
Abstract
Background. Environmental enrichment (EE) defined as “a combination of complex inanimate and social stimulation” influences brain function and anatomy by enhancing sensory, cognitive, motor, and social stimulation. The beneficial effects of EE in the presence of brain damage have been partially attributed to upregulation of neurotrophins, proteins involved in neuronal survival and in activity-dependent plasticity. Objective. The authors tested the hypothesis that EE may have advantageous effects on recovery of motor function after cerebellar damage, associated with changes in local neurotrophin production. Methods. They performed a hemicerebellectomy in rats previously exposed to EE or reared in standard conditions. The time course of compensation of motor symptoms was analyzed in both lesioned groups. Then, the local production of the nerve growth factor (NGF) and brain-derived neurotrophic factor (BDNF) in the spared hemicerebellum and other extracerebellar regions was evaluated. Results. Long-term exposure to EE accelerated the motor recovery in hemicerebellectomized rats and elicited an increase in NGF levels in the spared hemicerebellum, as compared with nonenriched lesioned and control rats. BDNF levels were higher in hemicerebellectomized rats but not influenced by EE. In the frontal cortex, both NGF and BDNF levels were upregulated in hemicerebellectomized enriched rats as compared with hemicerebellectomized nonenriched and control rats. Conclusions. This study suggests that the beneficial effects of EE on motor symptoms after cerebellar damage may be, at least partly, because of modulation of neurotrophic proteins involved in the regeneration processes.
Introduction
Cerebellar damage elicits motor disorders in muscle coordination, balance, and strength. 1 Most of these symptoms gradually and efficiently compensate over time in both humans and animal models following surgical ablation or stroke injury. Although the cellular mechanisms of the cerebellar compensation are not fully elucidated, it is believed that recovery of motor function and maintenance of a compensated state are a result of synaptic and neurochemical modifications in cerebellar and other brain regions. 2-5
A number of studies have shown the importance of environmental enrichment (EE) in maintaining motor and cognitive performance even in the presence of brain pathology. 6-8 The underlying molecular mechanism of EE-induced neuroprotection has been partially attributed to the upregulation of the levels of neurotrophins, proteins involved in both neuronal survival and death, and in activity-dependent plasticity. 9-11 Studies conducted in rodents have convincingly demonstrated that exposure to an enriched environment increases the production of neurotrophins, such as nerve growth factor (NGF) and brain-derived neurotrophic factor (BDNF), in the cerebral cortex and hippocampal formation. 12-17 In a previous article, 18 we demonstrated that exposure to an enriched environment is also able to modulate the expression of NGF and BDNF in the cerebellum. Neurotrophins in the cerebellum have a role in neuronal regulation during brain development 19 and in neuroprotection 20-22 from glutamate-induced neurotoxicity. To date, no study has addressed the question on whether the prolonged exposure to complex environments is able to modify the recovery from deficits elicited by surgical or traumatic ablations of cerebellar regions. On such a basis, it is reasonable to advance the possibility that EE may have beneficial effects on recovery of motor function after cerebellar damage, and these effects might be associated with changes in local neurotrophin production. To test this hypothesis we performed a hemicerebellectomy (HCb) in rats reared in an enriched environment or in standard conditions and evaluated the recovery of motor function as well as local neurotrophin production in the spared hemicerebellum and in extracerebellar brain regions.
Materials and Methods
Animals
Adult male Wistar rats (250-300 g; Harlan, Italy) were used in the study. The animals were kept under standard conditions with food and water ad libitum on a 12/12-hour dark/light cycle. Rats were assigned to 3 experimental groups: enriched-reared hemicerebellectomized rats (EH group), standard-reared hemicerebellectomized rats (SH group), and standard-reared control rats (SC group). The animals were maintained according to the guidelines for ethical conduct developed by the European Communities Council Directive of November 24, 1986 (86/609/EEC). All efforts were made to minimize pain or discomfort of the animals.
