
Abstract
Influenza D virus (IDV) is considered a new agent involved in bovine respiratory disease (BRD). Based on seroprevalence studies or isolation from clinical samples, this virus has been detected on several continents and in several animal species, including cattle, pigs, camel, horses, and goats. We used an indirect in-house ELISA to detect anti-IDV antibodies in 165 serum samples from bulls on 116 farms in the province of La Pampa, Argentina. Eighty-five of 116 (73%) farms had at least 1 positive animal, and 112 of 165 (68%) of the analyzed samples were positive. There were no significant differences in the proportion of seropositive samples depending on the geographic region in which the samples were taken. Our results suggest that IDV infection is endemic in La Pampa; the clinical importance of IDV in Argentina remains to be investigated.
Bovine respiratory disease (BRD) is one of the disease complexes that generates the most significant production losses and consequently the greatest economic losses in feedlots.1,7 Combined with predisposing factors, viruses and bacteria are involved in the development of BRD. Several viruses can initiate BRD and, although some generate severe clinical signs by themselves, others contribute to the onset of disease and predispose to secondary bacterial infections that require antimicrobial treatment. 7 The most commonly detected viral BRD pathogens are bovine respiratory syncytial virus (BRSV; Bovine orthopneumovirus), bovine parainfluenza virus 3 (BPIV-3; Bovine respirovirus 3), bovine coronavirus (BCoV; Betacoronavirus 1), bovine herpesvirus 1 (BoHV-1; Bovine alphaherpesvirus 1), and bovine viral diarrhea virus (BVDV; Pestivirus).
In 2011, a new virus of the Orthomyxoviridae family was discovered in pigs with respiratory signs in the United States.8,9 Subsequently, several studies classified this virus as influenza D virus (IDV), and the virus and/or IDV-specific antibodies were detected in cattle, horses, sheep, goats, and camels on several continents.2–6,9–14,19 Moreover, an investigation based on hemagglutination inhibition (HI) in the United States demonstrated serologic evidence of IDV infection in 91% of sampled people who worked with cattle, which highlights possible zoonotic aspects of this emerging pathogen. 20
Experimental infections of cattle demonstrated replication of the virus in the respiratory tract, and correlations were made between clinical signs of BRD and the detection of IDV in beef and dairy cattle farms.13,15,17 It also became clear that the virus can be transmitted both by direct contact and aerogenously, and that it causes mild lesions in the upper and lower respiratory tract of cattle. 17 The above studies suggest that IDV may play a role in BRD, and that this virus is widespread.
Whereas some of these investigations, such as those performed in Italy 6 and Luxembourg, 19 demonstrated high seroprevalence (92.4% and 80.2%, respectively), other studies identified large variations of the seropositivity rate in different parts of various countries. 10 Based on this information and the lack of data from South America, we investigated this new agent in cattle in the province of La Pampa, a major beef-raising area in Argentina.
We used 165 serum samples, which had been collected in 2013 to estimate the seroprevalence of reproductive diseases in bulls, such as bovine brucellosis, and had been stored in the serum bank of the Instituto Nacional de Investigación Agropecuaria (INTA) in Anguil, La Pampa. All animals sampled were bulls > 3 y old, dedicated to reproduction in extensive beef-breeding systems. No information was available about the occurrence of respiratory disease in these bulls. We analyzed 1–3 samples from each of 116 farms. For data analysis, the province of La Pampa was divided into 3 geographic zones according to the territorial division established in the INTA regional projects (Fig. 1).
