
Abstract
Background:
The development of thoracic aortic aneurysm and dissection (TAAD) is mediated by inflammasome activation, which exacerbates the secretion of pro-inflammatory cytokines, chemokines, matrix metalloproteinases (MMPs), and reactive oxygen species (ROS). The glycolytic enzyme pyruvate kinase M2 (PKM2) has shown a protective role against various disorders with an inflammatory basis, such as sepsis, tumorigenesis, and diabetic nephropathy. However, its potential role in TAAD has not been investigated so far.
Approach and Results:
We analyzed aortic tissues from TAAD patients and the β-aminopropionitrile fumarate (BAPN)–induced mouse model of TAAD and observed elevated levels of PKM2 in the aortic lesions of both. Treatment with the PKM2 activator TEPP-46 markedly attenuated the progression of TAAD in the mouse model as demonstrated by decreased morbidity and luminal diameter of the aorta. In addition, the thoracic aortas of the BAPN-induced mice showed reduced monocytes and macrophages infiltration and lower levels of IL-1β, MMPs, and ROS when treated with TEPP-46. Furthermore, TEPP-46 treatment also suppressed the activation of the NOD-like receptor (NLR) family and pyrin domain–containing protein 3 (NLRP3) inflammasome by downregulating p-STAT3 and HIF1-α.
Conclusion:
Pyruvate kinase M2 plays a protective role in TAAD development, and its activation is a promising therapeutic strategy against the progression of TAAD.
Keywords
Introduction
Thoracic aortic aneurysm and dissection (TAAD) is a lethal cardiovascular condition with a high mortality rate. 1 The aortal wall of the TAAD patients shows loss of smooth muscle cells (SMCs), infiltration of inflammatory cells, degraded extracellular matrix (ECM), and fractionated elastic laminae. 2 -4 These changes progressively dilate and weaken the aorta wall and tear the intima, which lead to aortic aneurysm and dissection as well as arterial rupture. 5 -8 Although surgical interventions such as thoracotomy or endovascular repair are established therapies for TAAD, noninvasive therapies that reduce the risk of aneurysm are also needed.
Inflammation is a critical factor in development of TAAD, and patients with type A dissection show increased infiltration of pro-inflammatory cells such as macrophages, T cells, and neutrophils. 9,10 The macrophages and monocytes secrete pro-inflammatory cytokines, matrix metalloproteinases (MMPs), and reactive oxygen species (ROS), which deplete the elastic fibers and degrade the ECM, resulting in increased collagen turnover. 11 -13 In a previous study, we found that injecting Smad3−/− mice with the neutralizing antibody against granulocyte-macrophage colony stimulating factor significantly alleviated the inflammatory reaction and thoracic aorta dilation. 14 In addition, compared to healthy individuals, TAAD patients show increased levels of IL-1β, the primary effector of the inflammatory cascade, in the aortic wall and serum. 12,15,16 Inflammasome is a large multimolecular cytosolic complex that mediates the activation of caspase 1 and converts the pro-IL-1β and pro-IL-18 precursors into their biologically active forms. 17,18 Nucleotide-binding oligomerization domain-like receptor (NLR) family pyrin domain–containing 3 (NLRP3) is the best characterized inflammasome that can be activated by diverse stimuli such as inflammation, K+ efflux, Ca2+ signaling, mitochondrial dysfunction, ROS, and lysosomal rupture. 19,20
Recent studies have implicated a pathogenic role of metabolism reprogramming, especially that of glucose, in cardiovascular diseases. 21,22 Furthermore, inflammatory disorders such as lipopolysaccharide (LPS)-induced acute lung injury and sepsis are also mediated by glucose reprogramming, and hexokinase, the enzyme catalyzing the first step of glycolysis, is known to regulate NLRP3 activation. 23 -26 Aerobic glycolysis or “Warburg effect” is a metabolic reprogramming process common to cancer cells, wherein glycolysis rate increases even during aerobic conditions, while mitochondrial oxidative phosphorylation is reduced. 27 Pyruvate is the common substrate of both pathways, and the M2 isoform of pyruvate kinase (PKM2) catalyzes the rate-limiting step of phosphoenolpyruvate conversion into pyruvate. The secretion of interleukin 1β (IL-1β) is closely related to PKM2-dependent aerobic glycolysis in LPS-stimulated macrophages. 25,28
We hypothesized that PKM2-dependent glycolysis regulates IL-1β secretion via the activation of inflammasome in TAAD. Lysyl oxidase (LOX) is an enzyme responsible for the cross-linking of collagen and elastin both in vitro and in vivo. Lysyl oxidase (LOX) and related LOX-like (LOXL) isoforms play a vital role in remodeling the extracellular matrix (ECM). 29 The LOX family controls ECM formation by cross-linking collagen and elastin chains. 30 Beta-aminopropionitrile (BAPN) is a potent irreversible inhibitor of lysyl oxidase. 31 To this end, we established a mouse model of TAAD by oral administration of BAPN and concurrently treated them with the PKM2 activator TEPP-46. Pyruvate kinase M2 was upregulated in the TAAD lesions of both murine and human aorta samples, and its pharmacological activation suppressed NLRP3-mediated IL-1β secretion and TAAD progression. In conclusion, PKM2 activation is a promising therapeutic strategy for preventing and treating TAAD.
