
Abstract
We used a real-time PCR (rtPCR) targeting a 150-bp amplicon of the mitochondrial small subunit of ribosomal RNA (mtSSU rRNA) to screen for Pneumocystis DNA in lungs of wild squirrels (Callosciurus finlaysonii, n = 85) and river rats (Myocastor coypus, n = 43) in Italy. The rtPCR revealed Pneumocystis DNA in 20 of 85 (24%) squirrels and in 35 of 43 (81%) river rats, and was more sensitive than a nested PCR that targets a portion of the mtSSU rRNA and the mitochondrial large subunit of rRNA (mtLSU rRNA). Phylogenetic analysis based on mtSSU rRNA and mtLSU rRNA sequences showed distinct Pneumocystis sequence types in these rodents. The rtPCR assay should be reliable for screening large populations for this potential pathogen, thereby allowing cost-effective monitoring of the disease in wild animals.
Introduction
Genus Pneumocystis fungi are typically restricted to the lungs. A wide range of mammals can be parasitized by this potential pathogen, with each mammalian host studied infected by a species-specific Pneumocystis, sp. 1 , 16 , 24 Pneumocystis spp. are not zoonotic organisms. They cannot exist as saprophytes in nature without their mammalian host, have adapted to a lifecycle exclusively within lungs, and over time have evolved to live inside a great variety of terrestrial and aerial mammals. 2 They do not grow outside the lung on artificial media.6,15 Phylogenetic analysis based on gene sequences provides the basis for species and sequence-type distinctions within the genus. 3 Although numerous Pneumocystis species are likely to exist within mammals, only 5 have been formally described: P. carinii and P. wakefieldiae in rats,7,8 P. murina in mice, 18 P. oryctolagi in rabbits, 10 and P. jirovecii in humans. 27
Pneumocystis organisms are transmitted from host to host via aerosols. They may cause severe pneumonia, often lethal if untreated, in immunologically impaired domestic mammals. 9 In contrast, in wildlife, Pneumocystis spp. are commonly reported in apparently healthy individuals,8,13,16,19,24 suggesting that animals are frequently colonized rather than infected. 5 The significance of Pneumocystis colonization in the clinical context 25 is acquiring relevance given that modern studies have demonstrated that the Pneumocystis genome has evolved to permit colonization in immunocompetent hosts.2,22
Because Pneumocystis cannot grow on synthetic media in vitro, various detection methods, including immunohistochemical stains (conducted since the 1970s), and recently more sensitive sequence-based typing, have been applied to a variety of specimens, including bronchoalveolar lavage (BAL) liquid, sputum, oral swabs, and lung tissue from mammals.4,8,16,19,20,32 Understanding and harmonizing detection methods is pivotal to compare epidemiologic data among the numerous animal species living in different environments and countries. Among the existing molecular assays, nested PCR (nPCR) assays targeting the mitochondrial large and small subunit of ribosomal RNA (mtLSU rRNA and mtSSU rRNA, respectively) are the most common assays used to investigate Pneumocystis in wildlife.4,8,16,19,20,32
We assessed the efficiency of a SYBR Green–based, real-time PCR (rtPCR) targeting a 150-bp amplicon of the mtSSU rRNA 9 to screen for Pneumocystis DNA in lungs of 2 groups of free-living wild rodents (squirrel, Callosciurus finlaysonii; river rat, synonyms coypu, nutria, Myocastor coypus). The performance of the screening assay was compared with the existing nPCR, which targets a portion of mtSSU rRNA and mtLSU rRNA.
Materials and methods
Lungs of 85 squirrels and 43 river rats were screened for Pneumocystis DNA at the Mycology Unit of the Parasitology Laboratory, Istituto Zooprofilattico Sperimentale delle Venezie (Italy). Squirrels were from the Basilicata region (southern Italy) 17 and river rats from the Veneto region (northern Italy), where they were trapped and euthanized according to a project for the control of allochthonous wildlife populations and approved by the “Istituto Superiore per la Protezione e la Ricerca Ambientale” (12/08/2009, 75 AG-153998). At autopsy, lungs were removed and stored at −20°C in single plastic tubes (50 mL) until tested. A sample of lung tissue (0.3 g), placed into a 2-mL tube containing 800 µL of phosphate-buffered saline (PBS) and 1 glass bead (3 mm diameter), was lysed (Tissue lyser, Qiagen, Hilden, Germany) for 2 min at 30 Hz. The 0.3-g specimen represents up to 10% and 3% of the weight of the right lung in squirrels and river rats, respectively.
