
Abstract
The prevalence of organisms known to be associated with bovine respiratory disease (BRD) was investigated in cattle prior to export. A quantitative reverse transcription polymerase chain reaction assay was used to detect nucleic acids from the following viruses and bacteria in nasal swab samples: Bovine coronavirus (BoCV; Betacoronavirus 1), Bovine herpesvirus 1 (BoHV-1), Bovine viral diarrhea virus 1 (BVDV-1), Bovine respiratory syncytial virus (BRSV), Bovine parainfluenza virus 3 (BPIV-3), Histophilus somni, Mycoplasma bovis, Mannheimia haemolytica, and Pasteurella multocida. Between 2010 and 2012, nasal swabs were collected from 1,484 apparently healthy cattle destined for export to the Middle East and Russian Federation. In addition, whole blood samples from 334 animals were tested for antibodies to BoHV-1, BRSV, BVDV-1, and BPIV-3 using enzyme-linked immunosorbent assay. The nasal prevalence of BoCV at the individual animal level was 40.1%. The nasal and seroprevalence of BoHV-1, BRSV, BVDV-1, and BPIV-3 was 1.0% and 39%, 1.2% and 46%, 3.0% and 56%, and 1.4% and 87%, respectively. The nasal prevalence of H. somni, M. bovis, M. haemolytica, and P. multocida was 42%, 4.8%, 13.4%, and 26%, respectively. Significant differences in nasal and seroprevalence were detected between groups of animals from different geographical locations. The results of the current study provide baseline data on the prevalence of organisms associated with BRD in Australian live export cattle in the preassembly period. This data could be used to develop strategies for BRD prevention and control prior to loading.
Introduction
Respiratory disease associated with long distance transport of animals was first reported in the scientific literature in 1925, although the disease had been recognized by veterinarians for at least 20 years prior to that publication. 34 Bovine respiratory disease (BRD) is an important cause of death in cattle exported live from Australia.47,49 Morbidity and mortality due to BRD is the culmination of a combination of host, potential pathogen, and environmental risk factors. As the pathogens involved in fatal BRD on live export voyages from Australia are similar to those associated with BRD in feedlot cattle, 47 it is likely that the pathogenesis of BRD in these 2 cattle productions systems is similar.
Inconsistent access to feed and water, exposure to noise and vibration during transportation, exposure to a new environment, and comingling with unfamiliar animals activate the stress response in cattle (Sainsbury D, Sainsbury P: 1988, Livestock health and housing. In: Report on the welfare of animals during transport. Scientific Committee on Animal Health and Animal Welfare, Brussels, Belgium),45,53,77 causing the release of glucocorticoids and/or catecholamines, which can reduce immunity. 16 An impaired immune response predisposes to increased viral shedding in subclinically affected animals and subsequent proliferation of potentially pathogenic commensal bacteria.45,52 Inhibition of respiratory clearance mechanisms and direct tissue damage by viral infection facilitates proliferation and invasion of secondary bacterial organisms, often resulting in a fatal bacterial pneumonia.52,68
The Australian Government requires animals destined for export to be held in a registered facility (also known as an “assembly depot”) for a minimum period of time prior to being loaded onto a vessel (Australian Standards for the Export of Livestock, Standard 3, http://www.agriculture.gov.au/biosecurity/export/live-animals/advisory-notices/2011/export_advisory_notice_2011-01). Time in the assembly depot allows the animals to recover from road transport, acclimatize to shipboard rations, and receive any health checks or treatments required by the importing country. However, comingling, handling, and sorting of animals that have been recently transported from multiple farms also provides an ideal environment for transmission of viruses and bacteria between animals in the assembly depot.
Viruses implicated in the pathogenesis of BRD include Bovine herpesvirus 1 (BoHV-1), Bovine respiratory syncytial virus (BRSV), Bovine viral diarrhea virus 1 (BVDV-1), Bovine parainfluenza virus 3 (BPIV-3), Bovine coronavirus (BoCV), bovine rhinoviruses, and bovine adenoviruses. Bovine rhinoviruses and adenoviruses probably have only a minimal role in the pathogenesis of BRD, while BoHV-1, BPIV-3, and BRSV are recognized as primary respiratory pathogens, and BVDV-1 as a significant predisposing pathogen (reviewed in previous publications20,52). Since its first isolation in association with respiratory disease in 1993, 67 BoCV has received attention as an emerging BRD pathogen, and is now thought to make a substantial contribution to the development of BRD in feedlot cattle.59,65
Mannheimia haemolytica, Pasteurella multocida, Histophilus somni, and Mycoplasma bovis are considered to be the most important bacteria involved in BRD (reviewed in previous publications29,68). Additional bacterial agents isolated from pneumonic bovine lung tissue include Trueperella (formerly Arcanobacterium) pyogenes78,79 and Bibersteinia (formerly Pasteurella) trehalosi.4,78
The current study reports the pre-export prevalence of viruses and bacteria associated with BRD in Australian live export cattle and identifies a number of factors that influence the prevalence of viruses and bacteria in nasal secretions. The seroprevalence of BoHV-1, BRSV, BVDV-1, and BPIV-3 were also determined. The baseline data generated by the present study will be used to develop pathogen management strategies, both on-farm and at the assembly depot, with a view to reducing the impact of important organisms on morbidity and mortality during live export voyages.
Materials and methods
Study population
Nasal swab samples were collected from 1,484 cattle on 4 voyages between December 2010 and May 2012. The number of cattle sampled represented 1–5% of the total cattle on the voyage (Table 1). The percentage of animals sampled from each cohort ranged from 1.3% to 12.5% (mean = 5.0, median = 4.9, standard error = 1.18).
Voyage data for voyages from which nasal swabs and/or serological samples were collected.