Materials and Methods
Establishment of the TAAD Model and Treatment Regimen
Three-week-old C57BL/6 mice weighing 9 to 11 g were purchased from Beijing Vital River Laboratory Animal Technology Co. Ltd (Beijing, China) and housed in a specific pathogen-free environment at Tongji Medical College. The mice were randomly divided into the following 3 groups (n = 14 each) and treated accordingly for 4 weeks: (1) sham group: given normal drinking water with intraperitoneal injection of 0.5% DMSO; (2) control group: administered 1 mg/kg BAPN dissolved in the drinking water to induce TAAD along with DMSO injections; and (3) TEPP-46 group: administered BAPN along with intraperitoneal injections of 5 mg/kg/d TEPP-46. The BAPN solution was freshly prepared every day, and the mice were fed normal chow. The thoracic aortas were harvested on day 28 of the regimen after ultrasonic imaging, and fixed overnight in 4% paraformaldehyde or snap frozen in liquid nitrogen. The Institutional Animal Care and Use Committee of Tongji Medical College approved all animal experiments.
Echocardiography
Ultrasonic cardiograms were obtained using the Vevo 1100 Microimaging System with a 30-MHz transducer (VisualSonics, Inc, Australia). Images of the ascending aorta were captured from a parasternal long-axis view of the section as reported previously. The measurements were performed in triplicates in each mouse, and the technician was blinded to the experimental groups.
Quantitative Real-Time Polymerase Chain Reaction
Total RNA was isolated from frozen thoracic aorta samples or BMDMs using Trizol reagent, and 500 ng RNA per sample was reversed-transcribed into complementary DNA using the PrimeScript RT Master Mix (Takara, Shiga, Japan) according to the manufacturer’s instructions. Quantitative real-time polymerase chain reaction (RT-PCR) was performed on a StepOne Plus Real-time PCR System (Applied Biosystems, Foster City, California) using SYBR Green I reagent. The reaction parameters were as follows: 95°C for 30 seconds, followed by 40 cycles of 95°C for 5 seconds, and 60°C for 1 minute. The genomic copy number was quantified by comparative ΔΔCt method, and the mRNA expression levels of the target genes were normalized to β-actin. The primers are listed in the Supplementary Table S1.
Western Blotting
The harvested BMDMs and tissue samples were lysed with RIPA lysis buffer supplemented with protease and phosphatase inhibitor cocktails. The protein concentration was measured using Pierce BCA Protein Assay kit (Pierce), and equal amount of protein per sample were boiled in nondenaturing Gel Sample Loading Buffer (5×). The protein samples were loaded onto and run through a BeyoGel Plus Precast PAGE Gel (Hepes, 10%, 10 well, P0508S; Beyotime, Shanghai, China) for Tris-Gly System and transferred to a polyvinylidene difluoride membrane. The latter was blocked with 5% skimmed milk powder for 1 hour and incubated overnight with the primary antibodies at 4°C on a shaker. After washing with Tris-buffered saline Tween, the membrane was incubated with peroxidase-conjugated secondary antibody for 1 hour at 37°C. The blots were developed using ECL reagent, and gray values was measured by Image J software (v1.8.0) and normalized to β-actin or GAPDH. The antibodies used are listed in the Supplementary Table S2.
Enzyme-Linked Immunosorbent Assay
Serum levels of IL-1b and IL-18 were measured using commercially available enzyme-linked immunosorbent assay kits according to the manufacturers’ instructions.
Primary BMDM Culture
The femurs and tibia were harvested from 8-week-old male C57BL/6 mice, and the epiphyses were cut using sterile micro-dissecting scissors. A 1-mL sterilized syringe needle was inserted in the bone cavity, and the marrow was flushed out thrice with phosphate-buffered saline till the bones became pale. The red blood cells were lysed using the ACK lysis buffer, and the remaining cells were passed through a 0.45-µm nylon mesh. The cells were washed and resuspended in Dulbecco modified Eagle medium containing 10% fetal bovine serum (FBS) and 1% penicillin–streptomycin, and cultured at 37°C under 5% CO2.
Histology and Immunostaining
The fixed thoracic aorta samples were embedded in paraffin and cut into 5-μm-thick sections for hematoxylin and eosin (H&E) staining, elastin von Gieson (EVG), and Masson trichrome staining according to standard protocols. For immunostaining, the aortic sections were cleared with xylene, rehydrated through an ethanol gradient, and rinsed with distilled water. After soaking in boiling citric acid solution for 10 minutes to unmask antigens, the sections were blocked with 10% FBS. The processed sections were incubated overnight with the primary antibodies at 4°C, rewarmed for 45 minutes at room temperature, and then incubated with the secondary antibodies for 1 hour. The stained sections were observed under a fluorescence microscope.