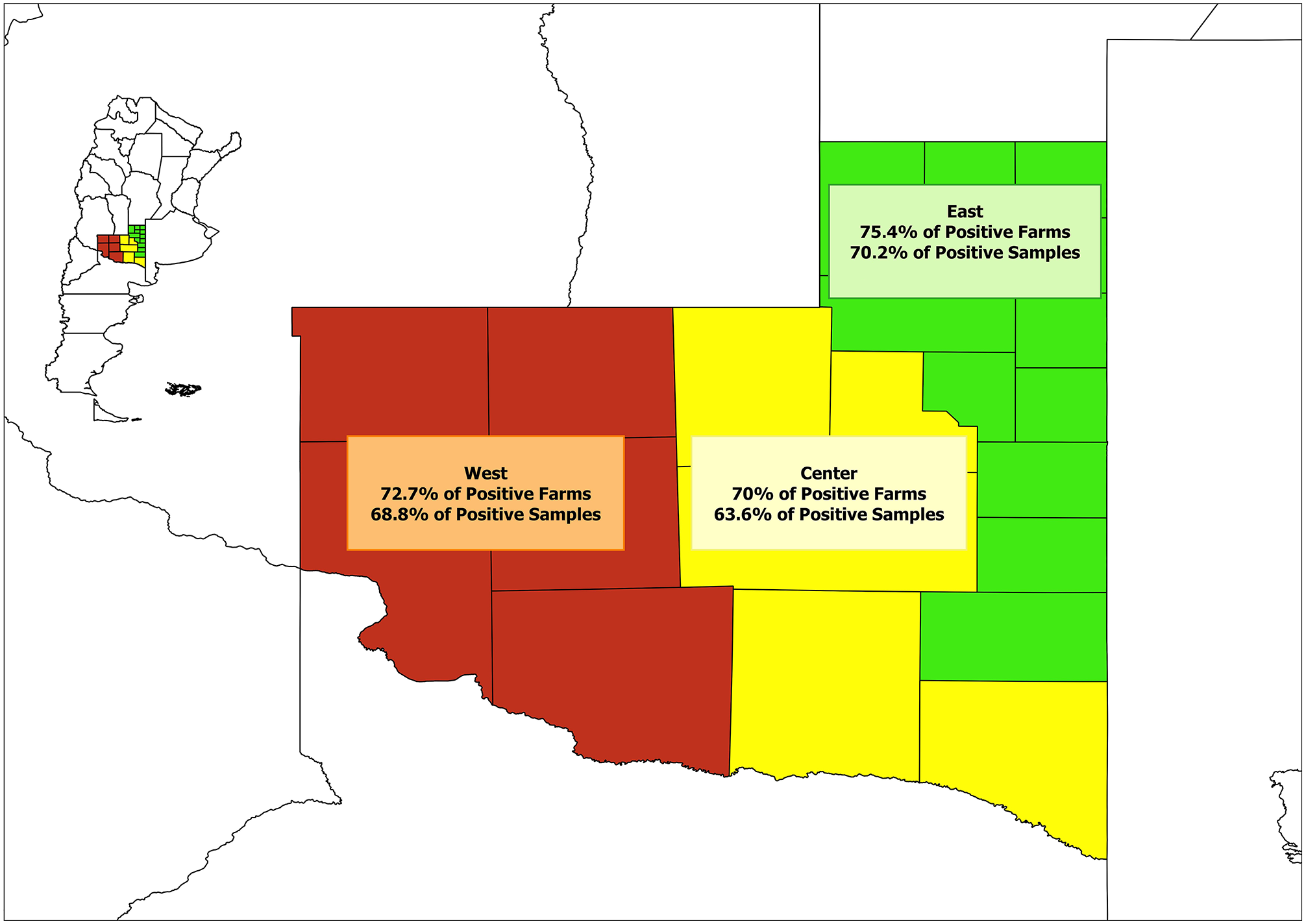
Geographic zones of La Pampa from which samples were collected from bulls to be tested for anti–influenza D virus antibodies.
We used an in-house indirect ELISA as described previously. 17 Briefly, 96-well plates were coated with 0.05 M sodium carbonate–bicarbonate buffer containing lysates of cells infected with a French IDV strain (D/bovine/France/5920/2014) and noninfected cells, and were incubated at 4°C overnight. The plates were then washed twice in washing buffer consisting of phosphate-buffered saline (PBS) 0.05% (v/v) Tween 20 (PBS-T) and were blocked with the same buffer, at 25°C for 1 h. Serum samples and control sera were diluted 1:50 with washing buffer and were added in a volume of 100 µL per well; the plates were incubated at 37°C for 1 h, and then washed 3 times as described above. A conjugate (BRSV-Ab kit; Boehringer Ingelheim Svanova) was diluted 1:2 in PBS-T, 100 μL added per well, and incubated for 1 h at 37°C. After incubation, the plates were again washed as described above, and 100 µL of 3,3’,5,5’-tetramethylbenzidine (TMB) substrate (Boehringer Ingelheim Svanova) was added per well, and incubated at room temperature for 10 min. Thereafter, 50 µL of stop solution (Boehringer Ingelheim Svanova) was added per well, and optical density (OD) was analyzed in a spectrophotometer at 450 nm. Corrected OD (COD) values were calculated by subtracting OD values of sera tested on control antigen from those obtained when the same sera were tested on IDV antigen. To validate this ELISA, samples from natural infections and experimental infections were used that had been tested previously by HI and ELISA. The HI assays were performed using standard techniques as described previously, 21 and samples were considered positive if antibody titers were ≥ 1:20. The control serum was derived from a calf that had been experimentally infected with IDV strain D/bovine/France/5929/2014 (kindly provided by Drs. M. Ducatez and G. Meyer, Université de Toulouse, Toulose, France). For the ELISA, the positive control serum used was the same as that used for HI, and the negative control serum used was derived from a Swedish calf with no clinical signs of BRD and with negative HI results. Results were expressed as percentage of the COD of a positive control serum with a HI titer of 1:1,024. A serum was considered positive for IDV antibody if the COD was > 40% of the positive control.