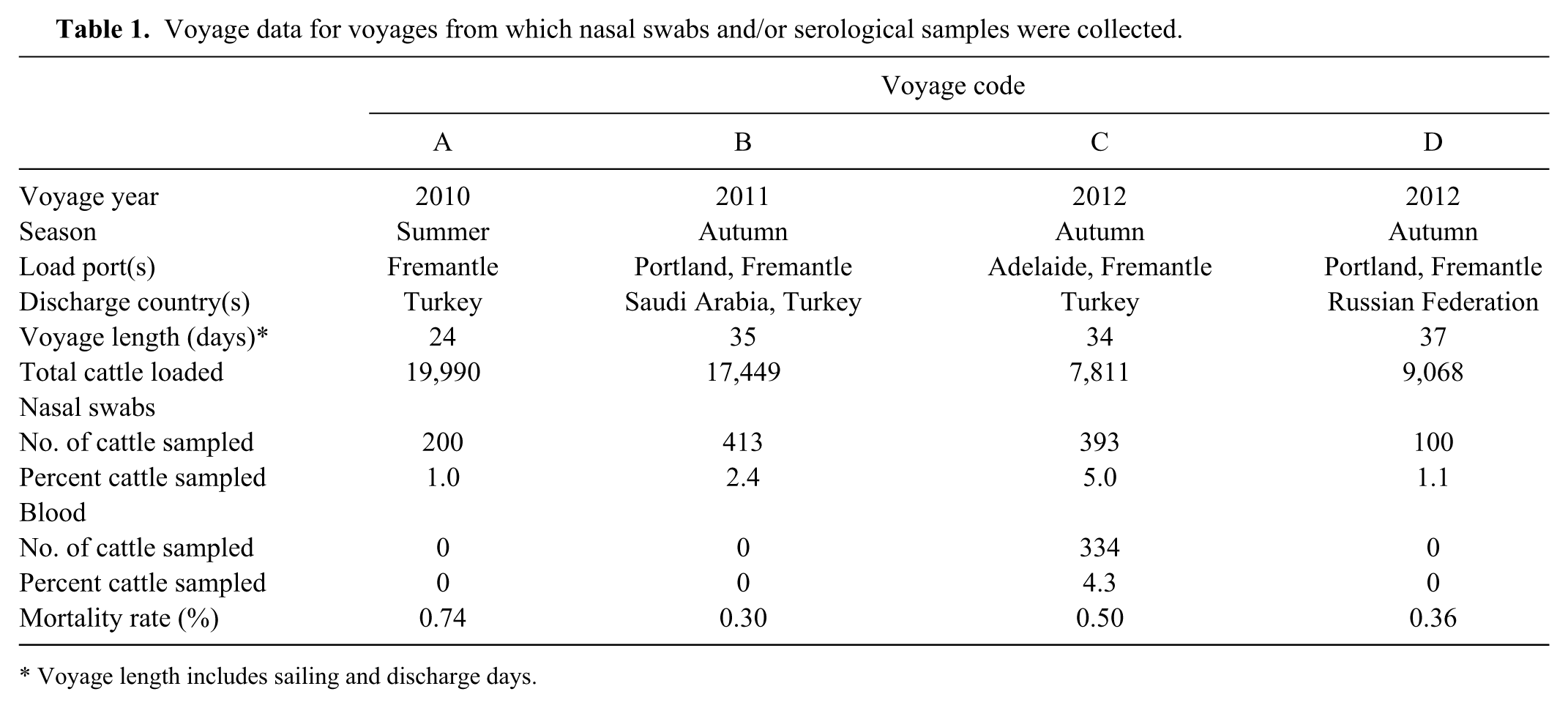
Voyage length includes sailing and discharge days.
Cattle were sampled at 8 assembly depots across 3 states (Table 2): 6 depots in Western Australia (depot codes 1–6) all loading out of Fremantle, 1 in Victoria (depot code 7, load port = Portland), and 1 in South Australia (depot code 8, load port = Adelaide). Animals were grouped into 14 cohorts based on class (feeder, slaughter, breeder) and load-out port (Fremantle, Portland, Adelaide; Table 2).
Voyage, assembly depot, and animal data for cohorts from which nasal swabs ± blood were collected.*
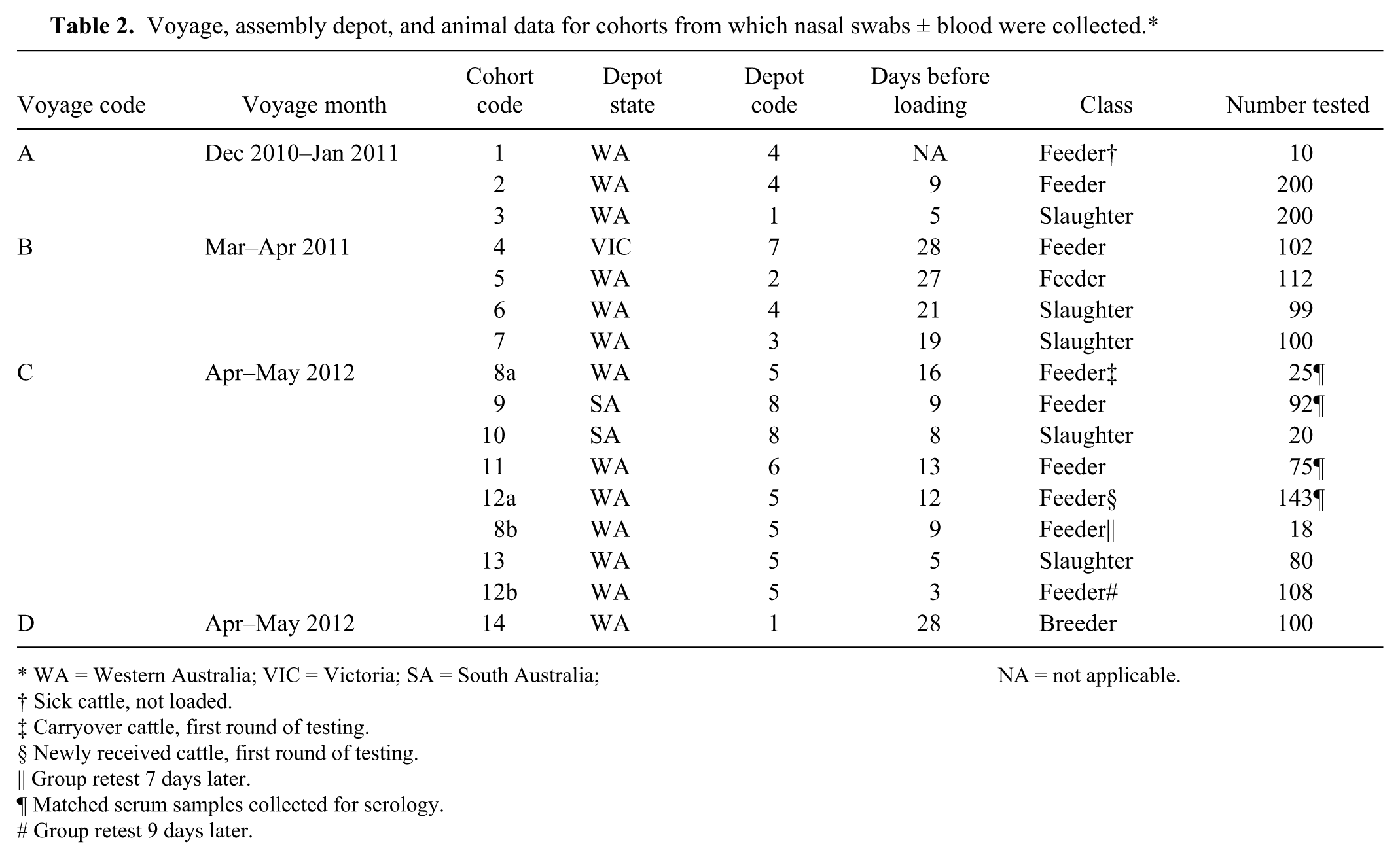
WA = Western Australia; VIC = Victoria; SA = South Australia; NA = not applicable.
Sick cattle, not loaded.
Carryover cattle, first round of testing.