Rearing Conditions
On the 21st postnatal day, an even number of male littermates of the same dam were randomly assigned to enriched or standard rearing conditions. The enriched rats were housed in groups of 10 in a large cage (100 × 50 × 80 cm3), with an extra level constructed of galvanized wire mesh and connected by ramps to create 2 interconnected levels. The cage contained wood shavings, a running wheel, a shelter, colored plastic toys, and small objects. Throughout the enrichment period, the shelter and running wheel were kept in the cage, but the toys and constructions were changed twice a week. Once a week, the feeding boxes and water bottles were moved to different areas of the cage to encourage foraging and explorative behaviors. Furthermore, each enriched animal was handled daily for at least 10 minutes. The rats reared in standard conditions were pair housed in a standard cage (42 × 26 × 18 cm3) containing wood shavings but no objects. Feeding boxes and water bottles were kept in the same position. These animals received the usual care provided by the animal facilities staff but no particular or prolonged manipulation.
Surgery
On the 75th postnatal day, enriched- and standard-reared rats received a right HCb, as shown in Figure 1. Rats were anesthetized with Zoletil 100 (tiletamine and zolazepam: 50 mg/kg intraperitoneally; Virbac SRL, Milan, Italy) and Rompun (xylazine: 10 mg/kg intraperitoneally; Bayer SPA, Milan, Italy). A craniotomy was performed over the right hemicerebellum. The dura was excised, and the right cerebellar hemisphere and hemivermis as well as the fastigial, interpositus, and dentate cerebellar nuclei of the right side were ablated by suction (Figure 1). Care was taken not to lesion the extracerebellar structures. Animals belonging to the sham surgery group (controls) were anesthetized to perform the craniotomy over the cerebellar structures, but neither excision of meningeal membranes nor cerebellar ablation was performed. The wound edges were then sutured, and the animals were allowed to recover from anesthesia and surgical stress. They were maintained in their respective housing conditions for the entire testing period.
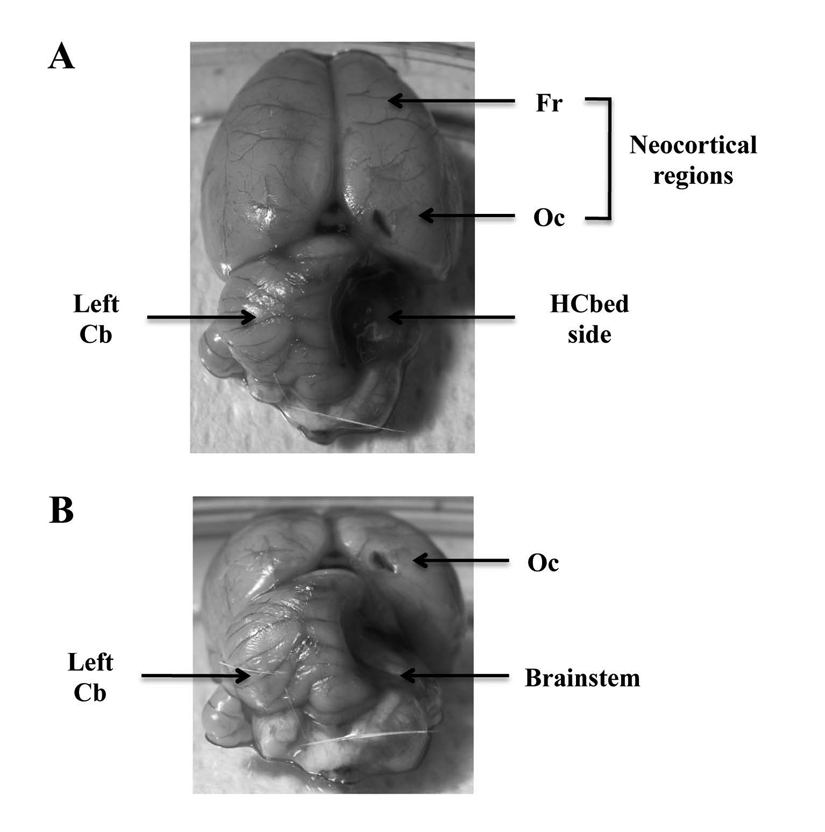
Representative macrophotographs illustrating the ablation of the right hemicerebellum. (A) Dorsal view of the brain. (B) Coronal view of the brain. Note the absence of the right hemicerebellum, the visibility of the right brainstem and the fourth ventricle and the sparing of the left hemicerebellum and of the extracerebellar structures
Postural Evaluation
Three hemicerebellectomized rats died during surgery or during the first days of postoperative recovery. Furthermore, data obtained from 1 lesioned animal that completed the behavioral testing were discarded because lesion verification indicated lesion incorrectness. Thus, the EH and SH groups comprised 8 lesioned animals each. To equalize the number of animals in the 3 groups, the SC group was constituted of 8 sham-lesioned rats.
