
Abstract
The complexity and diversity of many human diseases pose significant hurdles to the development of novel therapeutics. New scientific and technological advances, such as pharmacogenetics, provide valuable frameworks for understanding genetic predisposition to disease and tools for diagnosis and drug development. However, another framework is emerging based on recent scientific advances, one we suggest to call pharmacoempirics. Pharmacoempirics takes advantage of merging two nascent fields: first, the generation of induced pluripotent stem cells, which are differentiated into mature cell types and represent patient-specific genetic backgrounds, and, second, bioengineering advances allowing sophisticated re-creation of human pathophysiology in laboratory settings. The combination of these two innovative technologies should allow new experimentation on disease biology and drug discovery, efficacy, and toxicology unencumbered by hypothesis generation and testing. In this review, we discuss the challenges and promises of this exciting new type of discovery platform and outline its implementation for cardiovascular drug discovery.
Introduction
There has been much debate about the relative decline in recent years of the rate of approved new therapeutic entities. Several eloquent reviews, describing the pharmaceutical discovery, development, clinical trials, and approval aspects of this trend, suggest potential explanations for this decline in technological productivity, which appears to be unique to the pharmaceutical industry.1–3 What is perhaps more troubling than the simple number of approved new molecular entities is the relative number of first-in-class drugs, or entities with novel molecular mechanisms of action and generally with novel biochemical targets. In a fundamental way, the therapeutic target represents the scientific community’s idea about how to intervene in a particular disease process, granted that there are numerous restrictions on targets such as specificity and druggability. Data analyzed by Swinney and Anthony 1 in 2011 illustrate that over the decade beginning in 1999, not only had the number of approved therapeutics per year significantly declined, but also the number of first-in-class therapeutics. More strikingly, of the 75 first-in-class Food and Drug Administration–approved new molecular entities during this decade, only 5 are designated as therapies for cardiovascular disease, the most prevalent disease class and the most frequent cause of mortality in the United States and other parts of the world. 4 An analysis by Pammolli and colleagues 3 indicates that the number of pharmaceutical industrial projects focused on cardiovascular disease has dropped in the past quarter century. For such a complex landscape as cardiovascular drug discovery and development, there are likely multifactorial phenomena contributing to the relative rarity of first-in-class cardiovascular therapeutics. These include pharmaceutical industrial and business strategy, perceived market needs, regulatory requirements, probable need for chronic treatment, complex genetics, insufficient understanding of cardiovascular pathophysiology, and lack of specific clinical biomarkers. However, recent focus has also been placed on the concept of biological reductionism. Reductionism has played a fundamental role in hypothesis-driven biological sciences and in the drug discovery process; however, there has been discussion in the scientific community about whether such reductionist thinking has been overemployed to the detriment of drug and scientific discovery.5–8 Hence, here we propose a novel strategy for experimentation applicable to cardiovascular drug discovery and basic research on disease biology, with the intent to peel back some layers of reduction in disease modeling experiments and compound screening.
Motivation for Pharmacoempirics as a Drug Discovery Strategy
The most relevant model system in which to study human disease biology and evaluate therapeutic effects is of course the living human organism. Importantly, ethical and practical limitations preclude such human research beyond appropriately regulated clinical trials. Thus, scientists must introduce varying degrees of reductionism and surrogacy into disease-related research with the hope of maintaining faithful reproduction of human physiology and pathology amid the ever present risk of introducing irrelevant experimental effects. The persistent and in fact growing 3 attrition of experimental therapeutics during all stages of clinical trials and following approval due to either insufficient efficacy or intolerable patient safety suggests that species-specific and experiment-related effects of reductionist experimental approaches are real and very costly. This margin of failure should prompt the academic and pharmaceutical communities to scrupulously ask to what extent is reduction appropriate and to what extent is it excessive. The answer to this, no doubt, is complicated and certainly dependent on which processes or diseases one is interested. It is for these reasons that we wish to highlight a strategy for modeling human disease amenable for drug discovery with the hope of limiting the layers of biological reduction often imposed. Pharmacoempirics is the use of differentiated cells derived from stem cells of human individuals combined with functional phenotypic readouts for the purpose of drug discovery. This in vitro drug discovery strategy seeks to find new targets and therapeutics by integrating three elements with traditional compound screening: (1) cellular models with disease-relevant human genetic background, (2) re-creation of the cellular microenvironment of the target cells, and (3) evaluation of cellular function as the primary screen readout.