Newly received cattle, first round of testing.
Group retest 7 days later.
Matched serum samples collected for serology.
Group retest 9 days later.
Selection of voyages for sampling was opportunistic and based on the depot induction timetable and location of the assembly depots. Cohorts of cattle were selected to provide a representative subset of the final composition of the shipment (with regard to load port, class, type, and sex) from as many depots and properties as possible, and to collect enough samples to be able to perform statistical comparisons between explanatory variables, while minimizing the impact of study sampling on processing of cattle.
Data collected from each animal at sampling included the radio frequency identification (RFID), property identification code (PIC), breed, sex, and weight. With the exception of RFID, not all data were available for all animals.
For voyages A–C, the consignments comprised a mixture of feeder and slaughter cattle. Voyage D was a shipment of breeder cattle to the Russian Federation. Overall, the class breakdown was 60% feeder, 34% slaughter, and 6% breeder. Eighty-four percent of cattle were Bos taurus and their crosses, and 16% were Bos indicus. Eighty-seven percent of cattle were male (65% steer, 23% bull; 62% feeder, 38% slaughter) and 13% were female (57% breeder, 43% feeder).
As per import protocol requirements, all feeder cattle were vaccinated against BoHV-1, and breeder cattle on voyage D were vaccinated against BoHV-1, BVDV-1, and BRSV. All vaccinations were administered as a single shot at depot entry.
Nasal swab samples
Nasal swabs were collected from live export cattle during induction processing up to 24 hr after the cattle entered the pre-export assembly depot. All nasal swab sampling was performed by a single member of the project team (SJ Moore) with the exception of sampling of slaughter cattle at depot 7 (cohort 10) that was undertaken by an Australian Quarantine and Inspection Service–accredited third-party veterinarian.
For nasal swab collection, the animal was restrained in a head bail and a 20-cm cotton swab was inserted approximately 10 cm into the nasal cavity. The swab was rotated across the nasal mucosa to collect a sample of the nasal secretions. During both insertion and removal, care was taken to prevent the swab being contaminated by dirt et cetera on the nostrils. The swab was immediately placed into a 5-ml plastic tube filled with 1–2-ml viral transport media (VTM; Hanks balanced salt solution, penicillin G [1,000 units/ml], streptomycin [25 mg/ml], and amphotericin B [0.1 mg/ml]; Department of Agriculture and Food, Western Australia) and kept chilled until transport back to the laboratory.
To investigate whether the prevalence of viral and bacterial shedding changed with time, 2 cohorts of animals were sampled 7 days (cohorts 8a and 8b) and 9 days (cohorts 12a and 12b) apart. Cohorts 8a and 8b were carryover animals from a previous shipment, and cohorts 12a and 12b were newly received animals (Table 2). It was not possible to select the same individuals for sampling at the first and second sampling sessions, but animals that were sampled at the second session were selected from pens containing animals that had been sampled at the first session. Nasal swab samples were processed, and nucleic acids from the organisms commonly associated with BRD 52 —BoCV, BoHV-1, BVDV-1, BRSV, BPIV-3, H. somni, M. bovis, M. haemolytica, P. multocida—were detected using quantitative reverse transcription polymerase chain reaction (qRT-PCR) assays as described previously. 47
Serological samples
Serological samples were collected from animals from a single voyage (voyage C; Table 1). Serum and nasal swab samples collected from the same animal at a single time-point (depot entry) were available for 334 feeder cattle: a single cohort of 91 animals from South Australia, and 3 cohorts from Western Australia, a total of 243 animals (Table 2). Cohort 8a comprised animals that had been carried over from a previous shipment. Cohorts 9, 11, and 12a were newly received animals.
Whole blood was collected at entry to the pre-export assembly depot as part of the pre-export health checks required by the importing country. While the animal was restrained in a crush, blood was collected into sterile plain Vacutainer tubes a by venipuncture of the coccygeal vein. Serum samples were sent to the Department of Agriculture and Food, Western Australia, for commercial Bluetongue virus and Bovine leukemia virus antibody testing. After serum aliquots had been taken for commercial testing, another aliquot of serum was taken for serology as part of the study reported herein.
Aliquots of 200 μl of serum were tested for the presence of circulating antibodies to the viruses of interest using commercially available enzyme-linked immunosorbent assay (ELISA) kits according to the manufacturer’s instructions for the following viruses: BoHV-1, b BRSV, c BVDV-1, d and BPIV-3. e It was not possible to test for antibodies to BoCV because a validated commercial test is not available. Samples with optical density reading above the level set by the kit manufacturer were considered positive, below negative, and in-between were inconclusive. Inconclusive samples were not retested, and animals with inconclusive test results were not included in the analysis.
Statistical analysis
Sample size estimates were based on power analyses conducted using Win Episcope 2.0 (http://www.clive.ed.ac.uk), suggesting that 20–100 samples per property of origin would be sufficient to allow description of prevalence and preliminary assessment of association between risk factors and pathogen prevalence while adjusting for property level clustering. The final approach involved sampling 20–25 animals per property and up to 4–5 properties per cohort.
The statistical software package R was used for descriptive statistics 55 and commercial software f for regression analyses. Ninety-five percent confidence intervals for descriptive prevalence estimates were generated using the Wilson approach and expressed as percentages.7,48 Overall prevalence estimates were generated as an average of the individual cohort prevalence estimates, and as an overall individual animal prevalence.
Negative binomial regression with adjustment for clustering at the property of origin (PIC) level was used to investigate associations between the apparent prevalence of viruses and bacteria of interest and available explanatory variables: voyage, sampling month, sampling year, assembly depot, assembly depot state (Western Australia, Victoria, South Australia), sex, type (Bos taurus, Bos indicus), class (feeder, slaughter, breeder), and live-weight (in kilograms). Data on animal breed was only available for a limited subset of animals and was not used in the analysis. Because of the sparseness of the data and presence of confounding between many explanatory variables, regression models were run as univariate screening models only to assess for associations between explanatory variables and apparent pathogen prevalence. Spearman rank correlation test was used to test for correlations among the presence of various organisms, as detected by qPCR, in the nasal swab samples. A threshold of P = 0.05 was used for all statistical analyses.
Results
Nasal prevalence
One or more of the viruses or bacteria of interest were detected in the nasal swabs from 1,150 out of 1,484 (77%) cattle. The remaining 334 (23%) animals were negative for all organisms tested. Pathogen prevalences for all viruses and bacteria were found to vary significantly between cohorts (P < 0.05).
Nasal pathogen prevalences for viruses and bacteria of interest can be found in Table 3. Bovine coronavirus was the most commonly detected virus (Table 3). The remaining viruses were present in cattle from 2 (BoHV-1), 3 (BVDV-1, BPIV-3), or 4 (BRSV) voyages but generally at low prevalences. Exceptions included the prevalence of BRSV for voyage D (10%) and the prevalence of BVDV-1 for 2 cohorts in voyage C (cohort 9 = 12%, cohort 11 = 10%; Table 3).