DNA extraction was performed (DNeasy blood & tissue kit, Qiagen) following the manufacturer’s instructions with minor modifications. From each tube, 100 µL of the homogenized sample was transferred into a 1.5-mL tube with 200 µL of buffer ATL (lysis buffer) and 20 µL of proteinase K, and incubated overnight at 56°C. DNA was eluted in 100 µL of elution buffer. A negative control was systematically included in each series of DNA extractions.
Lung tissues (n = 128) were tested for Pneumocystis DNA by using real-time PCR (rtPCR) targeting a 150-bp fragment of mtSSU rRNA, as reported previously, 9 with minor modifications. For each reaction (20-µL final volume), 10 µL of 2× QuantiFast SYBR Green master mix (Qiagen, Valencia, CA), 3 µL of DNA, and 0.2 µM of each primer (PneuSSU189-PneuSSU362) were used. Thermal cycling conditions were as follows: denaturation and activation 1 cycle of 95°C for 5 min, followed by 40 cycles of 95°C for 15 s, annealing and extension at 60°C for 1 min (StepOnePlus real-time PCR system, Applied Biosystems, Foster City, CA). Mitochondrial SSU rRNA and mtLSU rRNA of the mitochondrial genome were amplified by using nested mtLSU rRNA and mtSSU rRNA PCR with primers and conditions described previously. 8
Because the nested mtLSU rRNA PCR did not amplify Pneumocystis DNA from squirrels or river rats, a new set of primers was designed to target a 210–230-bp fragment of mtLSU rRNA, included between the internal primers of the nested mtLSU rRNA PCR. 8 The new primers PneumoLSU 74-94 (forward: 5’-AGGATATAGCTGGTTTTCTGC-3’) and PneumoLSU 325-308 (reverse: 5’-TRTTCTGGGCTGTTYCCC-3’) were designed manually, according to general primer design rules, 14 based on a sequence alignment of reference sequences from different Pneumocystis spp. available from the NCBI database (http://www.ncbi.nlm.nih.gov). The sequence specificity of the primers used was confirmed in a BLASTn 23 search against GenBank. Amplification conditions were as described above for rtPCR screening except for using an annealing and extension at 58°C for 30s and 60°C for 30s, instead of 60°C for 1 min (StepOnePlus real-time PCR system, Applied Biosystems). Following amplification, melting curve analysis was performed to discriminate between specific amplicons and nonspecific amplification products. Negative (sterile water) and positive controls (Pneumocystis DNA derived from Sorex araneus) were included in each PCR run, to monitor for contamination and correctness of the amplification. Amplification of 18S rRNA was used as an internal control to monitor for DNA inhibition. 26 Samples were considered positive for Pneumocystis when a PCR product of the expected size was amplified and sequenced.
Sequencing reactions were performed from both ends as reported previously. 8 Sequence alignment was performed using the ClustalW algorithm, integrated in MEGA6.1, 28 which was subsequently refined manually. Only sequences obtained using the nested mtSSU rRNA PCR and the mtLSU rRNA rtPCR were used for phylogenetic trees. Molecular phylogeny was performed using the maximum likelihood (ML) approach on mtLSU rRNA and mtSSU rRNA datasets separately, and on the concatenated dataset. Tamura-3 parameter and G distribution was determined as the best model in MEGA6.1. We also added sequences to our dataset from P. murina and P. wakefieldiae, as well as other Pneumocystis sequences available from GenBank, and isolated from other rodents. Sequences of Pneumocystis isolated from Rousettus aegyptiacus (order Chiroptera, family Pteropodidae) were used as an outgroup (Table 1).
GenBank accessions of Pneumocystis sequences used for phylogeny.

Dash (—) = sequence not available; mtLSU rRNA, mtSSU rRNA = mitochondrial large and small subunit of ribosomal RNA, respectively. Sequences produced in the current study are reported in boldface.
Agreement among the results of PCR protocols was tested using kappa coefficient (k). 31 Differences among prevalence were tested using the chi-square test or Fisher exact test, when appropriate. The software used was WinEpi (http://www.winepi.net).