Specifically, a relevant human disease model uses cells of human origin, of the appropriate cell type, and with the identical genetic background as the disease-relevant cells within the human organism. Enough significant differences exist between human and rodent or other animal physiology, cell biology, toxicology, and genetics to warrant substantial caution in employing nonhuman model systems in drug discovery efforts. The use of cell types other than the type relevant and implicated in a particular disease may obscure the molecular network associated with a drug target that would be affected by the drug within the relevant cell type.6,9 The precise genetic background of patients affected by cardiovascular disease is essential as it is believed that in some cases, multiple complex genetic traits, some discovered and some undiscovered, can influence cardiovascular disease risk in a convoluted fashion.10,11
Second, years of research across disciplines have indicated that cellular phenotype is dependent in nontrivial ways on the rich details of the cellular microenvironment, whether those are the biochemical effects of the extracellular matrix,12,13 the mechanical properties of the substrate,14–16 complex biomechanical forces,17,18 growth factor gradients,19,20 or interactions with other cell types.21–23 Due to this interdependency between the microenvironment and cellular phenotype, incorporating the most salient aspects of the microenvironment may expose different and more physiologically relevant biochemical pathways, potentially altering the outcome of in vitro experiments and the effects of experimental therapeutics.
Last, a pharmacoempiric approach to drug discovery incorporates elements of empiricism or, in applied biological sciences, phenotypic observation as the output of experiments and screens. As a number of cogent studies have recently elaborated,1,24–26 phenotypic as opposed to target-based screening bears several benefits, such as the potential to affect multiple biochemical targets and eliminate bias regarding particular pathways. Phenotypic screening is an effective approach in the discovery of first-in-class therapeutics, recently producing more small-molecule therapeutics than target-based approaches. Furthermore, experimentation with a functional rather than a biochemical phenotypic output will likely have greater relevance. Perhaps at its most fundamental, the success of phenotype-based approaches is an admission of humility regarding the scientific community’s inability to prospectively identify specific potent targets for therapeutics able to alter the course of human disease progression.
We propose that by incorporating these three elements—relevant human disease models, realistic human microenvironments, and systematic observation and probing of cellular function—pharmacoempirics liberates experiments and drug discovery screens from several layers of reductionism with the ultimate goal of discovering new therapeutics and improving clinical trial attrition ( Fig. 1 ). This strategy is ideal for pathologies with poorly understood molecular etiologies and rare genetic diseases but certainly can be useful more broadly. As will be described, because of recent inspiring advances within the past decade in human pluripotent stem cell biology, in conjunction with clever, precise biomedical engineering, such pharmacoempiric experiments may be possible and even feasible within the scales relevant to the pharmaceutical industry.