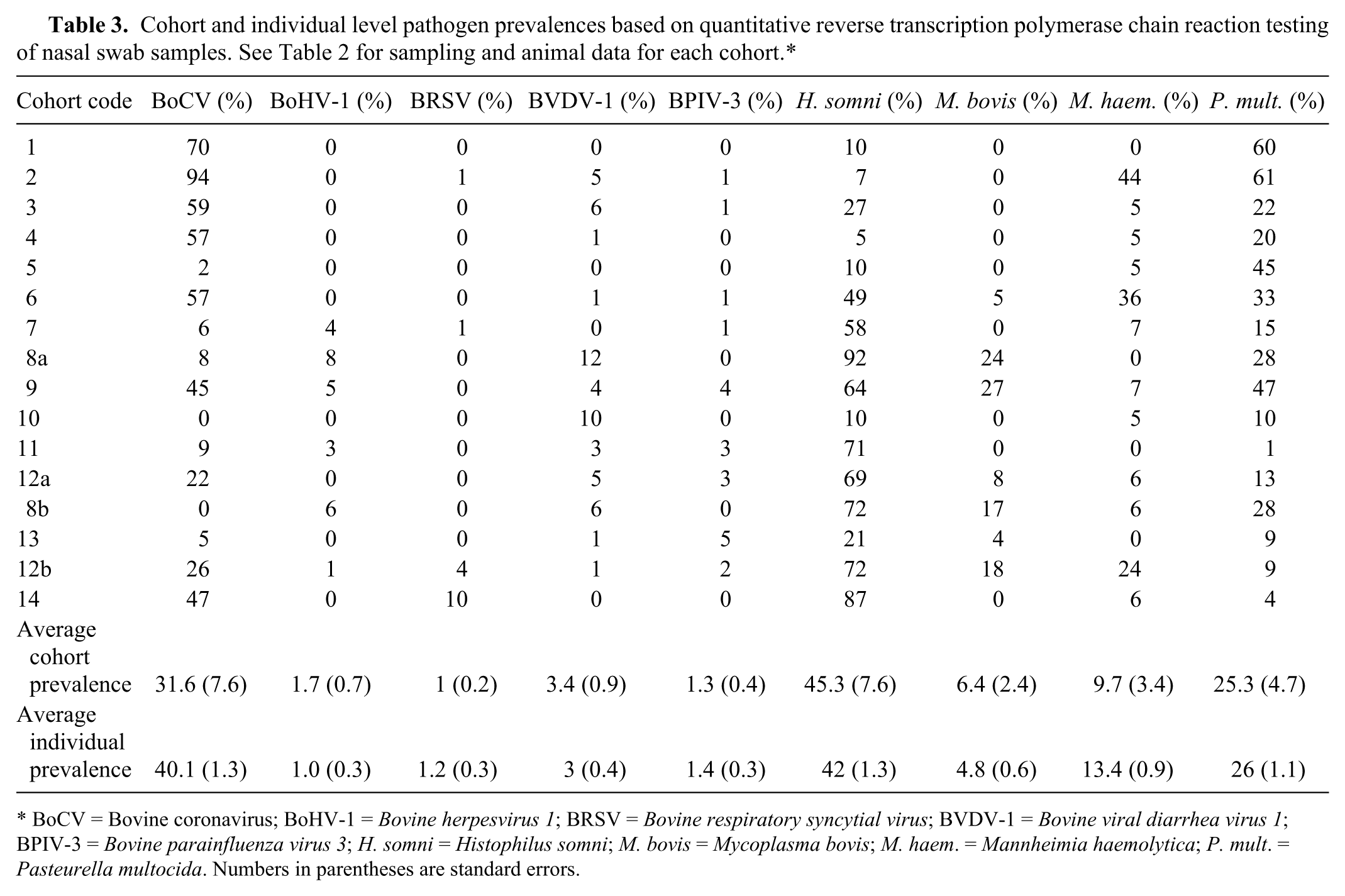
BoCV = Bovine coronavirus; BoHV-1 = Bovine herpesvirus 1; BRSV = Bovine respiratory syncytial virus; BVDV-1 = Bovine viral diarrhea virus 1; BPIV-3 = Bovine parainfluenza virus 3; H. somni = Histophilus somni; M. bovis = Mycoplasma bovis; M. haem. = Mannheimia haemolytica; P. mult. = Pasteurella multocida. Numbers in parentheses are standard errors.
Multiple organisms were detected in nasal swab samples from many animals in the current study. Overall, a single bacterium only was detected in 503 out of 1,150 (43.7%) animals, while 1 or more viruses and bacteria were detected in 434 (37.7%) animals, and 1 or more viruses in 213 (18.5%) animals. Bovine coronavirus was present in 92% of single viral detections and 92% of mixed viral–bacterial detections. The presence of BoCV was significantly correlated with the presence of BRSV, M. haemolytica, P. multocida, and H. somni (all P < 0.01). Bovine viral diarrhea virus was present in 8% of mixed infections but was not significantly correlated with any other viruses or bacteria (all P > 0.1). The presence of BRSV was significantly correlated with the presence of H. somni (P < 0.001). Bovine herpesvirus 1, BVDV-1, and BPIV-3 were not significantly correlated with any other organism.
Histophilus somni was present in 77% of single bacterial detections and 54% of mixed viral–bacterial detections. The presence of H. somni was significantly correlated with the presence of BoCV, BRSV, M. bovis (all P < 0.001), and P. multocida (P < 0.05). Pasteurella multocida was present in 33% of single bacterial detections and 51% of mixed viral–bacterial detections. The presence of P. multocida was significantly correlated with the presence of BoCV (P < 0.001), M. haemolytica (P < 0.001), and H. somni (P < 0.05).
Of the resampled cohorts, significant decreases in pathogen prevalence were observed for BoCV and H. somni in the carryover cohorts (cohorts 8a and 8b; both P < 0.01). For the newly received cohorts (12a and 12b) the prevalence of M. haemolytica (P < 0.001), BRSV (P < 0.01), and M. bovis (P < 0.05) was higher at the second testing session, and BVDV-1 was borderline significantly lower (P = 0.057).
Analysis of nasal prevalence versus explanatory variables
The total number of animals that tested positive for BoCV, BoHV-1, BRSV, BVDV-1, and BPIV-3 was 594, 15, 17, 44, and 21, respectively. For H. somni, M. bovis, M. haemolytica, and P. multocida, the total number of positive animals was 622, 72, 199, and 387, respectively.
Negative binomial regression with PIC as a fixed effect showed a number of associations between pathogen prevalence for BoCV, H. somni, M. haemolytica, and P. multocida, and available explanatory variables (Supplementary Table 1, available at http://vdi.sagepub.com/content/by/supplemental-data). Data sets for BoHV-1, BRSV, BVDV-1, BPIV-3, and M. bovis were not suitable for univariate analysis due to the low number of animals testing positive for these organisms.