Results
Pneumocystis DNA was detected in 55 of 128 (43%) rodents by using rtPCR (Table 2). Although a single melting point of 75 ± 0.5°C (for squirrels) and 74 ± 0.5°C (for coypus) was generated without formation of important primer–dimer interferences (Fig. 1), the specimen was considered positive only when Pneumocystis identity was confirmed by sequencing.
Number of positive animals and prevalence of Pneumocystis in lung tissue of squirrels and coypus according to different PCR protocols.
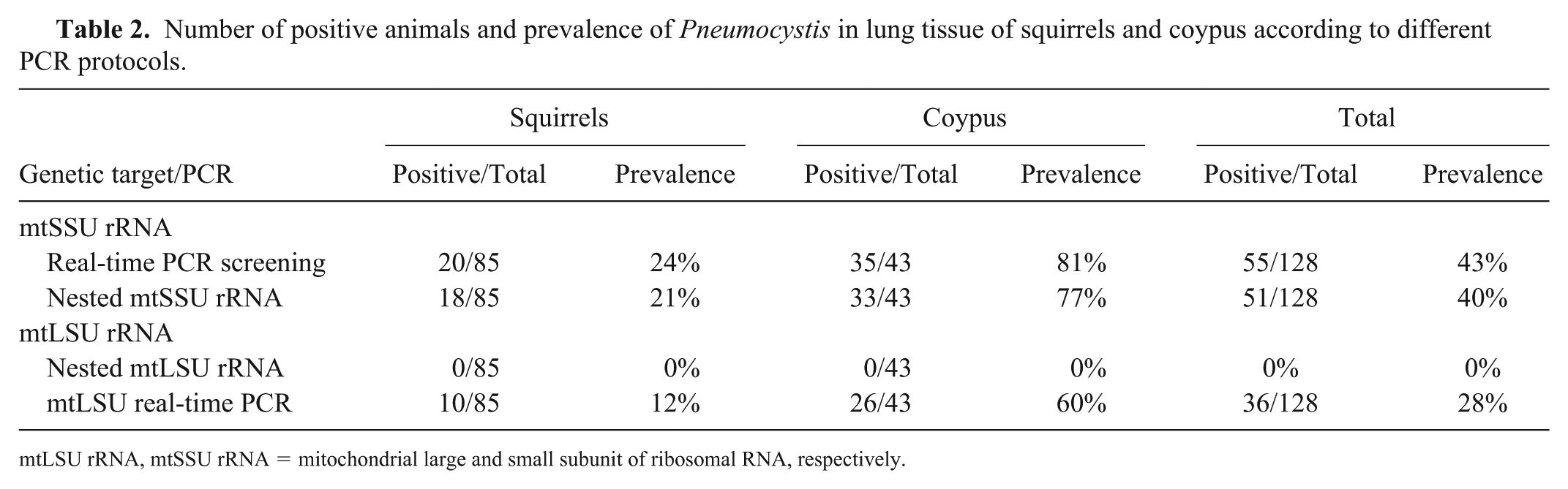
mtLSU rRNA, mtSSU rRNA = mitochondrial large and small subunit of ribosomal RNA, respectively.
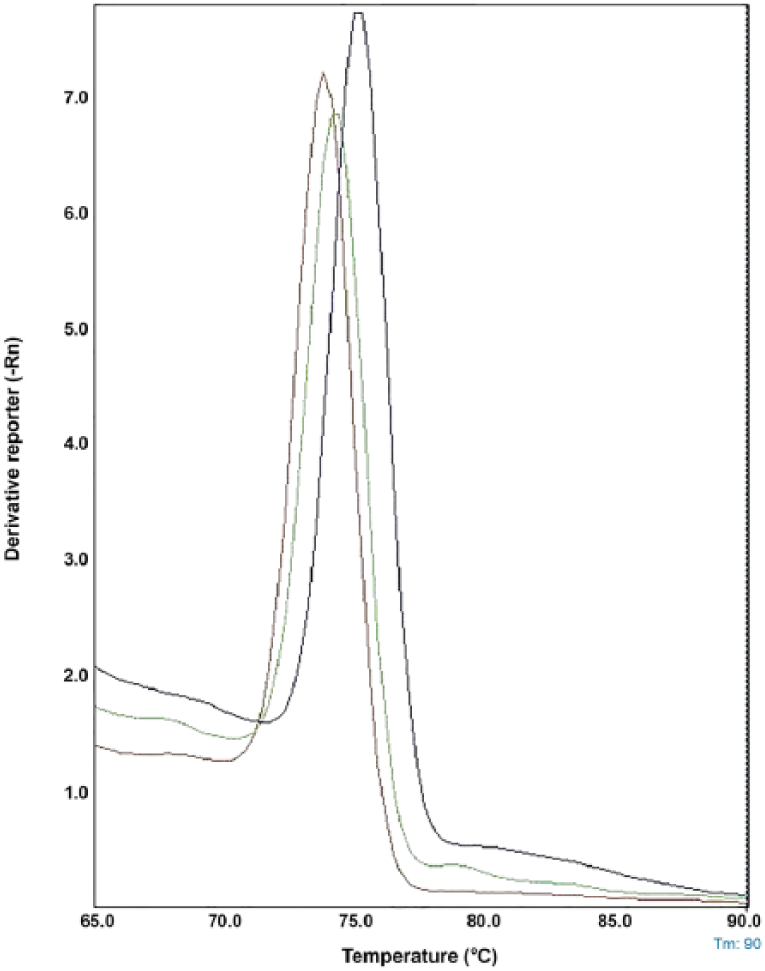
Melting curve analysis for Pneumocystis templates amplified with primers targeting part of the mitochondrial small subunit of ribosomal RNA (mtSSU rRNA) gene. Peaks correspond to Pneumocystis DNA from Sorex araneus (orange, positive control), Myocastor coypus (green), and Callosciurus finlaysonii (blue).
River rats (35 of 43; 81%) were colonized more commonly (p < 0.01) than were squirrels (20 of 85; 24%). Similarly, differences between the groups were confirmed by nested mtSSU rRNA PCR in 77% and 21% of river rats and squirrels, respectively (p < 0.01).
Overall, among the mitochondrial genes, amplification targeting mtSSU rRNA performed better (40–43%) than mtLSU rRNA (0–28%; Table 2). Interestingly, using the mtLSU rRNA gene target, Pneumocystis DNA was amplified exclusively with mtLSU rtPCR, using the newly designed primers. No amplifications were obtained using the nested mtLSU rRNA PCR protocol.