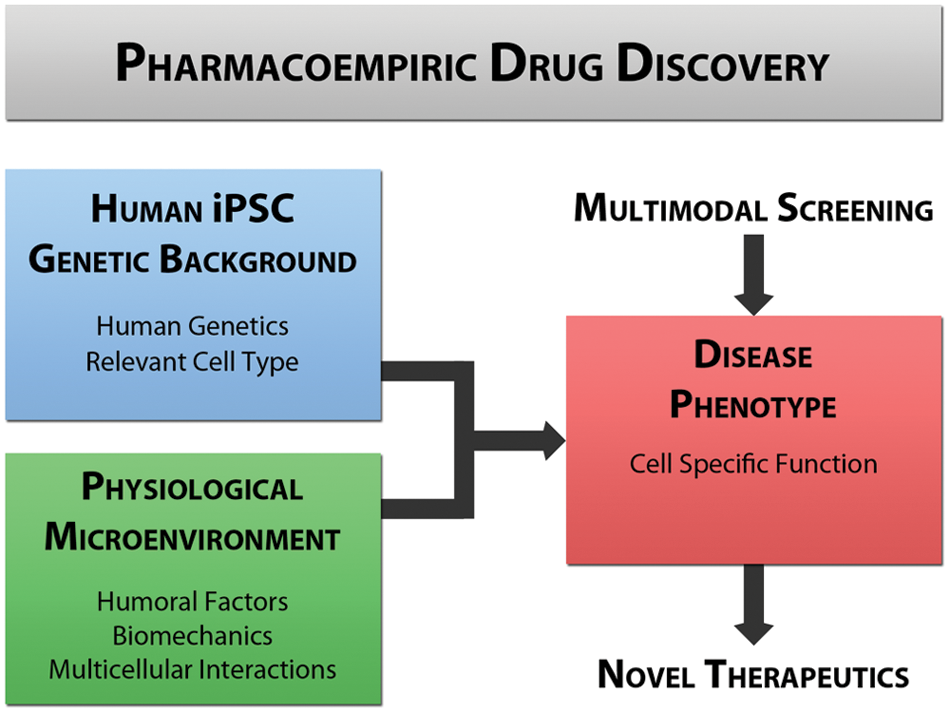
Outline of the pharmacoempiric approach to drug discovery and disease research intended to limit the reductionism in biological experimentation. iPSC, induced pluripotent stem cell.
Human Disease Models with Induced Pluripotent Stem Cells
A central element of pharmacoempiric experimentation is the incorporation of relevant human cell types bearing particular genetic backgrounds. Traditionally, this dimension has been problematic as most human cell types are not easily procured but derived from cell lines or harvested from scarce discarded pathology tissue or cadaveric donors. Cell line behavior may deviate from their parental phenotype in significant ways, whereas primary cells have limited expansion potential in vitro before phenotypic drift. In particular, acquiring a robust and reproducible source of vascular cells, such as vascular endothelium, smooth muscle, or cardiac myocytes, is a significant challenge for drug discovery. This availability of vascular tissue from patients with specific genetic diseases is even more problematic.
The discovery of cellular reprograming as a technology to generate induced pluripotent stem cells (iPSCs) offers a potential solution to the challenge of limited cardiovascular cell sources. In this approach, human adult somatic cells are dedifferentiated into stem cells offering comparable function to human embryonic stem cells in their ability to develop differentiated progeny from all developmental lineages in the organism. This technology enables the scientist to choose the particular patient source and corresponding genetic background for further study or screening. This patient sample is induced to pluripotency and subsequently directed to differentiate into the desired cell type. Significantly, these human iPSCs are infinitely expandable in their pluripotent form and lack the ethical concerns surrounding manipulation of the human embryo as in embryonic stem cell derivation.
Human-induced pluripotent stem cells were originally derived by retroviral overexpression of four transcription factors—Oct4, Sox2, Klf4, and c-myc—observed to be present and essential in human embryonic stem cells. 27 However, since this seminal discovery, numerous alternative strategies for derivation have been accomplished, some involving integration of exogenous factors into the genome and some without any genome modification, as reviewed elsewhere. 28 The methodology to achieve reprogramming of adult human somatic cells to pluripotency will no doubt continue to be a subject of great academic and commercial interest with efficiency, speed, and reproducibility to improve.
To be useful for modeling cardiovascular disease and participating in the drug discovery process, iPSCs need to be differentiated into cell types relevant for various cardiovascular diseases—namely, endothelium, smooth muscle, cardiac myocytes, fibroblasts, and hematopoietic lineage cell types known to participate in vascular disease pathogenesis such as monocytes and lymphocytes. There has been some progress in generating such cardiovascular disease–relevant cell types. In general, this was accomplished either through the formation of differentiating heterogeneous 3D embryoid bodies from which a cell type may be isolated, as exemplified within Figure 2 , or though addition of various factors to encourage pluripotent colonies to differentiate along a specific lineage. In particular, human iPSCs within various in vitro contexts have been differentiated into endothelium,29–32 smooth muscle cells,30,33–35 monocytes, 36 and neutrophils.37–39 Despite these reports, which describe the emergence of cell populations largely bearing molecular markers of the desired cell type, there is a paucity of functional characterization of the differentiated cell types. There are still significant unanswered questions about the character of the iPSC-derived vascular cells—namely, whether they faithfully represent the mature phenotype and can perform the many dynamic functions of adult vascular cell types offering a surrogate model system for human primary cells.