There were significant differences between voyages, sampling month, and sampling season for all organisms (all P < 0.01) on which univariate analysis was performed. Sampling year was significant for M. haemolytica and P. multocida (both P < 0.01), and assembly depot for BoCV and P. multocida (both P < 0.001). Results for sampling year, sampling season, and sampling month should be interpreted with caution due to a high degree of confounding between these variables and voyage. Assembly depot state, sex, type, class, and weight were not significant for any organisms (all P > 0.1; Supplementary Table 1, available at http://vdi.sagepub.com/content/by/supplemental-data). Multivariate analyses were attempted but were not successful due to the sparsity of data and confounding between explanatory variables.
Seroprevalence
Overall, 88% of animals were seropositive for BPIV-3, 58% for BVDV-1, 50% for BRSV, and 43% for BoHV-1 (Table 4). There were significant differences in seroprevalence between PICs (all P < 0.001) and cohorts for all viruses. For newly received cattle, the average seropositive rate was 87% for BPIV-3, 56% for BVDV-1, 46% for BoHV-1, and 39% for BRSV. Carryover animals had significantly higher seropositive rates for all viruses when compared to newly received animals (P < 0.01). Of the 330 animals for which serological results were available for all 4 viruses of interest (i.e., excluding animals with inconclusive results for any virus), 12 (4%) were seronegative, and 47 (14%) were seropositive for antibodies to a single virus, 89 (27%) to 2 viruses, 130 (39%) to 3 viruses, and 52 (26%) to all 4 viruses (Table 5).
Summary data showing the number of animals returning conclusive seroprevalence test results and the seroprevalence estimates (percentage positive [mean] and 95% confidence interval) for 4 sampled cohorts each tested for presence of antibodies to 4 bovine respiratory disease viruses.*
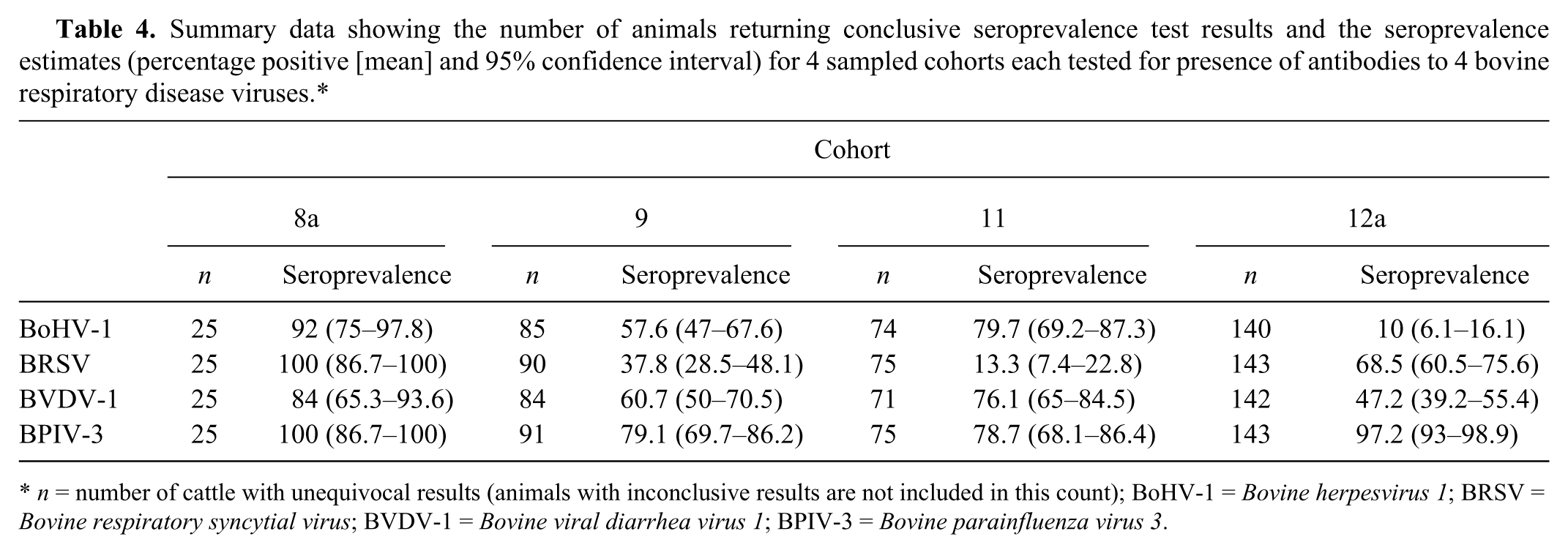
n = number of cattle with unequivocal results (animals with inconclusive results are not included in this count); BoHV-1 = Bovine herpesvirus 1; BRSV = Bovine respiratory syncytial virus; BVDV-1 = Bovine viral diarrhea virus 1; BPIV-3 = Bovine parainfluenza virus 3.
Combinations of seropositive results to single or multiple viruses. Animals with inconclusive results for 1 or more viruses are not included in this table.*
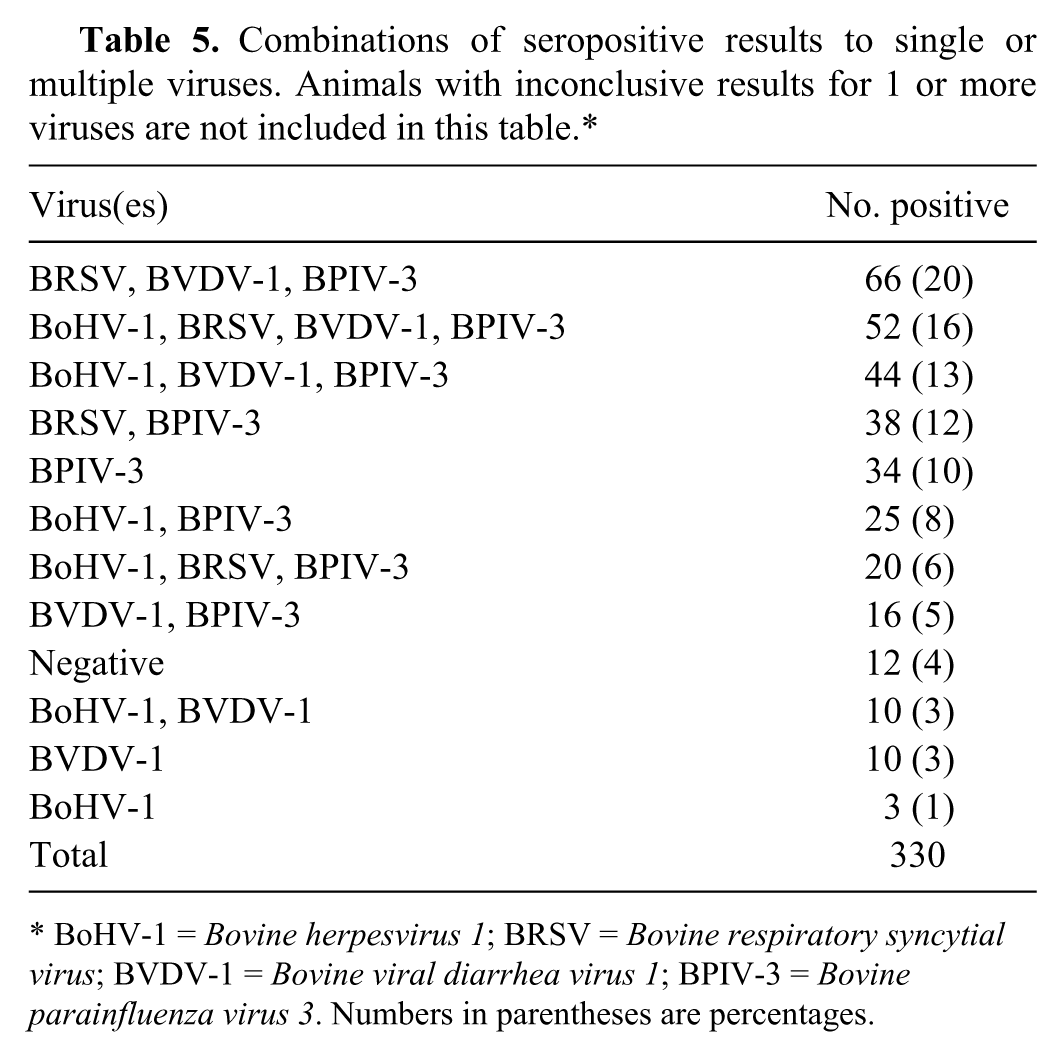
BoHV-1 = Bovine herpesvirus 1; BRSV = Bovine respiratory syncytial virus; BVDV-1 = Bovine viral diarrhea virus 1; BPIV-3 = Bovine parainfluenza virus 3. Numbers in parentheses are percentages.