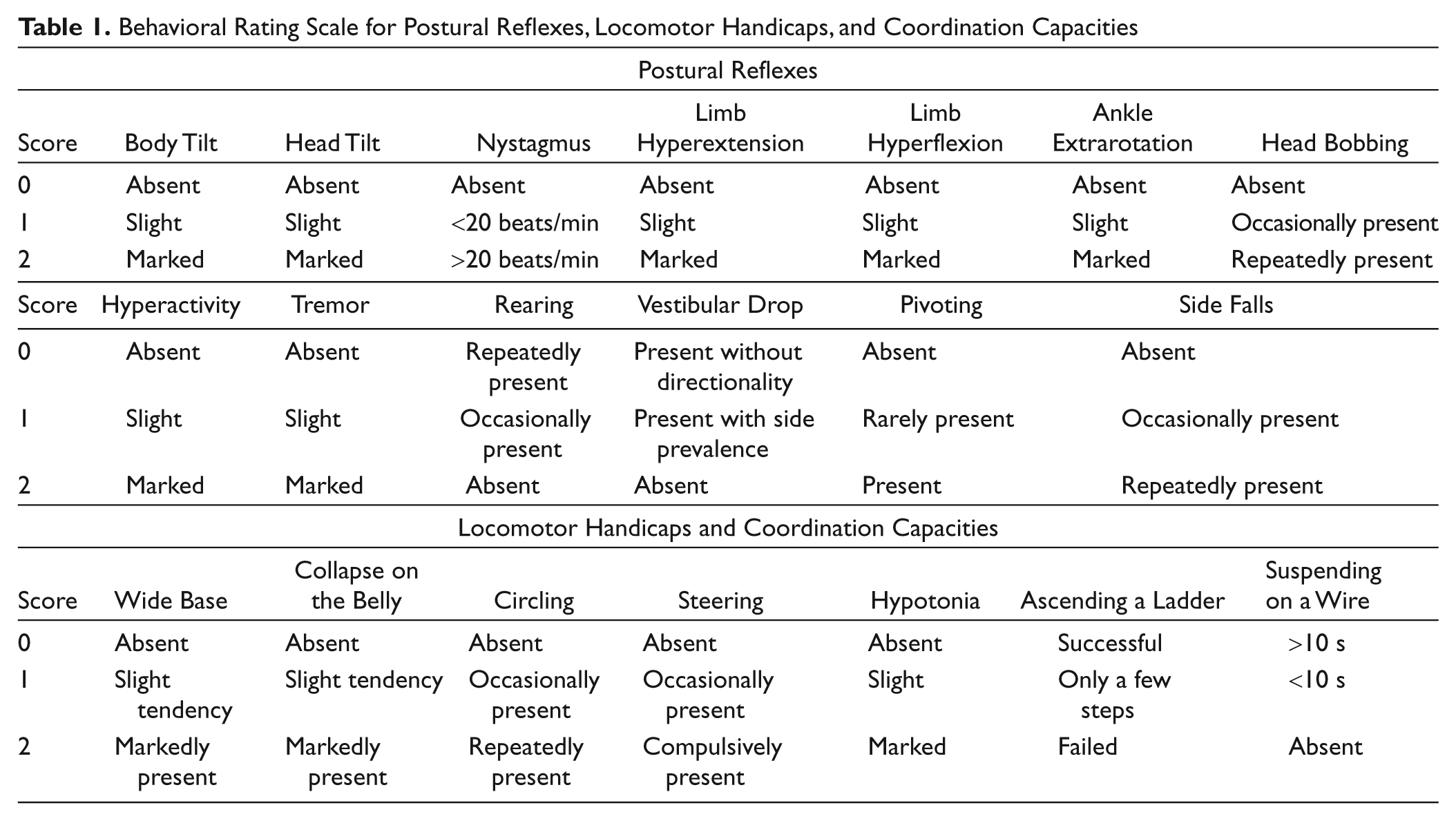
Postural evaluation started at 24 hours after HCb, and it was performed at variable time intervals till the 42nd postoperative day. Postural asymmetries and motor behavior were assessed by means of a behavioral rating scale described elsewhere in detail. 23,24 Postural evaluations were scored by an examiner unaware of rearing conditions the animals were submitted to. Presence or absence of head and body tilt, hyperflexion or hyperextension of forelimbs and hindlimbs in relation to the trunk, ankle extrarotation, hypotonia, eye nystagmus, head oscillations (bobbing), and tremor were evaluated. Some characteristics of locomotion—namely, wide-based, collapsing on the belly, steering, circling, pivoting, and falling to the side—were also analyzed. Specific postural and motor abilities, such as the vestibular drop reaction and rearing behavior, were also examined. Finally, complex motor skills, such as ascending a ladder and suspension on a wire, were assessed. Video records were taken throughout the entire testing cycle and were used to supplement direct behavioral observations. A score from 0 to 2 was assigned to each symptom according to its degree of severity, as described in Table 1. As 20 behaviors were taken into account, the total score ranged from 0 (complete absence of postural and motor deficits) to 40 (presence of all postural and motor symptoms to the highest degree).