Agreement between rtPCR screening (mtSSU rRNA) and nested mtSSU rRNA PCR was excellent (k = 0.932), and was good (k = 0.665) with the mtLSU rRNA rtPCR.
Sequences of all amplicons confirmed the Pneumocystis identity, with 2 different sequence types according to the animal host. Considering the mtSSU rRNA genetic target, sequences were found to be identical among squirrels as well as among river rats, with a single mtSSU rRNA sequence type for each species. Similarly, mtLSU rRNA sequences showed a single sequence type for each of the 2 animal hosts. Representative sequences of mtSSU rRNA and mtLSU rRNA from squirrels and river rats were deposited in GenBank (Table 1). In the obtained phylogenetic trees, which were built using separate mtSSU rRNA and mtLSU rRNA sequence datasets, Pneumocystis spp. from squirrels and river rats grouped in 2 separate clades according to the animal host (Supplementary Figs. 1, 2).
Pneumocystis from the squirrels clustered with Pneumocystis from Sciurus aestuans, the only squirrel sequence in the database. Pneumocystis sequences from the 2 squirrel species (C. finlaysonii and S. aestuans) differed by 2.8% in mtSSU rRNA and by 13.7% in mtLSU rRNA. The mtLSU rRNA sequence from M. coypus was identical to the KR704345 sequence in GenBank, obtained in our previous study. 8 In the concatenated tree mtSSU/mtLSU rRNA, Pneumocystis sequences from squirrels and river rats confirmed groups in separate clades. Callosciurus and Myocastor sequences are also distinct and well separated from the Pneumocystis sequence type of other rodents (Fig. 2).