The results from the matched nasal swab and serum samples for each animal were combined to determine that animal’s shedding and serological status for each virus at a single time-point. Animals were placed into 1 of 4 groups: seropositive and nasal swab negative, seronegative and nasal swab negative, seropositive and nasal swab positive, and seronegative and nasal swab positive (Table 6). For all viruses, the proportion of animals that were nasal swab negative and seropositive was significantly higher (all P < 0.01), and the proportion nasal swab negative and seronegative was significantly lower (all P < 0.05) for carryover animals compared to newly received animals.
Serological status and nasal swab results for each virus for newly received animals.*
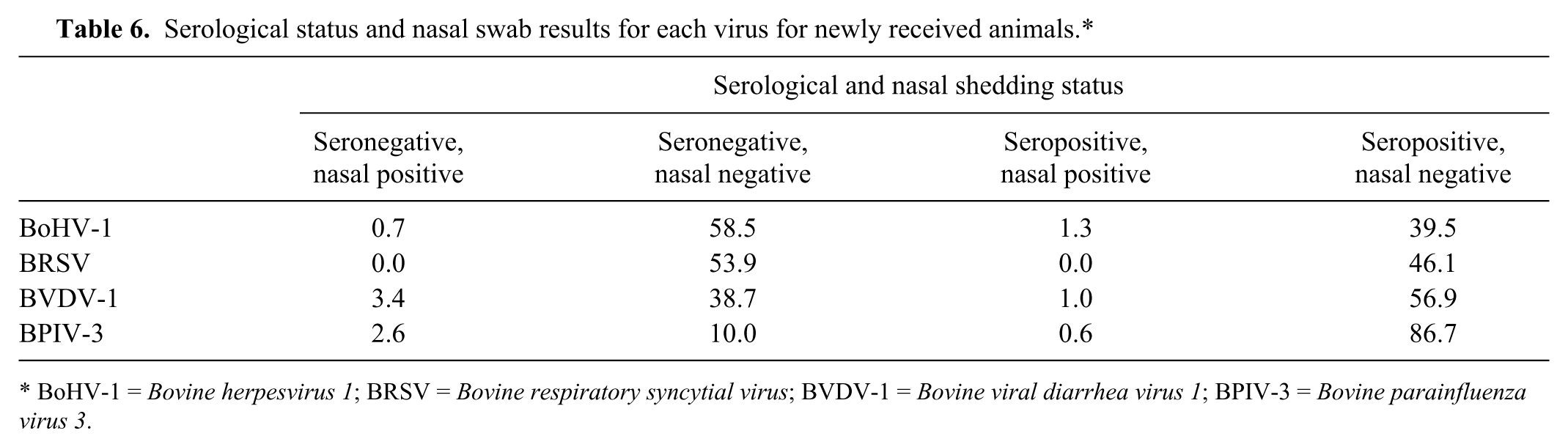
BoHV-1 = Bovine herpesvirus 1; BRSV = Bovine respiratory syncytial virus; BVDV-1 = Bovine viral diarrhea virus 1; BPIV-3 = Bovine parainfluenza virus 3.
The percentage of animals that were nasal swab positive and seronegative was higher in the carryover cohort (cohort 8a; 3/25 animals; 12%) than in the 3 cohorts of newly received cattle (cohorts 9, 11, and 12; 2.7–4.9%). Animals in cohort 8a had been carried over from a previous shipment for reasons including a positive serum BVDV-1 protein A antibody-capture ELISA (PACE) antigen test.
Discussion
The current study describes a systematic evaluation of the prevalence of organisms known to be associated with BRD, based on qPCR testing of nasal swab samples, for Australian cattle. In addition, to the authors’ knowledge, this is the first time that the seroprevalence of viruses known to be associated with BRD has been determined for Australian live export cattle, although there have been a number of previous serosurveys for these viruses in Australian cattle (Dunn SE, Godwin J, Hoare R, Kirkland PD: 2000, Diseases of feedlot cattle in eastern Australia: Meat Research Corporation project DAN.064 (1990–1994). In: Proceedings of the Australian Association of Cattle Veterinarians, pp. 76–81. Perth, Western Australia; Dunn SE, Godwin J, Hoare RJT, Kirkland PD: 1995, Diseases of feedlot cattle: Project DAN.064. Final report to the Meat Research Corporation (Australia). Meat & Livestock Australia Limited, Sydney, Australia).15,41,62–64,69,83 Comparison of the nasal and seroprevalences determined in the current study to previously published work, combined with available data on BRD-related risk factors, allows hypotheses to be made about the role of these organisms in the development of BRD in live export cattle.
The major constraints on the number of samples and range of animals (type, class, sex) that could be collected were finances, time and/or labor, and industry tolerance. Because of the large numbers of animals that must be processed and the economic incentives to minimize the length of time that the animals spend in the assembly depot prior to load-out, animals are protocolled through multiple depots on the same day. Depots may be hundreds or thousands of kilometers apart so it was not possible to sample cattle at all depots on all days.