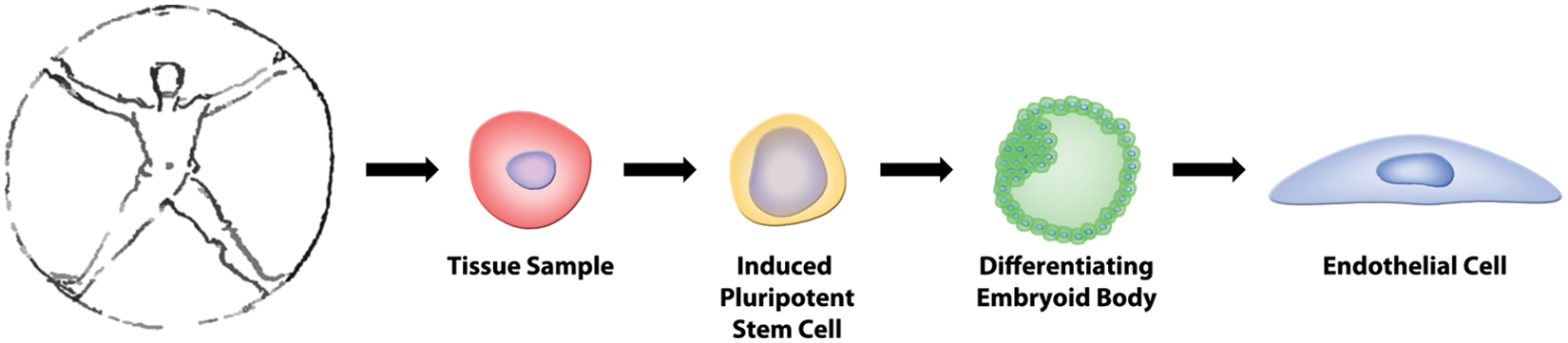
The process of how patient samples or biopsies can be reprogrammed to an induced pluripotent stem cell, brought to differentiate and to produce a particular cell type of interest.
Employing iPSCs to generate specific cell types to model a genetic disease presents a greater challenge. Here, one must create an iPSC line derived from tissue from patients presenting with a genetic disease of interest. These cells must be reprogrammed and differentiated into the cell type of interest. Although this must be theoretically possible as the living patient is evidence that the disease-causing genetic mutation does not preclude pluripotency and development, there may be challenges efficiently re-creating this process in vitro in the context of genetic abnormalities. Once a particular cell type with a given genetic mutation is produced, the mutation must be shown to be present in the cell type and further to have a functional consequence. There has been discussion as to the appropriate experimental control for such disease modeling, with the strongest candidate being iPSC lines genetically engineered to be isogenic to the disease cell line with the exception of a genetic alteration unraveling the original disease-causing genetic mutation. There has been some precedent for this genome editing approach with iPSC lines using zinc finger nuclease technology40,41 and with transcription activator–like effector nucleases. 42 These technologies create two iPSC lines genetically identical with the exception of a point mutation corrected, hence giving power to the specificity of a phenotype illustrated with the patient iPSC-derived cells. However, there are many situations where such an approach may be inappropriate or not possible. More complex genetic abnormalities such as trisomies, copy number variations, and other complex chromosomal disorders are not able to be corrected in this fashion. Moreover, one of the appealing strengths of iPSCs in general is that the particular genetic mutations responsible for an observed phenotype known to be hereditary need not even be identified to produce iPSC lines and hence differentiated progeny bearing that unidentified mutation for further study. In such cases, other approaches to create appropriate control comparisons include generating iPSC lines from unafflicted siblings or parents, which will bear genetic similarity to the disease model cell line. Alternatively, a strength-in-numbers approach is possible where iPSC lines are generated from many individuals both healthy and afflicted with a particular disorder with the hope of capturing and perhaps averaging across different genetic variation.