Behavioral Rating Scale for Postural Reflexes, Locomotor Handicaps, and Coordination Capacities
Tissue Dissection for Neurotrophin Determination
At the end of the postural evaluation (5 months of age), 5 of the 8 animals in each group were randomly selected for neurotrophin determination. The animals were decapitated (starting at 10.00
NGF and BDNF Determination by ELISA
Concentrations of BDNF and NGF proteins were assessed using a 2-site enzyme immunoassay kit (Promega, USA). Briefly, 96-well immunoplates (Nunc A/S, Roskilde, Denmark) were coated (50 µL/well) with the corresponding captured antibody binding the neurotrophin of interest, overnight at 4°C. The next day, serial dilutions of known amounts of NGF and BDNF, ranging from 0 to 500 pg/mL, were performed in duplicate to generate the standard curve. Then, the plates were washed 3 times with wash buffer, and the standard curves and supernatants of brain tissue homogenates were incubated in the coated wells (100 µL each) for 2 h at room temperature (RT) with shaking. After additional washes, the antigen was incubated with the second specific antibody overnight at 4°C (NGF) or for 2 h at RT (BDNF), as specified by the protocol. The plates were washed again with wash buffer and then incubated with an anti-IgY horseradish peroxidase (HRP) for 1 h at RT. After another wash, the plates were incubated with a tetramethylbenzidine (TMB)/peroxidase substrate solution for 15 minutes and phosphoric acid 1M (100 µL/well) was added to the wells. The colorimetric reaction product was measured at 450 nm using a microplate reader (Dynatech MR 5000, Germany). Neurotrophin concentrations were determined from the regression line for the neurotrophin standard (ranging from 7.8 to 500 pg/mL purified mouse NGF or BDNF) incubated under similar conditions in each assay. For each assay kit, the cross-reactivity with other related neurotrophins was less than 3%. Neurotrophin concentration was expressed as pg/g wet weight, and all assays were performed in triplicate.
Statistical Analysis
The data were first tested for normality and homoscedasticity. Then, they were analyzed by 1-way or 2-way analyses of variance (ANOVAs) for independent (group) and repeated measures (day) followed by Tukey’s Honestly Significant Difference test. The significance level was set at P ≤ .05.
Results
Postural Evaluation
Dramatic rearing condition effects were observed in the postural and locomotor behaviors of the 2 groups of lesioned animals (Figure 2). Immediately after the cerebellar ablation, both SH and EH groups exhibited postural impairment of the same severity. From the second postoperative day onward, the EH group exhibited significantly less impaired posture, locomotion, and coordination compared with the SH group. The time course of compensation of the postural and motor symptoms of the EH and SH groups together with the baseline of the postural evaluation of the SC group are depicted in Figure 2. All animals were evaluated by means of the rating scale described in Table 1. However, because this rating scale evaluated symptoms and asymmetries of cerebellar origin, SC animals always obtained a score of 0 (complete absence of deficits). This kind of evaluation prevented the inclusion of the scores obtained by the SC group in the ANOVA. Thus, a 2-way ANOVA (group × day) was performed on the cerebellar deficit scores attributed to the 2 groups of hemicerebellectomized animals. This analysis revealed significant group [F(1, 14) = 150.45; P = .000001] and day [F(13, 182) = 127.57; P = .000001] effects. The interaction was also significant [F(13, 182) = 18.35; P = .00001]. Post hoc comparisons showed that performances of the 2 hemicerebellectomized groups were significantly different from the 2nd to the 28th day. These findings provide evidence of the beneficial effects of rearing in an enriched environment on the postural and locomotor performances of hemicerebellectomized animals. No relation was noted between housing conditions and lesion size.