Human Aortic Sample Analysis
Thoracic aortic wall specimens were collected from 4 patients with TAAD who underwent open surgery and 4 who underwent aortic surgery. Non-aneurysmal abdominal aortic wall specimens were used as controls. Inflamed or rapidly expanding aneurysms were eliminated from the analysis. Informed consent was obtained from all patients, and the study was approved by a local medical ethics committee for clinical studies.
Statistical Analyses
The data are expressed as the mean ± standard deviation. All data were analyzed using Prism 6.0 software (GraphPad Software, California) and evaluated for equal variance and normality. Two-tailed Student t test and 1-way analysis of variance with Bonferroni post hoc test were used to, respectively, compare 2 and multiple groups. The survival curve was plotted using the Kaplan-Meier method and analyzed using the log-rank test. P values <.05 were considered statistically significant.
Results
Levels of PKM2 Are Elevated in TAAD Lesions
Figure 1A is a schematic illustration of key metabolites and genes involved in glucose metabolism. Pyruvate kinase M2 is a rate-limiting enzyme in the conversion of phosphoenolpyruvate to pyruvate in the glycolysis. Thoracic aorta tissues from TAAD patients showed significantly higher PKM2 messenger RNA (mRNA; Figure 1B) and protein levels compared to the healthy tissues (Figure 1C-F). In addition, the change in the expression levels of PKM2 was more significant compared to the other rate-limiting enzymes. We also established a murine model of TAAD by administering BAPN to the mice for 28 days as described previously by Ren et al. 31 The incidence of TAAD among BAPN-treated mice was 85.7%, and the BAPN group showed aneurysms in the thoracic aorta and 14.3% of which died due to the rupture. No spontaneous TAAD developed in the sham group mice (Figure 1G). The aortic tissues of the TAAD mice also showed elevated levels of PKM2 mRNA (Figure 1H) and protein (Figure 1I and J) compared to the sham group mice. Taken together, PKM2 is upregulated during TAAD and therefore has therapeutic potential.

Pyruvate kinase M2 (PKM2) expression was elevated in thoracic aortic aneurysm and dissection (TAAD). A, Schematic map illustrating key metabolites and genes in glucose metabolism. B, Glycolysis-related genes (including HK2, GLUT1, PFKB3, PKM2, LDHA, and PDK2) and messenger RNA (mRNA) levels were determined by real-time quantitative polymerase chain reaction (RT-qPCR) in aortic tissues from TAAD patients (n = 4 per group) compared to control specimens (n = 4). C and D, The protein expression of PKM2 was determined using Western blot analysis. Quantification is shown on the right. E and F, Immunohistochemistry staining of PKM2 in human aortic tissues. Quantification is shown on the right. G, Gross appearance of the thoracic aortic aneurysm and dissection rupture from β-aminopropionitrile fumarate (BAPN)–induced mice compared with sham group. H, Glycolysis-related genes mRNA levels was determined by qPCR in mice aortic tissues (n = 6 per group). I and J, Immunofluorescence staining of PKM2 in mice aortic tissues. Quantification is shown on the right. A 2-tailed Student t test was used for comparisons between 2 groups. The data are presented as the means ± standard deviation (SD) for each group. *P < .05. Scale bars: 1000 µm (×40) 200 µm (×200 and ×400); 100 µm (×400). NS indicates no significance.
The PKM2 Activator TEPP-46 Alleviated TAAD in the BAPN-Treated Mice
To determine the role of PKM2 in TAAD, we concurrently administered the PKM2 activator TEPP-46 or dimethyl sulfoxide (DMSO; vehicle) in addition to BAPN. After testing different concentrations of TEPP-46 in preliminary experiments (Supplementary Figure 1), the dosage of 5 mg/kg was selected. During the 28-day BAPN inducement, progressive dilation was seen in the region between the ascending and the descending thoracic aorta, which was significantly decreased upon administration of TEPP-46 (Figure 2A). In addition, the TEPP-46-treated mice showed decreased TAAD incidence and related morbidity, along with higher survival rates (Figure 2B and C). Furthermore, ultrasonic imaging revealed aortic luminal expansion during BAPN inducement, which was also decreased by TEPP-46 (Figure 2D and 2E).

Pyruvate kinase M2 (PKM2) activator TEPP-46 alleviated the formation of thoracic aortic aneurysm and dissection (TAAD) in from β-aminopropionitrile fumarate (BAPN)–induced mice. A, Representative photographs showed the visible changes in the BAPN-induced TAAD model from different groups (indicated by the yellow arrows). B, The Kaplan-Meier survival curve showed the survival of mice from 2 groups 4 weeks after BAPN-induced (BAPN + vehicle, n=14; and BAPN + TEPP-46, n=14; log-rank test). C, Morbidity and TAAD incidence of 2 groups were counted. D, Typical ultrasound images of the thoracic aortas from each group on day 21 after BAPN-induced TAAD. E The luminal diameter of the ascending aorta was echocardiographically measured (n = 6/group). A 2-tailed Student t test was used for comparisons between 2 groups, and the log-rank test was used for the evaluation of Kaplan–Meier survival curves. The data are presented as the means ± standard deviation (SD) for each group. *P < .05. NS indicates no significance.