Thus far, there are limited examples of modeling human cardiovascular diseases with iPSCs. In particular, two groups have generated iPSC lines from patients with Hutchinson-Gilford progeria syndrome (HGPS), a genetic premature aging disorder that bears clinical resemblance to physiological aging. 43 Specifically, patients with HGPS develop advanced atherosclerosis, among other pathologies, at a young age that frequently gives rise to fatal myocardial infarctions and strokes. Previously, the study of cardiovascular disease within this rare disease population was restricted to studies on primary harvested dermal fibroblasts or histological study of cadaveric tissue. Both groups created HGPS iPSC lines and differentiated these cells into vascular smooth muscle cells to show modest increased senescence under several in vitro conditions, suggesting a possible mechanism for the premature vascular disease observed in this disorder.44,45
Human iPSCs offer tremendous promise as a source of human cardiovascular cells and especially as a cellular source from patients with rare genetic disorders, multiple complex genetic polymorphisms, or hereditary conditions of undefined genetic origin. In essence, iPSCs are an in vitro human analogue for some transgenic animal models. The most pressing need remaining to fully realize this new resource is for further involvement of cardiovascular cell biologists in this nascent field of stem cell biology to define the conditions to generate cell types relevant for cardiovascular disease bearing the complete biomolecular and functional repertoire as the adult human cell types.
Re-creation of the Vascular Microenvironment
The human vasculature is a dispersed complex organ composed of several cell types principally including endothelium, smooth muscle, and fibroblasts. The microenvironment of the vasculature is rich with biomechanical, heterotypic intercellular, and humoral stimulation, all of which can significantly influence vascular cellular phenotype. These interactions are often fundamental to many physiological and pathological processes, which also are the subject of disease modeling and therapeutic screening experiments. Complete construction and long-term maintenance of a functional human blood vessel in vitro have not been possible yet; however, numerous insights have been proposed to introduce complex physiological stimuli and structures present in the in vivo microenvironment into in vitro experimentation to more realistically model vascular cell function critical for cardiovascular disease. Here we highlight such efforts around the context of a single cell type, the vascular endothelial cell.
Biomechanical Forces
The vascular endothelium, the single cell layer lining the lumen of blood vessels, is a dynamic interface responsible for maintaining vascular homeostasis, participating in inflammatory responses, initiating thrombosis, regulating vasomotor tone, and mediating vascular permeability. Being the interface between flowing blood and the rest of the vessel wall, endothelial cells are exposed to complex dynamic biomechanical forces in the form of shear stress applied to their luminal surface, hydrostatic pressure, and strain as the vessel geometry changes during the cardiac cycle. There has been significant focus on endothelial mechanotransduction, the sensation and biological interpretation of biomechanical stimuli.18,46–49 To maintain vascular homeostasis, endothelial cells sense hemodynamic fluid shear stress and rapidly produce several mediators that act in an autocrine and paracrine manner within the vessel wall and beyond. Furthermore, endothelial cells react to different fluid shear stress waveforms by acquiring distinct functional phenotypes, including a proinflammatory atherosclerosis-susceptible or an atherosclerosis-resistant phenotype depending on the particular biomechanical stimulus.50–54 Multiple studies have documented that hemodynamic forces can influence endothelial cell cytoskeletal structure, cell migration, oxidative state, secretion of growth factors and cytokines, and monolayer permeability. 