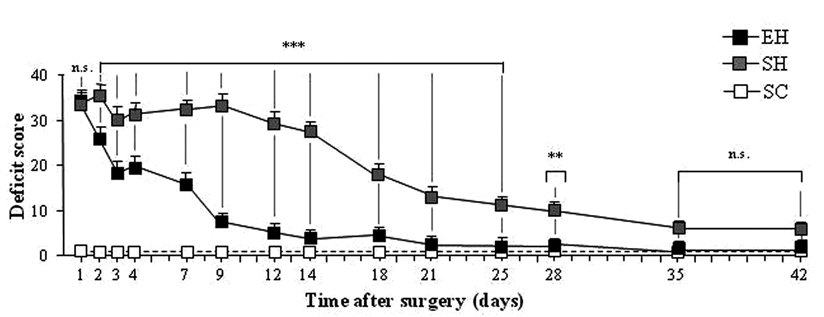
Postural and motor symptoms in enriched hemicerebellectomized (EH; n = 8) and standard-reared hemicerebellectomized (SH; n = 8) rats: the dashed line represents the baseline of postural evaluation of standard-reared controls (SCs; n = 8)a
NGF and BDNF Levels in the Cerebellum
A 1-way ANOVA on NGF levels in the cerebellum showed a significant group difference [F(2, 12) = 84.53; P < .00001]; see Figure 3. Post hoc comparisons showed that the EH group displayed significantly enhanced NGF levels compared with the SH (P < .0005) and SC (P < .0005) groups. Furthermore, the SH rats displayed increased NGF levels compared with the SC rats (P < .005). BDNF levels also differed significantly among groups [F(2, 12) = 10.88; P = .002]; see Figure 3. Post hoc comparisons revealed that although no differences were observed between EH and SH groups, both groups were characterized by elevated BDNF levels as compared with the SC group (P < .01 for both comparisons).
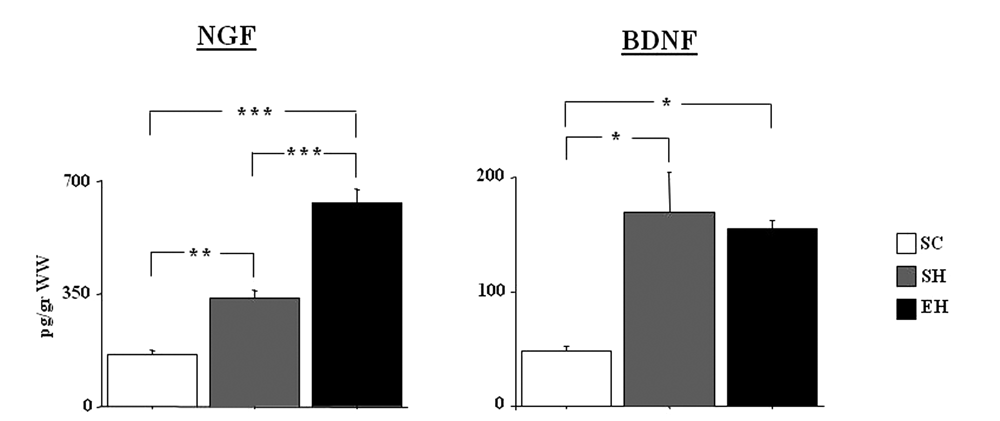
NGF and BDNF levels (pg/g) in the cerebellum of the 3 experimental groups (n = 5 rats/group)a
NGF and BDNF in Extracerebellar Brain Regions
In the hippocampus, there were no significant differences among groups for either NGF [F(2, 12) = 0.10; P not significant] or BDNF [F(2, 12) = 0.03; P not significant] levels. Conversely, in the frontal cortex, NGF levels differed significantly among groups [F(2, 12) = 8.31; P = .005]. Post hoc comparisons showed that the EH group had significantly higher levels in comparison with the SC and SH groups (P < .05 for both comparisons). Similarly, BDNF levels differed significantly among groups [F(2, 12) = 16.09; P < .0005]. Post hoc comparisons showed that the EH group had significantly higher levels in comparison with the SC and SH groups (P < .005 for both comparisons). In the striatum, no changes in NGF levels were observed among groups [F(2, 12) = 1.15; P not significant]. Conversely, a significant difference among groups was found when BDNF levels were analyzed [F(2, 12) = 15.85; P < .0005]. Post hoc comparisons showed that the EH rats had significantly lower BDNF levels than the SH (P < .0005) and SC (P < .05) rats (Figure 4).