TEPP-46 Protects the Structural Integrity of the Aortic Wall in the BAPN-Induced TAAD Model
BAPN significantly increased the thickness of the ascending aorta wall compared to that in the control (Figure 3A), which was alleviated by TEPP-46 (Figure 3A and B). The development of an aortic aneurysm is accompanied with vascular remodeling and loss of the vascular smooth muscle cells (VSMCs). First of all, we detected the expression of PKM2 in the ascending aorta wall, and BAPN significantly increased the expression of PKM2, which was not obviously changed by TEPP-46 (Figure 3C and D). As shown in Figure 3C, the α-SMA-positive areas were significantly decreased upon induction of TAAD, while concurrent TEPP-46 treatment prevented the loss of α-SMA-positive VSMCs (Figure 3E and F). In addition, EVG staining showed parallel and uniformly arranged elastin fibers in the thoracic aorta of the sham group mice and disrupted and irregularly arranged fibers in the untreated TAAD mice. TEPP-46 treatment restored the density of the elastin laminae (Figure 3G and H). Since the media and adventitia of aortic walls are replaced by fibrous connective tissue during TAAD development, we next analyzed the extent of collagen deposition with Masson’s trichrome staining. β-aminopropionitrile fumarate–induced TAAD significantly increased the deposition of collagen and fibrin around the smooth muscle cells, which was alleviated by TEPP-46 (Figure 3I and J). Taken together, TEPP-46 protected the structural integrity of the aortic wall in BAPN-induced TAAD.

TEPP-46 treatment protects the structural integrity of aortic wall in the β-aminopropionitrile fumarate (BAPN)–induced thoracic aortic aneurysm and dissection (TAAD) model. A and B, Hematoxylin and eosin (H&E) staining and aortic wall thickness analysis of the ascending and descending aortas (n = 6/group). C and D, Representative immunostaining of pyruvate kinase M2 (PKM2) and the semiquantitative histological analysis (n = 6/group). E and F, Microscopic examination of vascular smooth muscle cells and vascular smooth muscle cells destruction score (n = 6/group). G and H, Verhoeff Van Gieson (EVG) staining and elastin fragmentation score of the ascending and descending aortas (n = 6/group). I and J, Trichrome staining and deposition of collage score of the thoracic aortas (n = 6/group). The data are presented as the means ± standard deviation (SD) for each group. Scale bars: 1000 µm (×40) 200 µm (×100) and 100 µm (×400).*P < .05.
TEPP-46 Significantly Reduced the Secretion of MMPs and IL-1β in the BAPN-Induced TAAD Model
Inflammation plays a key in the initiation and progression of TAAD. Accordingly, we found that the increased infiltration of CD68+ monocytes/macrophages was more obvious than the Gr-1+ neutrophils and CD3+ T cells into the aortic wall of the TAAD mice, which was significantly decreased in the TEPP-46-treated group (Figure 4A and B). Furthermore, we detected the expression of PKM2 and IL-1β in the ascending aorta wall, BAPN apparently increased the expression of PKM2, which was not obviously changed by TEPP-46 (Figure 4C and D). However, TEPP-46 treatment significantly reduced IL-1β levels in the aortic wall and serum (Figure 4C-E), in addition to the circulating levels of IL-18 and caspase 1 (Figure 4E and F). Given the extensive elastolysis seen during TAAD, the expression of MMPs in the aortic wall was also evaluated. MMP-2, 9, and 13 were significantly increased in both the aortic walls (Figure 4G and H) and the sera (Figure 4I) of the untreated BAPN-induced group and restored to normal levels by TEPP-46. Taken together, TEPP-46 attenuated the inflammatory response and decreased MMPs levels during TAAD.