47 As these effects of fluid shear stress on the endothelium are fundamental to endothelial phenotype and function, it will be very insightful to study vascular disease biology and the effects of experimental compounds under physiologically relevant biomechanical conditions. Traditionally, this has been performed by placing cultured endothelial cells under fluid shear stress driven either within a motorized cone and stationary plate system, as originally described by Dewey et al, 55 or through soft lithography-based microfluidic devices.56,57 Although some commercial devices offer parallelization in 24-well format, 58 there is need for greater miniaturization, throughput, and, importantly, faithful re-creation of human biomechanical stimuli so that hemodynamic conditions can be incorporated into drug discovery screening applications. Influencing endothelial function and dysfunction with biomechanical stimuli may prove widely useful as endothelial dysfunction contributes to multiple cardiovascular diseases, 59 particularly coronary artery disease, stroke, peripheral vascular disease, and deep vein thrombosis, and leads to significant complications in diabetes mellitus 60 and chronic kidney disease, 61 systemic lupus erythematous, 62 Kawasaki disease, 63 several hereditary vascular malformations, 64 and pathological tumor angiogenesis. 65
Multicellular Systems
Larger and medium-size vessels contain vascular smooth muscle cells that respond to humoral, endothelial, and systemic nervous system signals to modulate their cellular contractility and hence vasomotor tone, blood pressure, and tissue perfusion. Endothelial and smooth muscle cell communication, accomplished through secretion of cytokines, growth factors, nitric oxide, and extracellular vesicle–delivered microRNAs, is able to regulate smooth muscle cell proliferation, contractility, and phenotype.66,67 Importantly, dysregulation of vascular smooth muscle cells is implicated in chronic hypertension, vascular calcification, and intimal hyperplasia found in atherosclerosis and settings of vascular injury.66,68 Within the microvasculature, capillary endothelium is in close association with pericytes known to be involved in stabilizing vessels during angiogenesis and regulating capillary perfusion and permeability.22,69,70 Dysfunction of the endothelial-pericyte interaction is implicated in disrupting the vasculature in tumor angiogenesis and within diabetic retinopathy. Furthermore, the multicellular microenvironment present in various organs sustains distinct organ-specific vascular phenotypes. This can be observed within the liver, where discontinuous fenestrated sinusoidal endothelium interacts with hepatocytes and fibroblasts, forming a specialized endothelial subtype that participates in transport to and from the circulation.71–74 Similarly, the glomerular endothelium of the kidney is another specialized fenestrated subtype that cooperates with podocytes and mesangial cells to maintain a functional selectively permeable filtration barrier.75,76 Analogously, interaction between cerebral capillary endothelium and astrocytes supports the blood-brain barrier, a specific and restrictive barrier characterized by paracellular tight junctions limiting permeability. 77 Due to the complexity of the interactions between endothelium and other cell types, as well as their implications on vascular function, there is motivation for multicellular culture systems that replicate these interactions and physiological processes within disease biology and drug discovery experiments focused on angiogenesis and complex vascular function.
Humoral Factors–Induced Endothelial Dysfunction
The vascular endothelium is incessantly in intimate contact with blood, providing oxygen, nutrition, and many humoral signals. However, within several pathological settings, the blood concentrates factors believed to have deleterious effects on the endothelium. Here, we cite two chronic disorders, chronic kidney disease and diabetes mellitus, where factors within the blood plasma negatively influence endothelial function through mechanisms that today remain poorly understood. The endothelial dysfunction frequently observed in these disorders very likely contributes to the heightened risk of adverse cardiovascular complications found within these patient populations. 78
Chronic kidney disease is a progressive deterioration of renal function and poor glomerular filtration of the blood, resulting in a toxic uremic milieu within the plasma. 61 This uremia can adversely affect the vasculature, resulting in endothelial dysfunction evident through impaired flow-mediated vasodilation and development of a proinflammatory phenotype in endothelial cells.79,80 Critically, the specific toxic elements responsible for endothelial dysfunction within the uremic milieu of patients with renal dysfunction have not been fully characterized.
