activated just below threshold voltage and amplify synaptic input. Deactivation of persistent currents occurs in response to short inhibitory inputs.20-25 For those units that demonstrated sustained firing following an MVC or a combined contraction, mean maximal firing rates were 10.3 ± 0.5 Hz and 15.1 ± 0.5 Hz, respectively. After each of these contraction types, the firing rates of these same units dropped to 6.1 ± 0.4 Hz and 6.4 ± 0.8 Hz, respectively. Thus, after both types of contractions, the sustained firing was comparable, indicating that the putative contribution from activated persistent inward currents was similar. The respective maximal firing rates, however, differed. The difference between the MVC and postcontraction sustained firing rates largely represents the rate increase resulting from maximal voluntary effort (4.2 ± 0.4 Hz; although the voluntary input may also grade the persistent inward current). Thus, the estimated MVC contribution to the overall firing rate (~40%) was lower than that seen during the sustained firing (~60%). In contrast, the difference between the rates measured during the combined contractions and the sustained firing rates (8.7 ± 0.5 Hz) suggested that ~40% of the overall unit firing rate arose from sustained firing. Similar values occur in uninjured subjects. 26 Together, these results reinforce that thenar motoneurons are still responsive to additional inputs after SCI, and intrinsic motoneuron properties make important contributions to muscle force production.
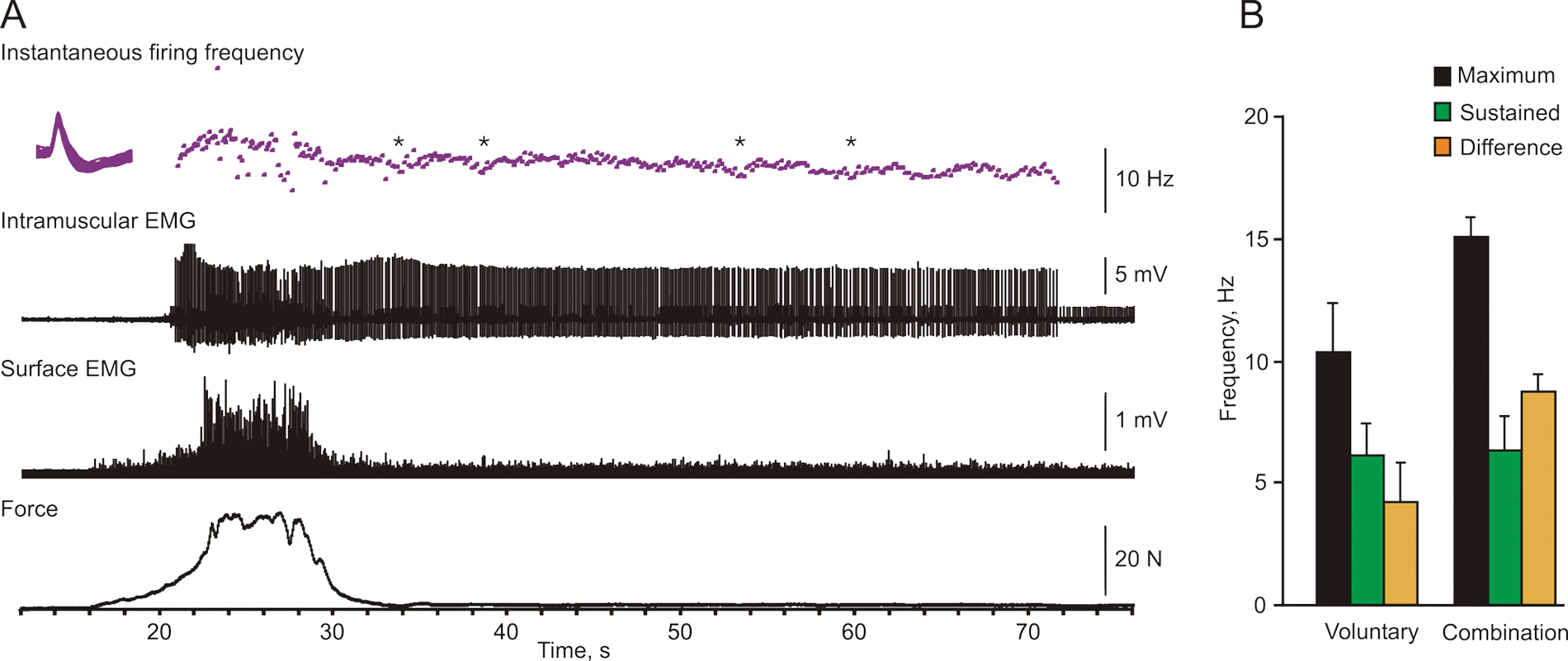
Sustained motor unit firing. (A) Overlays of the potentials and instantaneous firing rate for one motor unit, intramuscular electromyographic activity (EMG), distal surface EMG, and force (top to bottom) during a maximal voluntary contraction (MVC) with ice. The maximal motor unit firing rate was 15.2 Hz. Unit firing was sustained after the contraction at an average of 7.3 Hz. Asterisks denote the times when the subject made an extra effort to stop the motor unit firing. (B) Mean (+SD) maximal, sustained, and difference (maximum − sustained) unit firing rate for units that showed sustained firing after an MVC (n = 15) or a combined contraction (n = 3). Some data for MVCs came from Zijdewind and Thomas. 9
The sustained motor unit firing after some contractions resembles the firing characteristics of rat motoneurons after spinal cord transection. 21 This behavior represents the combination of a regenerative, persistent sodium current and a persistent calcium current. Synaptic input could be the driving force for regeneration of sodium persistent currents in this study as all of the subjects had some voluntary control of the test thenar muscles. One indication of this possibility is the ability of subjects to reduce unit firing frequency even though they were unable to stop the unit firing (see Figure 5). Inhibition is unlikely to underlie the decline of unit firing because an inhibitory input to the motoneuron pool would likely deactivate the persistent currents.