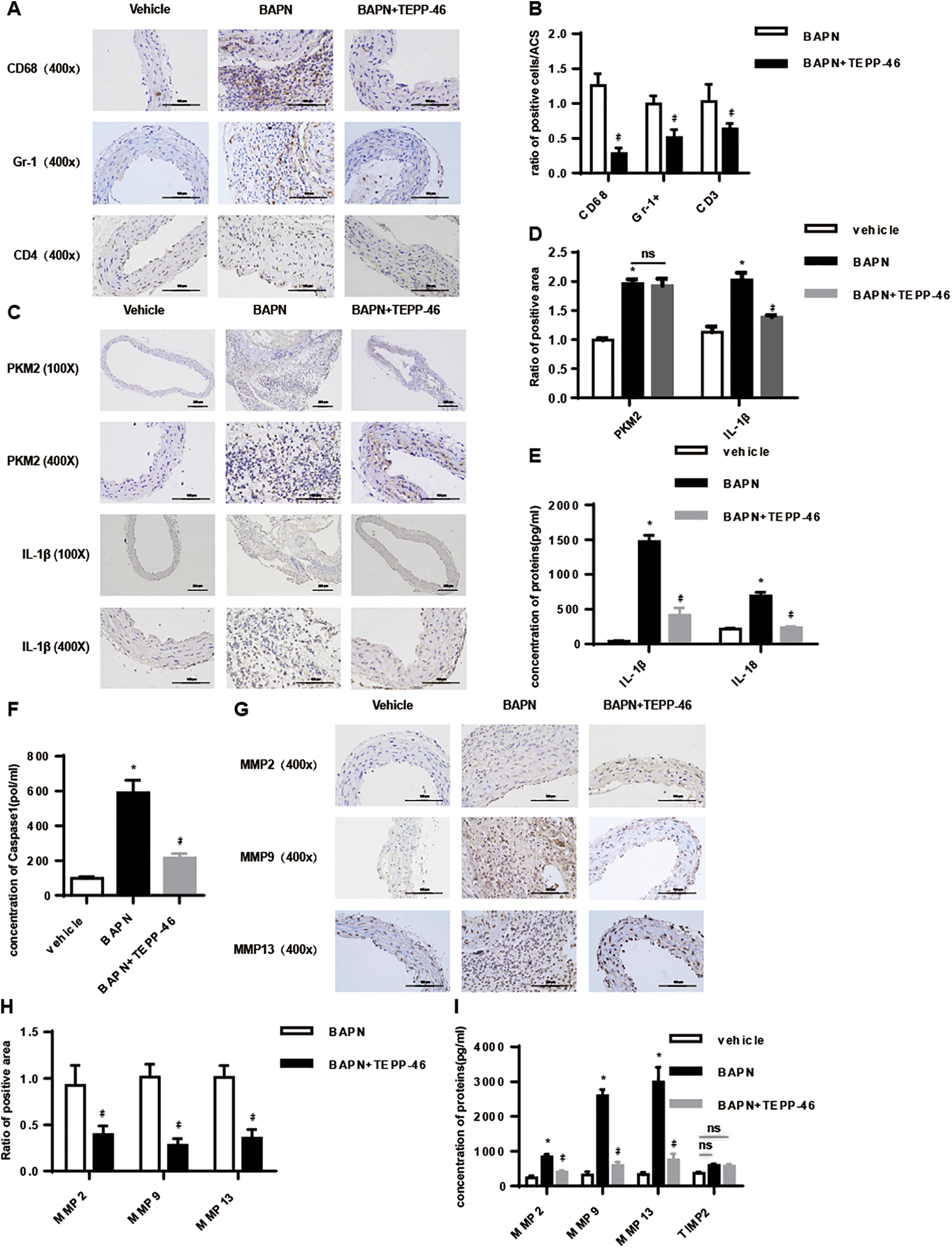
TEPP-46 treatment significantly reduced the secretion of metal matrix proteinases and interleukin 1β (IL-1β) in the β-aminopropionitrile fumarate (BAPN)–induced thoracic aortic aneurysm and dissection (TAAD) model. A and B, Microscopic examination of macrophage, neutrophil, CD4+ T-cell infiltration, and the semiquantitative histological analysis (n = 6/group). C and D, Representative immunostaining of pyruvate kinase M2 (PKM2) and IL-1β and the semiquantitative histological analysis was conducted (n = 6/group). E and F, Enzyme-linked immunosorbent assay (ELISA) showed IL-1β, IL-18, and caspase-1 concentration in serum (n = 6/group). G and H, Representative immunostaining of matrix metalloproteinase (MMP)-2, MMP-9, and MMP-13 expression in arterial walls (n = 6/group). I, ELISA showed MMP-2, MMP-9, MMP-13, and TIMP2 concentration in serum (n = 6/group). A 2-tailed Student t test was performed for comparisons between 2 groups and 1-way analysis of variance (ANOVA) with Bonferroni post hoc test were used to, respectively, compare multiple groups. The data are presented as the means ± standard deviation (SD) for each group. *P < .05 compared to vehicle group; #P < .05 compared to BAPN group. Scale bars: 200 µm (×100) and 100 µm (×400).
TEPP-46 Decreased ROS Levels and Inflammasome Activation in the BAPN-Induced TAAD Model
Numerous studies have implicated a critical role of ROS in TAAD development. Consistent with this, we observed intensive dihydroethidium staining in the aortic walls of BAPN-induced mice, indicating robust production of ROS. TEPP-46 treatment significantly decreased the ROS levels compared to the untreated mice (Figure 5A and B). Generation of ROS is closely associated with the activation of NLRP3 in the macrophages during aneurysm. 32 -34 The BAPN-induced mice showed elevated levels of NLRP3, ASC, and caspase-1 in the CD68+ macrophages of the aortic wall, clearly indicating inflammasome activation. TEPP-46 treatment significantly decreased inflammasome activation in vivo (Figure 5C-H) as well as in vitro (Supplementary Figure 2). Taken together, TEPP-46 treatment decreased ROS production and inflammasome activation during TAAD development.