Nasal swab samples were collected from 1,484 out of 55,902 (2.6%) cattle and matched serum samples from 334 (0.6%) cattle across the 4 study voyages. This means that, particularly in the case of serological results, the prevalences reported for sampled animals may not necessarily represent the prevalence of these organisms across the entire shipment. The collection of paired sera and calculation of seroconversion rates would have increased the power of the current study. It was not possible to access the same animals for a second sampling occasion during the assembly period, and there are no facilities on board the ship to rebleed these animals.
The individual animal prevalence of BoCV was similar to that reported previously for cattle on the south coast of New South Wales with clinical BRD, 35 and within the range of previously reported values for clinically normal beef feedlot cattle (0–85%).10,27,32,39,40,54,71 An association between nasal shedding of BoCV and clinical signs of respiratory disease has been reported in some studies32,66,67,71 but not others.10,33 The presence of BoCV in nasal secretions has been linked to increased odds of death due to BRD during live export voyages, 47 and an increased risk of treatment for respiratory disease,27,38,54 reduced weight gain, 71 and increased incidence of pulmonary lesions at slaughter 38 in feedlot cattle.
Serological testing for BoCV antibodies was not performed as part of the study reported herein due to a lack of a commercially available test kit. However, animals with BoCV in their nasal swabs tend to have low serological titers for BoCV antibodies,32,54,71 and low antibody titers at feedlot entry are associated with an increased likelihood of developing clinical respiratory disease38,53,71 and requiring treatment for respiratory disease.27,54 It is therefore likely that live export cattle that are shedding BoCV on entry to the assembly depot will have low antibody titers and will thus be more likely to develop and require treatment for respiratory disease during voyages than animals without BoCV in their nasal secretions.
To the authors’ knowledge, BVDV has not been previously detected in individual animal nasal swabs from naturally exposed cattle including beef and dairy cattle with clinical BRD,13,66 and beef calves at feedlot entry.22,24 The lack of previous reports of BVDV in nasal swabs may be unexpected considering that BVDV is a well-known BRD pathogen. However, this is likely to be a methodological rather than an epidemiological effect; most previous BVDV studies have used serology or detection of virus in blood or tissue samples to demonstrate infection, rather than PCR testing of nasal swab samples. Bovine viral diarrhea virus has been detected in nasal swabs and lung samples from animals that died during voyages but was not significantly associated with fatal respiratory disease. 47
The seroprevalence of BVDV-1 (56%) was within the range of previously reported values for Australian cattle (45–77%; Dunn SE, et al.: 1995, Diseases of feedlot cattle),15,41,64,69 and similar to the “approximately 60%” reported for Western Australian live export cattle in 1985. 41 Seroconversion during the feeding period is linked to an increased likelihood of requiring treatment for BRD. 50
In the present study, the 16 animals that were nasal swab positive and seropositive or seronegative for BVDV-1 were either transiently infected and captured immediately prior to, or soon after, mounting an immune response, or they were persistently infected (PI). Persistently infected animals have viral antigen in their nasal secretions and are mostly seronegative, although they may be seropositive if they have been exposed to a heterologous BVDV-1 strain.5,46
In feedlots, PI animals can infect 70–100% of susceptible contacts in a pen.22,25 Acute BVDV infection is associated with impaired function of cells of the innate and acquired immune systems, in particular lymphoid cells, which predisposes newly infected animals to secondary bacterial infections.21,57 The risk of initial treatment for BRD was reported to be 43% greater in cattle exposed to PI cattle than unexposed cattle, 42 although another study did not find an association between exposure to a PI animal and an increase in BRD prevalence. 51 Consideration should be given to the management of potential PI animals, particularly carryover animals, during the assembly period to minimize the risk of BVDV infection and subsequent development of BRD, bearing in mind that the application of a single PACE test to remove PI animals does not guarantee the removal of the virus from a live export consignment. 47
Bovine herpesvirus 1 prevalence was similar to that observed in cattle at feedlot entry in the United States (0–1.37%).26,66 Bovine herpesvirus 1 is known to play an important role in the development of BRD through both direct tissue damage and immunosuppressive effects that allow secondary bacterial infections to cause respiratory disease.3,37 However, BoHV-1 in nasal or lung samples collected from cattle on long-haul voyages is not significantly associated with mortality due to BRD, 47 and the presence of BoHV-1 in nasal swabs is not associated with subsequent illness or performance in American feedlot cattle. 26
Thirty-nine percent (122/309) of newly received animals were seropositive for antibodies to BoHV-1. This is higher than that found in a 1967 study on Western Australian cattle (29%) but within the range of previously reported seroprevalences for Australian beef cattle (13–85%; Dunn SE, et al.: 2000, Diseases of feedlot cattle in eastern Australia; Dunn SE, et al.: 1995, Diseases of feedlot cattle).15,64,83 The 61% of animals that were seronegative in the study reported herein were susceptible to infection by pen-mates shedding BoHV-1. However, no significant increase in BoHV-1 nasal prevalence was recorded for newly received animals sampled at feedlot entry and again 9 days later. Therefore, it is likely that BoHV-1 does not play a primary role in the development of BRD during voyages.
Bovine respiratory syncytial virus is recognized as a common primary pathogen in respiratory disease in calves less than 6 months of age6,58 and has also been reported in association with respiratory disease outbreaks in adult dairy cattle.2,17,18 Bovine respiratory syncytial virus has not previously been detected in nasal swabs from beef feedlot cattle over the age of 4 months with13,35,66,74 or without22,24 clinical signs of respiratory disease. However, BRSV was present in nasal swabs from 4 out of 616 (0.6%) 3-week-old, clinically normal, Scottish beef and dairy calves 36 and 67–100% of 2–6-month-old calves after experimental infection with BRSV. 73 In the current study, the significant increase in BRSV nasal prevalence between depot entry and re-resting 9 days later (cohorts 12a and 12b) is likely to indicate transmission of BRSV to naïve animals during this period.
The seroprevalence of BRSV in the study reported herein (46%) is higher than that reported from a serosurvey of Australian feedlot cattle (27%; Dunn SE, et al.: 1995, Diseases of feedlot cattle) and lower than that detected in cattle in an outbreak of respiratory disease in New South Wales (100%). 35 Seroconversion during the feeding period has been linked to an increased risk of treatment for BRD.8,71
The seroprevalence for BPIV-3 (87%) was the highest out of the 4 viruses. This, combined with a low nasal prevalence (1.4%) and lack of evidence for an association between BPIV-3 and respiratory disease in Australian live export cattle 47 or Australian (Dunn SE, et al.: 2000, Diseases of feedlot cattle in eastern Australia; Dunn SE, et al.: 1995, Diseases of feedlot cattle) 76 or North American 24 feedlot cattle, suggests that BPIV-3 plays a secondary role, if any, in the development of BRD in Australian live export cattle.
In all previous studies on the prevalence of bacteria in nasal swab samples, bacterial culture was used for isolation and identification of bacteria, while in the present study, culture was not performed prior to PCR detection of bacterial DNA. Bacterial culture has been found to be less sensitive than PCR for identifying bacteria of interest in lung swabs 70 and milk.23,44 Therefore, it is likely that some of the differences in prevalence observed between the current study and previous studies are due to diagnostic sensitivity and sample location.