Also, endothelial dysfunction and its consequence of impaired flow-mediated vasodilation are observed within patients with diabetes who display insulin resistance. 81 Research has identified several potential mechanisms through which chronic hyperglycemia may induce endothelial dysfunction, including participation of advanced glycation end products, creating oxidative stress and insulin signaling, although many of the pathological mechanisms remain unknown.82,83
Mitigating the endothelial dysfunction within these chronic disorders is likely to reduce the heightened risk of serious adverse cardiovascular events observed within these patient populations. There are no therapeutics currently available designed specifically to treat this endothelial dysfunction. This may be in part because the molecular etiologies of this endothelial dysfunction are unknown and because simple addition of defined factors to cultured endothelium cannot fully re-create these pathologic backgrounds.
Assessment of Cellular Function
The study of disease biology and drug discovery often employs one or several molecular biomarkers as experimental or screening readouts. As mentioned earlier, phenotypic screening has the advantage of interrogating compounds in a pathway-unbiased fashion, allowing compounds with multiple or unknown targets. Furthermore, phenotypic screening with integrated cellular functions as end points eliminates even the bias of the choice of biomarker. Fortunately, advances in cellular biology coupled with sophisticated biomedical engineering of tissue culture tools allow for real-time multidimensional quantitation of vascular endothelial function. Of the many dynamic functions performed by endothelial cells that are important in human disease, we describe here several with cell-based assays amenable to screening.
Angiogenesis
One of the fundamental functions of the vascular endothelium is to develop new blood vessels from existing ones. This process is controlled through local balance of pro- and antiangiogenic factors, pathways, and genes. There are several genetic disorders wherein mutations occurring within these angiogenic genes cause vascular malformations and disease. 84 Mutations within VEGFR2 and VEGFR3 genes have been linked to cases of infantile hemangioma, a vascular tumor. 85 Hereditary hemorrhagic telangiectasia is a disorder characterized by the presence of arteriovenous malformations, direct connections between arteries and veins eclipsing the capillary bed. In some instances, these malformations can lead to hypoxemia, stroke, heart failure, and fatal hemorrhage. Although mutations within genes involved in transforming growth factor (TGF)–β signaling pathways, such as SMAD4, 86 endoglin, 87 and ALK1, 88 have been identified in patients with hereditary hemorrhagic telangiectasia, the pathological mechanisms responsible for the malformations remain unclear, complicated by the intricate TGF-β signaling network. Arterial tortuosity syndrome, a disorder marked by geometric abnormalities of medium and large arteries, has been shown to be caused by mutation in a glucose transporter. 89 Mutations within several genes, CCM1–3, are associated with dilated capillaries and saccular vascular structures in the brain, termed cerebral cavernous malformations.90,91 There are numerous other examples of vascular malformations with genetic origins. 64 In all these instances, the creation of a human genetic model through induced pluripotent stem cells coupled with assessment of angiogenic potential may elucidate the functional consequences and molecular mechanisms behind these vascular malformations.
Development of therapeutics for promoting, inhibiting, or normalizing angiogenesis has leveraged knowledge regarding the various growth factors and associated receptors and signaling programs. However, the biology governing the endothelial decision to create new blood vessels, of a particular fate and size and at a particular anatomical location, is not fully understood. A nascent endothelial tube is dependent on the recruitment of vascular smooth muscle cells for arteries and veins and pericytes for capillary vasculature. For these reasons, it may be valuable to study disease biology or to evaluate the effects of experimental compounds employing in vitro angiogenesis as a functional phenotypic end point. There are multiple widely used protocols for assessing angiogenesis in vitro 92 and several schemes available for interpreting and quantifying visual high-content data for high-throughput applications performed in 96-, 384-, and 1536-well formats.93,94 The process of screening compounds that influence the angiogenic process can even be extended to in vivo screens with zebrafish in a 384-well format. 95
Leukocyte Interactions
In response to proinflammatory stimuli, the vascular endothelium acquires an activated phenotype characterized by the secretion of proinflammatory cytokines and chemoattractants and presentation of intercellular adhesion molecules to its surface.96–99 This coordinated response can promote the interaction of monocytes, neutrophils, and T cells and the vascular wall mediating many inflammatory processes. This leukocyte-endothelial interaction has significant consequences for the defense against pathogens, atherosclerosis,100,101 and autoimmune disorders such as systemic lupus erythematous, 102 Kawasaki disease, 103 and scleroderma. 104 Although many of the specific mediators, chemokines, and adhesion molecules have been identified, the signaling mechanisms responsible for these interactions are not fully understood, especially in the context of cardiovascular, autoimmune, and rare diseases. The interaction of leukocytes and endothelium has been studied in vitro under a variety of inflammatory contexts. As mentioned earlier, the local hemodynamic environment carries significant influence on the inflammatory potential of the vascular endothelium. In addition, the biophysical interactions of the leukocyte—rolling, slow rolling, arrest, and transmigration—occur amid specific hemodynamic conditions in the vasculature that can be recreated with in vitro flow systems.105–107 Here, the kinetics of rolling and the magnitude of adhesion within leukocyte-endothelial interactions can be assessed. Although traditionally, data are viewed by video, this data analysis can be automated and analyzed as the output of a compound screen. 108 Systems to measure adhesion of leukocytes to endothelium in the absence of flow are currently commercially available in a 96-well format.