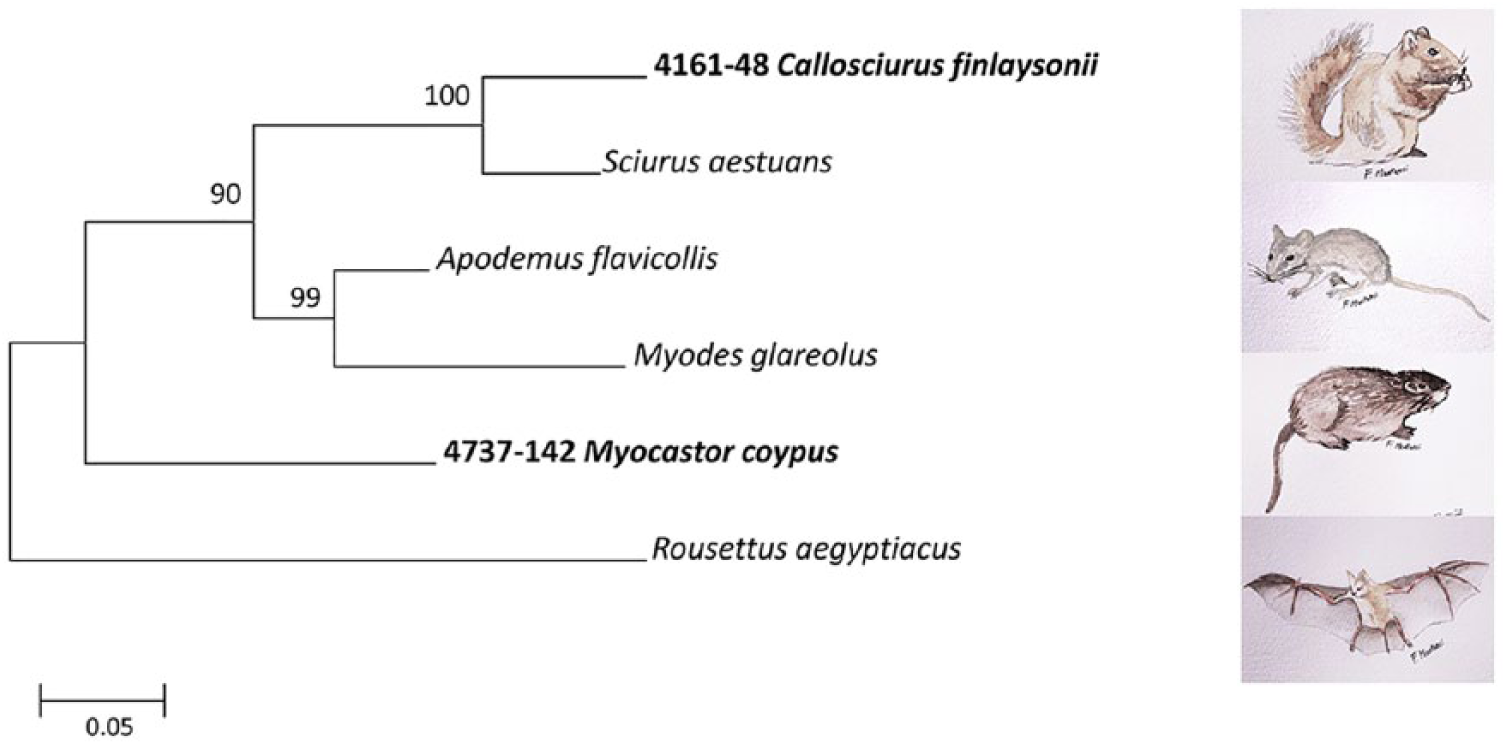
Phylogenetic maximum likelihood tree of concatenated Pneumocystis sequences of mitochondrial large and small subunit of ribosomal RNA (mtLSU/mtSSU rRNA). Tamura-3 parameter was used as best model. Pneumocystis sequences from our study are reported in bold. Pneumocystis sequences derived from different wild rodent species were compared and reported beside each branch. Pneumocystis derived from bat (Rousettus aegyptiacus) is used as outgroup. Bootstrap values (>80) are shown at the internal nodes.
Discussion
Our rtPCR assay proved to be an efficient tool for the detection of Pneumocystis in 2 wildlife species. The high sensitivity of this assay combines the use of a set of primers targeting a 150-bp segment of the mtSSU rRNA gene with SYBR Green–based detection in rtPCR. This protocol amplified Pneumocystis DNA exclusively (confirmed by sequence analysis), both in squirrels and in river rats, showing single peaks at similar but distinct melting temperatures according to the rodent species. This screening method presents a number of advantages: 1) it is fast to perform; 2) the method is cost-effective once used routinely; and 3) the amplified product can be sequenced directly. This rtPCR was not used for a quantitative purpose, but used only to detect “Pneumocystis-positive” individuals among animals in the group. In order to detect all Pneumocystis species, especially in wildlife in which novel species or sequence types are expected, we chose a “sequence-based” screening method, which is not as specific as probe-based methods, which label only specific DNA molecules. 21
We found that river rats were more likely to harbor Pneumocystis than squirrels, with a high prevalence (81%) similar to that reported for shrews. 20 The hypothesis that higher airborne transmission of Pneumocystis particles occurs when animals live in close contact with each other 11 links transmission to specific behavioral traits or phases of an animal’s life. Adult squirrels live mostly as single individuals or as a mating pair, with lower opportunity for disease transmission compared to river rats, which usually organize in small matriarchal harems. Vertical transmission of Pneumocystis, as reported for rabbits, has not been reported in rodents and might represent an efficient strategy for Pneumocystis to survive and propagate.