TEPP-46 treatment reduces reactive oxygen species (ROS) levels and inflammasome activation in the aortic wall of β-aminopropionitrile fumarate (BAPN)–induced thoracic aortic aneurysm and dissection (TAAD) model. A and B, Dihydroethidium (DHE) staining (red fluorescence) was performed to assess ROS levels. Representative images of DHE staining and ratio of ROS density from aortic wall segment of sham, BAPN, and TEPP-46 group (n = 6/group). C and D, Immunofluorescence staining for NLRP3 (green), CD68 (red), and ratio of NLRP3 intensity in thoracic aortas of each group (n = 6/group). E and F, Immunofluorescence staining for apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC; green), CD68 (red), and ratio of ASC intensity in thoracic aortas of each group (n = 6/group). G and H, Immunofluorescence staining for caspase 1 (green) CD68 (red), and ratio of intensity in thoracic aortas of each group (n = 6/group). A 2-tailed Student t test was used for comparisons between 2 groups. The data are presented as the means ± standard deviation (SD) for each group. *P < .05. Scale bars: 50 µm (×400). NS indicates nonsignificance.
TEPP-46 Inhibited Inflammasome Mediated IL-1β Secretion by Inactivating the STAT3/ HIF1α Signaling Pathway
To determine the mechanistic basis of TEPP-46-mediated inflammasome inhibition, we next analyzed the expression levels of its mediators and PKM2. In aorta tissues of the BAPN-induced TAAD, BAPN increased expression levels of total PKM2, which was not obviously changed by TEPP-46 (Figure 6D and E). Pyruvate kinase M2 was transferred from cytosol to nucleus. However, after TEPP-46 treatment, the transfer of PKM2 was weakened (Figure 6A and B). In mRNA level, TEPP-46 restored the levels of NLRP3, ASC, caspase 1, pro-caspase 1, and pro-IL-1β, which were significantly increased during TAAD (Figure 6C). In protein level, while TEPP-46 restored the levels of IL-1β and caspase 1 and NLRP3 were significantly increased during TAAD, it showed no effects on pro-caspase 1 and pro-IL-1β (Figure 6D and E). Since the Akt/GSK-3β signaling pathway is involved in inflammasome activation and the development of aneurysms, we also analyzed the effects of TEPP-46 on this pathway. As shown in Figure 6F and G, the p-AKT and p-GSK-3β levels were decreased in TAAD tissues compared to the aorta tissues of the sham group and were unaffected by TEPP-46. The mitogen-activated protein kinases (MAPKs) p38 and ERK1/2 are also crucial mediators of inflammasome activation, and increased levels of their phosphorylated forms were observed in TAAD lesions compared to normal tissues. However, no significant changes were seen after TEPP-46 treatment (Figure 6H and I). Finally, the STAT3/HIF1α signaling pathway was activated in the aortic walls of the TAAD mice as indicated by the increased levels of p-STAT3 and HIF1α, which was reversed upon TEPP-46 treatment. Additionally, the expression of HIF1α target genes, such as LDHA and PDHK1, was also changed following HIF1α (Figure 6J and K). Taken together, TEPP-46 inhibited inflammasome-mediated IL-1β secretion by targeting the STAT3/HIF1α signaling pathway, without affecting the Akt/GSK-3β or p38MAPK/ERK1/2 pathways.

TEPP-46 treatment inhibited inflammasome mediated interleukin 1β (IL-1β) secretion via HIF-1α and Stat3 suppression. A, Western blotting for pyruvate kinase M2 (PKM2) expression in nucleus and cytosol of the thoracic aortas of each group (n = 3). B, Confocal images were acquired in thoracic aortas of each group (n = 3) and stained with anti-PKM2 (green). Nuclei were localized by DAPI (blue). C, The messenger RNA (mRNA) levels of NLRP3 inflammasome components, including NLRP3, ASC, caspase 1, pro-IL-1β, and pro-IL-18 was determined by quantitative polymerase chain reaction (qPCR) in thoracic aortic tissues from thoracic aortic aneurysm and dissection (TAAD) models (n = 6 per group) compared to vehicle specimens (n = 6). D and E, The protein levels of PKM2 and NLRP3 inflammasome components in aortic tissues and secretion of casapase-1 and IL-1β in serum of vehicle, β-aminopropionitrile fumarate (BAPN), and TEPP-46 group (n = 3). F and G, Western blotting for p-AKT and p-GSK-3β expression in the thoracic aortas of each group (n = 3). H and I, Western blotting for p-p38 and p-ERK1/2 expression in the thoracic aortas of each group (n = 3). J and K, Western blotting for p-STAT3, T-STAT3, HIF1-α, LDHA, and PDHK1 expression in the thoracic aortas of each group (n = 3). A 2-tailed Student t test was used for comparisons between 2 groups and 1-way analysis of variance (ANOVA) with Bonferroni post hoc test were used to, respectively, compare multiple groups. The data are presented as the means ± standard deviation (SD) for each group. *P < .05 compared with vehicle group; #P < .05 compared with BAPN group. Scale bars: 50 µm (×400). NS indicates no significance.