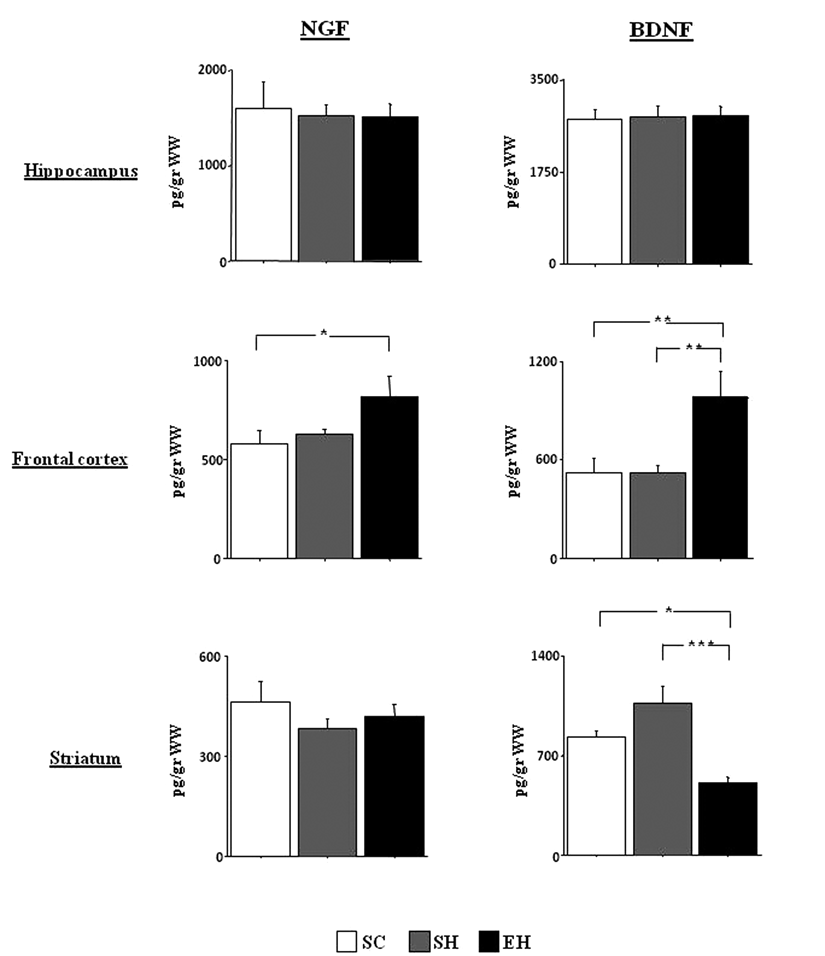
NGF and BDNF levels (pg/g) in the hippocampus, frontal cortex, and striatum of the 3 experimental groups (n = 5 rats/group)a
Discussion
A primary component of cerebellar motor symptomatology is the maximum degree of severity of the postural and locomotor impairment present immediately after the lesion, which decreases progressively as time goes by. EE represents a valid animal model to explore the effect of environmental complexity on brain resilience, the ability to cope with a neuronal damage while still functioning adequately. 26-28
In the present study, we analyzed the effects of a HCb in previously enriched rats. We found that long-term exposure to an enriched environment significantly accelerated the recovery of motor function from the deleterious effects of HCb. In addition, after the lesion, both NGF and BDNF were increased in the spared hemicerebellum. However, the enriched lesioned rats were characterized by a greater increase in NGF levels in the spared hemicerebellum as compared with nonenriched lesioned rats, whereas BDNF levels were not influenced by EE.