Pneumocystis organisms are commonly reported in apparently healthy animals, often in association with other low-grade respiratory pathogens.3,11,20,24 Unfortunately, we were not able to prepare tissues for cytology or histology from lungs to compare with the rtPCR data, because lungs were frozen upon arrival at our laboratory. The only additional information about the animal health status concerns “coinfection” of 2 of 20 squirrels harboring Pneumocystis, with Cryptococcus neoformans var. grubii and C. neoformans var. neoformans cultured from the lung and nasal cavity, respectively. 17 We do not possess data concerning the immunocompetence of rodents included in our study, but we assumed that in nature, clinically diseased animals are disabled and therefore more likely to be killed by predators or starvation. Therefore, trapped animals are more likely to be healthy.
In testing animals captured in nature, we expect low and subclinical loads of Pneumocystis. In order to reduce “false-negative results”, we used at least 3% (river rats) and 10% (squirrels) of the total lung volume, as suggested by previous studies. 30 We also tested our squirrels with nested mtSSU rRNA PCR using only 0.025 mg of lung; amplified Pneumocystis DNA could be amplified from only 5 of 85 (6%; data not shown) compared to 18 of 85 (21%) obtained in our study.
Real-time PCR was more sensitive than nested mtSSU rRNA PCR, amplifying Pneumocystis DNA in 55 versus 50 hosts, respectively. This is probably the result of the smaller size of the amplicons (50 vs. 320–340 bp), overcoming possible Pneumocystis DNA degradation.
In our study, mtSSU rRNA was a more efficient target for detecting Pneumocystis DNA than mtLSU rRNA. This may be explained by reports that the copy number of mitochondrial genes in P. jirovecii varies from gene to gene, with mtSSU rRNA being more common than others. 29 Indeed, in the case of a very low fungal load, mtSSU rRNA is constantly present, with a median ratio of 2.5 compared to mtLSU rRNA. 29 That would also explain why several authors were not able to amplify both mitochondrial SSU and LSU rRNA genes in animal Pneumocystis studies.8,12
Supplemental Material
DS1_JVDI_10.1177_1040638718797379 – Supplemental material for Real-time PCR assay for screening Pneumocystis in free-living wild squirrels and river rats in Italy
Supplemental material, DS1_JVDI_10.1177_1040638718797379 for Real-time PCR assay for screening Pneumocystis in free-living wild squirrels and river rats in Italy by Patrizia Danesi, Christian Falcaro, Silvia Ravagnan, Graziana Da Rold, Elena Porcellato, Michela Corrò, Roberta Iatta, Claudia Cafarchia, Antonio Frangipane di Regalbono, Wieland Meyer and Gioia Capelli in Journal of Veterinary Diagnostic Investigation
Supplemental Material
DS2_JVDI_10.1177_1040638718797379 – Supplemental material for Real-time PCR assay for screening Pneumocystis in free-living wild squirrels and river rats in Italy
Supplemental material, DS2_JVDI_10.1177_1040638718797379 for Real-time PCR assay for screening Pneumocystis in free-living wild squirrels and river rats in Italy by Patrizia Danesi, Christian Falcaro, Silvia Ravagnan, Graziana Da Rold, Elena Porcellato, Michela Corrò, Roberta Iatta, Claudia Cafarchia, Antonio Frangipane di Regalbono, Wieland Meyer and Gioia Capelli in Journal of Veterinary Diagnostic Investigation
Footnotes
Acknowledgements
We thank Dr. Fabrizio Montarsi for animal drawings used in phylogenetic trees, and Fabiano D’Este for figure editing.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
Funding was provided by the Italian Ministry of Health (project code RC IZSVe 01/2015).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.