Discussion
Dysfunctional VSMCs decrease the ability of the thoracic aorta wall to withstand the biomechanical forces of circulation, resulting in progressive enlargement, tearing of the inner layer, and dissection through the medial layer by blood, a phenomenon known as TAAD. 35 It is a fatal condition that can only be managed by surgical intervention, and noninvasive, pharmacological therapies are currently lacking. 36 We found elevated levels of the glycolytic enzyme PKM2 in the TAAD lesions, indicating metabolic reprogramming and therefore tested the therapeutic potential of its activator TEPP-46 against TAAD. TEPP-46 not only increased the survival of the TAAD mice but also significantly attenuated aortal wall thickness and other pathological changes induced by BAPN. Mechanistically, TEPP-46 decreased the secretion of IL-1β and MMPs, ROS production, and macrophage infiltration and alleviated inflammasome activation via STAT3/HIF1-α signaling pathway.
Since local inflammation plays a key role in TAAD pathogenesis, targeting pro-inflammatory cytokines would be a feasible therapeutic strategy. 37 Indeed, Johnston et al reported that disrupting IL-1β signaling suppressed experimental aortic aneurysm, and since then IL-1β has been recognized as an effective target for treating various inflammatory conditions such as gout, coronary artery disease, and diabetes mellitus. 38 -41 The NLRP3 inflammasome regulates the production of IL-1β and is triggered by metabolic substrates such as cholesterol and glucose. 42,43 Consistent with this, the inflammasome components such as NLRP3, ASC, and caspase 1 were elevated in the thoracic aorta of the TAAD mice and attenuated after TEPP-46 treatment. Taken together, inflammasome activation mediates the initiation and progression of TAAD and can be inhibited by TEPP-46.
The inflammatory response is often accompanied by the production of ROS, which is mainly secreted by the macrophages. It not only aggravates ECM degradation but also induces apoptosis in VSMCs. 44 -47 The ROS levels were significantly higher in the thoracic aorta of BAPN-induced mice and decreased following TEPP-46 treatment. MMPs mediated degradation of the elastin lamina, and collagen of aorta ECM accelerates the progression of TAAD. 48 Longo et al reported that no aneurysms were formed in 10-week MMP-9 or MMP-2 knockout mice even after abluminal application of calcium chloride. 49 Consistent with the results so far, TEPP-46 also significantly decreased MMP-2 and MMP-9 levels in the aorta of TAAD mice.
The AKT/GSK-3β signaling pathway is extensively involved in various inflammatory diseases. 50,51 AKT is inhibited in human aneurysmal aortic tissues, and Akt-2 knockout deteriorates Ang II-induced abdominal aortic aneurysms. 52,53 In addition, AKT/GSK-3β also activates the NLRP3/IL-1β pathway. 54,55 We found that both AKT and GSK-3β were inhibited in the TAAD lesions compared to normal tissues and were not affected by TEPP-46. The p38MAPK/ERK1/2 pathway regulates the expression of numerous pro-inflammatory and oxidative stress-inducing genes and also activates the inflammasome. 56 -58 Consistent with previous studies, 59 the p38MAPK/ERK1/2 pathway was activated in TAAD mice but remained unaffected by TEPP-46.
Pyruvate kinase M2 is one of the rate-limiting enzymes in the glycolytic pathway and exists primarily as an enzymatically inactive monomer or dimer. The inactive form can translocate to the nucleus and interact with transcription factors mediating the expression of downstream genes. 60 Demaria et al reported that PKM2 activated STAT3/HIF1α signaling in tumor cells, and Shirai et al found that targeting PKM2 inactivated STAT3 and inhibited IL-1β secretion by CAD macrophages. 61,62 This finding certainly indicated a functional relationship between PKM2 and NLRP3 inflammasome. TEPP-46 specifically binds to PKM2 forming a tight tetramer, which prevents its nuclear translocation. TEPP-46 attenuated the NLRP3 inflammasome-mediated production of IL-1β by inhibiting STAT3/ HIF1α signaling. In addition, although all key enzymes of the glycolytic pathway were upregulated in TAAD, 63 PKM2 showed the most significant change, indicating its therapeutic utility compared to the other enzymes.
In summary, the PKM2 activator TEPP-46 attenuated BAPN-induced TAAD by inhibiting NLRP3-mediated IL-1β secretion. PKM2 activation is therefore a promising therapeutic strategy against TAAD.