Thus, the present findings suggest that EE may accelerate the recovery processes from a cerebellar lesion and that neurotrophins are potentially involved in these plastic processes. This hypothesis is supported by the findings that in adult life, neurotrophins are able to regulate synaptic plasticity 29 and stimulate axonal regeneration. 30 Data obtained in other models of brain pathologies further support this notion. Exposure to an enriched environment ameliorated motor coordination deficits and partially normalized cerebellar BDNF expression in the female mice of a murine model of the Rett syndrome, a disorder characterized by deterioration of cognitive functions and motor coordination skills in early childhood. 31,32 Furthermore, research on the R6/1 transgenic mice model of Huntington’s disease demonstrated that exposure of Huntington’s disease mice to an enriched environment delayed the onset and progression of disease 33-36 and rescued the striatal deficit of BDNF. 35,37
In the cerebellum, both granule and Purkinje cells express NGF and BDNF and their receptors. 38-44 In a previous article, 18 we found that the same EE protocol used in the present research is able to increase the expression of both NGF and BDNF in the cerebellum of unlesioned rats, suggesting that these molecules are part of the mechanism of action of EE in this brain region. After the lesion, we found that although both neurotrophins were increased in the spared hemicerebellum, NGF cerebellar levels were further elevated in EE-enriched rats. The reason for these differences is not clear. It is known that in conditions of brain insult the 2 neurotrophins may differentially act on cerebellar neurons. For example, following a cortical trauma, inflammatory response triggered by cytokines induces an upregulation in NGF in the cortex and cerebellum. 45 NGF itself protects cerebellar Purkinje neurons from apoptosis induced by deprivation of trophic factors, 46 whereas cerebellar astrocytes in primary culture selectively respond to kainic acid and inflammatory cytokine exposure by increasing NGF but not BDNF. 47 Moreover, in developing cerebellar neurons, only NGF is able to induce a rapid release of glutamate and an increase of Ca2+ through a p75-dependent pathway. 48 Overall, these data indicate that NGF expression might be more important during development or after a brain insult, a hypothesis confirmed by the fact that the expression pattern of neurotrophins changes with age, with falling NGF but increasing granule cell BDNF. 19 Thus, one speculative explanation is that the faster motor recovery from a cerebellar lesion in enriched rats may be associated with a greater increase in NGF.
In addition, the present findings indicate that the changes in neurotrophin levels in other brain regions may participate in the recovery of motor function following a brain insult, such as HCb. Specifically, in the frontal cortex, NGF and BDNF levels were upregulated in EH rats as compared with SH and SC rats, in line with previous data obtained in enriched nonlesioned rats. 18,49 It is known that wide anatomofunctional connections 50 support the interplay between the cerebellar and frontal areas that interact in planning actions. The former permit acquisition of efficient procedural competencies, and the latter provide flexibility among already acquired and stored solutions. 51-53 Thus, it is possible that increased levels of neurotrophins in the frontal cortex may account for the improvement of motor performances that we observed in enriched lesioned animals.
We also observed a significant reduction of striatal BDNF levels in the EH rats. Striatal dopaminergic neurons utilize BDNF for their normal survival and function. It is well known that EE affects brain functions controlled by dopaminergic circuits, as the spontaneous and novelty-linked locomotor activity, 54 and normalizes the locomotor hyperactivity induced by administration of dopamine agonists. 55 Thus, it is possible that the reduced striatal BDNF synthesis of EH animals is associated with a reduced activity of dopaminergic striatal neurons. Alternatively, given that activity of striatal dopaminergic neurons is reported to be regulated by cerebellar 56-58 and prefrontal 59 regions, a speculative explanation advances that the reduced striatal BDNF levels might be a compensatory effect for increased BDNF production in the cerebellar and/or frontal cortex. Such an interpretation is in full agreement with a recent report in which we demonstrated that the striatal neuronal activity is markedly influenced by a cerebellar lesion and that synaptic changes occurring in the basal ganglia drive the compensation of motor deficits of cerebellar origin. 24
Conclusions
A long clinical and experimental history, going back to Lashley, 60 has examined the effects of preoperative experience on recovery from brain injury and the so-called brain reserve hypothesis produced by cognitive enrichment. 61 The present data emphasizing the positive effects of early and prolonged exposure to an enriched environment on HCb symptoms support the reserve hypothesis. A description of the neurochemical background linked to the modulation of neurotrophins underlying the protective effect of experience allows for better comprehension of this phenomenon and provides useful suggestions regarding efficacious lifestyles and rehabilitation strategies for coping with brain injury.
Footnotes
The author(s) declared no potential conflicts of interest with respect to the authorship and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: This work was supported by MIUR grants to Laura Petrosini.