Histophilus somni is a commensal organism of the upper respiratory tract 12 but is also commonly associated with a number of disease syndromes including fibrinopurulent pneumonia, myocarditis, and polyarthritis–serositis. 29 Previous reports of the prevalence of H. somni in nasal swabs range from 0% to 9%.1,11,24,30,61,75 There is currently no H. somni vaccination registered for use in Australia (https://portal.apvma.gov.au/pubcris) so the relatively high prevalence of H. somni reported herein (42%) is not due to detection of shedding of an attenuated vaccine strain. Nasal isolation rates at feedlot entry have not been associated with subsequent respiratory disease in feedlot cattle.1,24 Although the prevalence of H. somni in live export cattle that died during voyages was relatively high (48%), this bacteria was not significantly associated with either histological evidence of pneumonia or death due to BRD. 47
The prevalence of M. haemolytica (13%) is within the range of previously reported prevalences for M. haemolytica in nasal swabs from clinically normal feedlot cattle (2–33%).1,19,24,43,72,82 The observed increase in nasal prevalence of M. haemolytica between depot entry and resampling is likely to be due to the proliferation of commensal bacteria following inhibition of the immune system secondary to environmental stressors (e.g., transportation, comingling, and inter-animal transmission of viruses and bacteria).28,56 However, rhythmic variation in the presence or absence of M. haemolytica in the nasal secretions of healthy calves has been reported previously. 43 An association between a higher prevalence of M. haemolytica in nasal swab samples and clinical respiratory disease has been found in some studies1,72 but not others.19,24,43
The prevalence of P. multocida (26%) was within the range of previously reported prevalences for clinically normal beef cattle (6–61%).1,24,43,75 Most studies have not detected an association between nasal isolation rates of P. multocida and respiratory disease24,43,75 but a previous study 1 found that P. multocida was isolated more frequently from the nasopharynx of animals with clinical signs of respiratory disease than those without.
Mycoplasma bovis is an important cause of pneumonia, arthritis, and tenosynovitis in feedlot cattle although the role of M. bovis in the development of BRD is still not well defined. 9 Previously reported prevalences of M. bovis in apparently healthy feedlot calves range from 0% to 43%.1,31,80–82 A positive nasal swab result for M. bovis was associated with an increased odds of fever in 1 previous study, 81 but had no effect on morbidity or mortality risk in another. 31 However, evidence of a significant increase in the prevalence in M. bovis between depot entry and resampling, combined with the finding that M. bovis is significantly associated with mortality due to respiratory disease during voyages, 47 suggests that M. bovis is an important pathogen in live export cattle. However, all 4 bacteria of interest can be found as commensal organisms in the nasal cavity of apparently healthy cattle. Therefore, further work is needed to determine the link between the prevalence of bacteria in the assembly depot and risk of mortality due to BRD during voyages.
Significant associations were detected between the presence of BoCV and BRSV, M. haemolytica, P. multocida, and H. somni in nasal swab samples. A previous study 66 isolated a number of organisms—including BoCV, BRSV, M. haemolytica, and P. multocida—from 4–8-month-old beef feedlot calves with clinical BRD. Correlations between organisms are not reported, but the researchers hypothesized that bacterial infections, in particular M. haemolytica, played a role in enhancing shipping fever in the cattle in their study. 66 Co-shedding of BoCV and BRSV has not been reported previously.14,66
Of the epidemiological factors for which data were available, property of origin had the most significant effect on both nasal and seroprevalence. When differences in nasal prevalence between properties was taken into account, no significant differences were detected between animals from different states, or of different sex, type (Bos indicus vs. Bos taurus), class (feeder, slaughter, breeder), or weight categories. Commingling of cattle from multiple sources has been linked to an increased incidence of BRD in feedlot cattle.60,68 This may result from the exposure of large numbers of animals of unknown health status to a variety of viral and bacterial organisms.60,68 The majority of animals on long-haul voyages are sourced directly from their property of origin to meet import protocol requirements, so commingling at the assembly depot presents an important BRD risk.
Given that there are significant differences in nasal and seroprevalence between properties, strategies to boost immunity prior to loading (e.g., vaccination) would be more effective if they were applied at the property of origin, rather than once the animals have entered the pre-export assembly depot. However, on-farm vaccination is associated with logistic and planning implications that are not compatible with the existing live export cattle sourcing process. Therefore, while exporters could encourage producers preparing cattle for sale to the live export market to consider on-farm vaccination, the development of single shot, fast acting, efficacious vaccines that can be administered at arrival at the assembly depot may prove to be more cost effective and feasible from a management perspective.
Based on the results of the current study, BoCV would appear to be the viral pathogen of most significance for future studies into feedlot vaccination protocols. The other viruses of interest—BoHV-1, BPIV-3, BRSV, BVDV-1—were present at low prevalences in the assembly depot, and none of these viruses were significantly associated with BRD during voyages. 47 However, it is possible that these 4 viruses may be contributing to the development of BRD through their adverse effects on the respiratory tissues and immune system.52,68
The results of the present study show a mixed level of nasal prevalence and immunity to organisms associated with BRD in animals entering preexport feedlots. Therefore, the implementation of a vaccination strategy to raise antibody levels to the common viral BRD pathogens (e.g., BoCV) prior to export may mitigate the risk of development of BRD during voyages. In addition, informed selection of animals and the application of commingling strategies for animals from different properties have the potential to reduce exposure of naïve animals to BRD potential pathogens and thus reduce losses due to BRD morbidity and mortality during voyages.
Footnotes
Acknowledgements
This project would not have been possible without the cooperation of exporters, third-party Australian Quarantine and Inspection Service–accredited veterinarians, and stockmen. Thank you to Dr. Anne Barnes (Murdoch University) and Dr. John Creeper (Department of Agriculture and Food, Western Australia [DAFWA]) for editorial comments on this article. Special thanks to staff of the Bacteriology and Virology Departments at DAFWA for help with sample processing and testing.
a.
Greiner Bio-One VACUETTE TM, Greiner Bio-One, Kremsmünster, Austria.
b.
Infectious bovine rhinotracheitis (IBR) gB X2 Ab test, IDEXX Laboratories Pty. Ltd., Rydalmere, New South Wales, Australia.
c.
Bovine respiratory syncytial virus (BRSV) IgG antibody test kit, IDEXX Laboratories Pty. Ltd., Rydalmere, New South Wales, Australia.
d.
Bovine viral diarrhoea virus (BVDV) antibody test kit, IDEXX Laboratories Pty. Ltd., Rydalmere, New South Wales, Australia.
e.
Parainfluenza-3 virus (PI3) antibody test kit, IDEXX Montpellier SAS, Montpellier, France.
f.
Stata version 13, StataCorp LP, College Station, TX.
Declaration of conflicting interests
The author(s) declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
Funding for this project was provided by Meat & Livestock Australia and LiveCorp (project no. W.LIV.0252).