Supplemental Material
Supplementary_Figure1 - PKM2 Activator TEPP-46 Attenuates Thoracic Aortic Aneurysm and Dissection by Inhibiting NLRP3 Inflammasome-Mediated IL-1β Secretion
Supplementary_Figure1 for PKM2 Activator TEPP-46 Attenuates Thoracic Aortic Aneurysm and Dissection by Inhibiting NLRP3 Inflammasome-Mediated IL-1β Secretion by Sheng Le, Hao Zhang, Xiaofan Huang, Shu Chen, Jia Wu, Shanshan Chen, XiangChao Ding, Shanshan Chen, Jing Zhao, Heng Xu, Jikai Cui, Yanqiang Zou, Jizhang Yu, Lang Jiang, Jie Wu, Ping Ye and Jiahong Xia in Journal of Cardiovascular Pharmacology and Therapeutics
Supplemental Material
Supplementary_Figure2 - PKM2 Activator TEPP-46 Attenuates Thoracic Aortic Aneurysm and Dissection by Inhibiting NLRP3 Inflammasome-Mediated IL-1β Secretion
Supplementary_Figure2 for PKM2 Activator TEPP-46 Attenuates Thoracic Aortic Aneurysm and Dissection by Inhibiting NLRP3 Inflammasome-Mediated IL-1β Secretion by Sheng Le, Hao Zhang, Xiaofan Huang, Shu Chen, Jia Wu, Shanshan Chen, XiangChao Ding, Shanshan Chen, Jing Zhao, Heng Xu, Jikai Cui, Yanqiang Zou, Jizhang Yu, Lang Jiang, Jie Wu, Ping Ye and Jiahong Xia in Journal of Cardiovascular Pharmacology and Therapeutics
Supplemental Material
Supplemental Material, Supplementary_Figure_legends - PKM2 Activator TEPP-46 Attenuates Thoracic Aortic Aneurysm and Dissection by Inhibiting NLRP3 Inflammasome-Mediated IL-1β Secretion
Supplemental Material, Supplementary_Figure_legends for PKM2 Activator TEPP-46 Attenuates Thoracic Aortic Aneurysm and Dissection by Inhibiting NLRP3 Inflammasome-Mediated IL-1β Secretion by Sheng Le, Hao Zhang, Xiaofan Huang, Shu Chen, Jia Wu, Shanshan Chen, XiangChao Ding, Shanshan Chen, Jing Zhao, Heng Xu, Jikai Cui, Yanqiang Zou, Jizhang Yu, Lang Jiang, Jie Wu, Ping Ye and Jiahong Xia in Journal of Cardiovascular Pharmacology and Therapeutics
Supplemental Material
Supplemental Material, Table_1_(1) - PKM2 Activator TEPP-46 Attenuates Thoracic Aortic Aneurysm and Dissection by Inhibiting NLRP3 Inflammasome-Mediated IL-1β Secretion
Supplemental Material, Table_1_(1) for PKM2 Activator TEPP-46 Attenuates Thoracic Aortic Aneurysm and Dissection by Inhibiting NLRP3 Inflammasome-Mediated IL-1β Secretion by Sheng Le, Hao Zhang, Xiaofan Huang, Shu Chen, Jia Wu, Shanshan Chen, XiangChao Ding, Shanshan Chen, Jing Zhao, Heng Xu, Jikai Cui, Yanqiang Zou, Jizhang Yu, Lang Jiang, Jie Wu, Ping Ye and Jiahong Xia in Journal of Cardiovascular Pharmacology and Therapeutics
Supplemental Material
Supplemental Material, Table_2 - PKM2 Activator TEPP-46 Attenuates Thoracic Aortic Aneurysm and Dissection by Inhibiting NLRP3 Inflammasome-Mediated IL-1β Secretion
Supplemental Material, Table_2 for PKM2 Activator TEPP-46 Attenuates Thoracic Aortic Aneurysm and Dissection by Inhibiting NLRP3 Inflammasome-Mediated IL-1β Secretion by Sheng Le, Hao Zhang, Xiaofan Huang, Shu Chen, Jia Wu, Shanshan Chen, XiangChao Ding, Shanshan Chen, Jing Zhao, Heng Xu, Jikai Cui, Yanqiang Zou, Jizhang Yu, Lang Jiang, Jie Wu, Ping Ye and Jiahong Xia in Journal of Cardiovascular Pharmacology and Therapeutics
Footnotes
Authors’ Note
All the animal experimental protocols were approved by the Ethics Committee of Union Hospital, Huazhong University of Science and Technology, China.
Author Contribution
Sheng Le and Hao Zhang contributed equally to this article as first authors.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: This work was supported by the National Natural Science Foundation of China (81730015, 81571560, 81974048, 81701585, 81570325, and 81800413) and the Natural Science Foundation of Hubei Province (2017CFB357 and 2019AAA032).
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.