The higher unit firing rates with combined inputs may represent the effects of existing excitatory synapses, synapses that are unmasked and/or strengthened by SCI, or less inhibition. Axons may also sprout and make novel connections with motoneurons in response to the stripping of synapses that occurs with injury.27-31 If this is the case, new inputs may hinder subsequent synapse formation from regenerating axons.
Why Are Motor Unit Firing Rates Low in Hand Muscles After Cervical SCI?
Both the present data and our previous study 9 show that motoneurons receive enough input to be recruited at relatively low or high force levels. However, when voluntary effort is increased to maximum, the increases in unit firing rate are always small. Even when additional inputs enhance unit firing rates, overall rate modulation is reduced after SCI since MVC thenar unit firing rates averaged 34 Hz in uninjured subjects. 11 Limited rate modulation during voluntary contractions is not unique to SCI. Low maximal voluntary firing rates have also been observed in units of people with multiple sclerosis, 32 and low submaximal firing rates have been reported after stroke. 33 Whether the same mechanisms are responsible in these examples is unclear. Low maximal voluntary firing rates after SCI also seem specific to hand muscles. Data from motor units in triceps brachii of SCI subjects show normal to high firing rates. 34
Motoneurons to hand muscles are unique because they strongly depend on direct corticospinal input for activation. 35 A decline in both the number and strength of these projections to motoneurons (and interneurons) is plausible after SCI. Additionally, less than half of the cortical pyramidal cells increase their firing rate with an increase in force.36,37 Other pyramidal cells are more sensitive to kinematic-related parameters.37,38 These observations imply that recruitment of specific cortical cells is necessary to increase synaptic input to spinal motoneurons by voluntary effort. The numbers of cortical cells that remain connected to the spinal cord (or the number of cortical cells that modulate their firing with muscle force) may be insufficient after injury to ensure synchronized inputs to spinal motoneurons. This may reduce the magnitude of motor unit firing rate increases. Furthermore, central demyelination after human SCI 39 may temporally disperse the input to motoneurons, reduce the chance of action potential generation, and limit motor unit firing rates, as is probably the case in multiple sclerosis. 32
Besides less cortical control over motoneuron pools, it is common for motoneurons (and interneurons) to die near the lesion.40-44 Remaining axons can sprout within the spinal cord and make new connections.27-31 Hence, the traumatic incident ought to result in reorganization of the input to motoneuron pools. In able-bodied subjects, inputs from various sources (eg, corticospinal, reticulospinal, rubrospinal tracts) are unevenly distributed over the motoneuron pool. 45 Thus, a dramatic change in the balance between excitation and inhibition may change the gain of the motoneuron input–output relation,7,46,47 making the motoneurons less responsive to synaptic input. Activation of persistent inward currents will also make it more difficult to increase motor unit firing rates because of saturating effects. 48 Another possible explanation for the low maximal firing frequency after SCI may relate to differences in the relative location of remaining excitatory and inhibitory synapses. Excitatory synapses further away from the soma or close to inhibitory synapses will have smaller depolarizing effects.7,49,50
The effects of SCI on motor unit firing rates become even more complicated when cellular and molecular changes are considered. In animal experiments, SCI results in changes in intrinsic motoneuron properties (eg, differences in the threshold for action potential generation21,51; alterations in the expression of genes that influence motoneuron excitability 52 ; constitutive activation of 5-HT2C receptors 53 ; and declines in postsynaptic inhibition 54 ). If one considers all of the changes that could occur after SCI, combined with differences in the extent, nature, and location of the trauma, it is not surprising that individual motor units responded differently during combined contractions.
Combining Voluntary Drive With Afferent Inputs May Increase Muscle Function
The overall increase in motor unit firing frequency, force, and EMG with the addition of various sorts of input during MVCs, together with recruitment of new motor units, stresses the potential of coupling extra input to voluntary effort to improve muscle function. Both of the primary mechanisms for force modulation are targeted (motor unit recruitment and rate modulation). At the same time, control of the contraction is maintained voluntarily (Figure 1). This strategy of combining inputs during muscle function allows immediate facilitation of output throughout the contraction, as it is needed. Other studies have paired afferent and transcranial magnetic stimulation to transiently increase corticospinal output in SCI subjects with intact sensation, 55 which may facilitate function or have used training interventions that chronically couple peripheral nerve stimulation with functional tasks to improve subsequent hand function after SCI. 56
The effects of combining different sources of input strongly depended on the input and varied across subjects. These observations suggest that this kind of rehabilitation would have to be individually shaped for people with chronic injury. During the early phase of rehabilitation, it may be beneficial to introduce additional afferent input to increase synaptic strength or to maintain those intrinsic motoneuron properties essential for rate modulation. Here we present motor unit firing rate, force, and EMG data that provide the physiological basis for the strategy that spinal cord injured individuals may be able to learn to combine inputs (trigger a spasm with a voluntary contraction) to control functional tasks that they cannot perform with either input alone.
Footnotes
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the University Medical Center Groningen, the National Institutes of Health (NS-3-2351; NS-30226), and The Miami Project to Cure Paralysis.