Vascular Permeability
An essential function of the vascular endothelium is to maintain a dynamic and selectively permeable barrier regulating the transport of fluid and macromolecules between the circulation and the interstitial space. This transport is physically accomplished by sophisticated machinery mediating both transendothelial and paraendothelial pathways. The dynamism of this barrier function is regulated by a variety of soluble factors, cell surface receptors, signaling molecules, and cytoskeletal and other structural proteins.109,110 Vascular endothelial permeability is critical for maintaining vascular homeostasis and for access of pharmacological agents to cross the specialized blood-brain barrier. Furthermore, endothelial permeability contributes to inflammatory processes 111 and dysfunctional vasculature near solid tumors 112 as well as in chronic disease states. In particular, the advanced glycation end products present in diabetic vasculature have been shown to influence endothelial barrier function, resulting in vascular hyperpermeability113,114 with several molecular mediators identified. 115 The complex effects of these pathological settings on endothelial permeability can be assayed in vitro most commonly either through detection of transcellular transport of fluorescently labeled macromolecules such as dextran through dual-chambered culture devices or by measurement of electrical impedance between microelectrodes across endothelial monolayers.116,117 These assays are highly quantitative and are able to produce time-dependent permeability data able to discern transient effects. Miniaturization and parallelization of these assays, both fluorescent-based macromolecule transport 118 and commercially available 96-well platforms and culture plates for measuring transendothelial electrical resistance, permit screening assays to use endothelial permeability as a direct functional output.
Future
It may be that pharmacoempirics, by employing human cells with appropriate genetic background of relevant cell type, re-creating precise physiological environments, and using cell-based assays of cellular function, is able to more faithfully reproduce pathophysiological processes in vitro, offering a more relevant human experimental platform for drug discovery. Here we mentioned several aspects of the vascular microenvironment and vascular endothelial function that biotechnology has been able to mimic and miniaturize for at least medium-throughput experiments, which may prove useful for the discovery of new cardiovascular therapies and drug targets. The technology exists to further increase parallelization should industrial rather than academic laboratories become more interested in their use. This pharmacoempiric approach is not limited to vascular biology but is more widely applicable since the complexity of the cellular microenvironment and cell function are intertwined in many physiological settings. This may be of particular importance in research on orphan and rare diseases in which little is known about the molecular basis of the disease and where primary tissue is unavailable for study. Performing this empiricism-based drug discovery does not minimize the insightful and tremendously useful understanding of the specific biological mechanisms that are responsible for this interplay between genetics and pathophysiology that pharmacoempirics explores from a naive perspective. But pharmacoempirics is simply an admission that focusing solely on individual genes, proteins, or even pathways may not be the best strategy for drug discovery.
Footnotes
Acknowledgements
The authors wish to acknowledge productive scientific discussions with Dr Julien Pham, Jennifer Cloutier, Pranati Kuchimanchi, and Choah Kim.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
William J. Adams is a recipient of a Ruth L. Kirschstein National Research Service Award from the National Institutes of Health (F31 AG037249).