To validate our ELISA, 300 random samples of bovine sera collected between 2016 and 2017 and stored in the serum bank of the National Veterinary Institute, SVA, Department of Microbiology, Uppsala, Sweden were analyzed by ELISA and HI. All of the samples were negative by both techniques. In addition, 37 bovine serum samples were tested with both HI and ELISA to compare specificity and sensitivity between the 2 assays. These samples included sera from 25 cattle in which a natural IDV infection had been detected by RT-qPCR (kindly provided by Dr. C. Snook, Luxembourg Institute of Health) and from 12 experimentally infected cattle, obtained 3 wk post-inoculation (kindly provided by Drs. Ducatez and Meyer, France). Among the naturally infected animals, 19 of 25 (76%) were positive by ELISA and 18 of 25 (72%) were positive by HI. Among the experimentally infected animals, 12 of 12 (100%) were positive by ELISA and 10 of 12 (83%) were positive by HI (> 1:20). Another 2 of 12 (16%) had HI titers below the cutoff of 1:10.
These results suggest that the ELISA and HI tests performed similarly, with maybe a slightly higher sensitivity for the ELISA (Table 1). Furthermore, performing kappa analysis of both tests, an index value of 0.94 was obtained, demonstrating almost perfect correlation between ELISA and HI.
Differences between ELISA and hemagglutination inhibition detection of influenza D virus antibody in sera from experimentally and naturally infected cattle.

Our in-house ELISA was then used to analyze the 165 samples collected in La Pampa. We found an overall seroprevalence rate in bulls in La Pampa of 68% (112 of 165), with an average COD of 53% of the positive control (Table 2). In total, 73% of the farms had 1 or more seropositive animal among 85 of 116 animals tested, and, in the farms in which 3 samples were taken, there was always at least 1 positive animal (Table 3).Thus, as for other viral agents involved in BRD in Argentina, IDV is present widely, at least in extensive beef-breeding systems. 16 By using the chi-squared test, no statistical differences were observed among the 3 regions, either in the number of positive animals (p = 0.70) or in the number of positive farms (p = 0.83), despite a difference in animal density, suggesting that animal density did not influence seroprevalence.
Summary of the serologic survey for influenza D virus in bulls in La Pampa, Argentina.

Number of positive and negative farms based on the number of samples analyzed for each farm.

Although the diagnostic sensitivity of the ELISA was at least as good as the HI test, we cannot exclude that some IDV strains circulating in La Pampa do not cross-react perfectly with the strain used in our assays. Consequently, we may have underestimated the real prevalence of IDV.
Our results suggest that IDV is endemic in cattle in La Pampa, as in other countries for which data exist.2–6,9–14,19 Bulls in breeding systems such as those used in our study are not usually an animal category that suffers from respiratory diseases in Argentina; however, they can be used as sentinel animals given their long stay in herds and by being in contact with all of the cattle in the herd. By testing mature bulls, we monitored a prolonged time of possible exposure in the farms, and our data suggest that IDV has infected cattle in many herds in La Pampa. Research in the United States relates the likelihood of the presence of IDV to the presence of other viruses, such as BVDV. 18 We cannot exclude this possibility, considering that BVDV is endemic in Argentina. Evidence indicates that IDV can infect several animal species, but we do not know the entire range of susceptible animals; wild animals such as deer or wild boar could contribute to the spread of IDV among cattle herds.
None of the large number of BRD vaccines available in Argentina contain IDV. If the role of IDV as a cause of BRD is confirmed, efforts need to be made to characterize the virus present in Argentina in order to develop a vaccine that could protect against this virus.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This project was partly supported by the European Food Safety Authority (EFSA), Partnering Grants (GA/EFSA/AFSCO/2017/01-GA04). We thank the Cells for Life Platform, partly funded by the Infrastructure Committee at SLU, Sweden, for providing facilities and equipment.