
Abstract
Vesicular stomatitis (VS) is a vesicular disease of horses, cattle, and pigs in the Western Hemisphere caused by viruses in the genus Vesiculovirus. Disease manifests as vesicles and erosions on the oral mucosa, teats, prepuce, and coronary band, and is similar in presentation to foot-and-mouth disease. Laboratory confirmation is therefore required. Conventional assays include competitive (c)ELISA and complement fixation (CF). The cELISA provides more accurate herd-level detection of VSV-exposed cattle, but may lack the ability to capture fluctuating antibody levels in individual animals. The CF assay can confirm newly infected animals because of its ability to detect antigen–antibody complexes, thus is considered to be indicative of IgM. We evaluated the immune status of 2 herds affected by VSV in 2014 by testing sera collected in June 2015. Two conventional assays were compared to a novel IgM-IgG ELISA. When sampled in 2015, both herds had detectable VSV-specific antibodies; 18% and 36% of animals tested by cELISA and 2% and 8% of animals tested by CF were positive. The novel IgM-IgG assay exhibited fair agreement (adjusted kappa score of 48) with the conventional assays, and should be evaluated further to assess its ability to replace the 2 separate assays with a single assay system, or for its ability to replace the CF assay as a more sensitive method for defining newly exposed animals.
Introduction
Vesicular stomatitis (VS), a vesicular disease of horses, cattle, and pigs in the Western Hemisphere, is caused by vesicular stomatitis virus (VSV; genus Vesiculovirus, family Rhabdoviridae). Two species of vesiculovirus cause VS in the Americas: New Jersey vesiculovirus (NJVS) and Indiana vesiculovirus (INDVS).9,12 INDVS is comprised of 3 subtypes: Indiana 1 (VSIV1), Indiana 2 (VSIV2; also known as Cocal), and Indiana 3 (VSIV3; also known as Alagoas).9,11 Vesiculoviruses have a single-stranded, linear, negative-sense RNA genome that is prone to a high error rate in replication given inefficient functionality of the associated RNA polymerase, which results in replication errors in the viral genome. 13
Clinical disease manifests as a vesicular disease with both vesicular and erosive lesions of the oral cavity, teats, prepuce, and coronary band of affected animals. The severity of disease can vary. Transmission occurs via direct contact with infected animals that have vesicular lesions, and through insect vectors. Sandflies show evidence of being a biological vector of VSV and exhibit transovarial transmission of the virus; other vectors, such as black flies, eye gnats, and mosquitos, are mechanical vectors. 9
Laboratory detection of VSV infection in affected cattle and pigs occurs at a national reference laboratory because the disease is clinically indistinguishable from foot-and-mouth disease. Trade limitations are associated with VSV, requiring suspect cases to be ruled out or confirmed. Morbidity in a herd can vary widely; subclinical disease can occur in 90% of animals with 10% of animals showing clinical signs. 11 Mortality in cattle and horses is negligible. 11 Current laboratory techniques used routinely at the U.S. Department of Agriculture (USDA) Foreign Animal Disease Diagnostic Laboratory (FADDL; Greenport, NY) include reverse-transcription real-time (RT-rt)PCR, antibody competitive enzyme-linked immunosorbent assay (cELISA), and a complement fixation (CF) technique.
Of the 5 genes encoding VSV, both the surface glycoprotein (G) and nucleocapsid (N) proteins are highly immunogenic in infected animals.1,5,21 The cELISA utilizes a recombinant N antigen for detection; whereas a newly developed double-sandwich IgM-IgG ELISA utilizes an inactivated whole antigen that includes the G and N antigens. Sera of recently infected cattle contain high titers of virus-neutralizing antibodies induced by the G protein; the N protein induces a non-neutralizing antibody. 1 The cELISA has proven to be highly sensitive as a screening tool and will detect early and long-lasting antibodies, 7 but cannot distinguish between IgM and IgG. The CF test is a specific method used to confirm recent antibody–antigen interactions (presence of IgM); however, the CF test lacks sensitivity, can be laborious, cannot be used on anti-complementary or hemolyzed sera, and requires live virus. 19 Therefore, we are interested in replacing the CF assay with a more sensitive assay specific for IgM.
Serologic responses have been long-lasting in endemic countries of the Western Hemisphere.9,13 Complement-fixing IgM titers are reported to fall to nondetectable levels 2–4 mo post-exposure. 9 Yet, IgM responses may recur given antigenic stimulation associated with re-exposure of the immune system of animals as the result of viral RNA persistence in animals or re-exposure from a reservoir, 12 although the model for this is not well understood and this does not seem to result in viral replication. 15 Seropositive animals are not protected from re-infection. 12
In the first part of our study, we assessed the immune status of cattle herds >6 mo after they tested positive for VSV during the 2014 VS outbreak in Colorado. Immune status was re-assessed using traditional tests prior to the onset of disease diagnosis in 2015. The purpose was to understand the immune status of U.S. cattle herds that previously had animals diagnosed with VS based on the presence of clinical signs and laboratory assay results, with the consideration that VSV is not enzootic in the United States. In the second part of our study, we compared the performance of conventional assays to a novel IgM-IgG ELISA that is being validated at FADDL.
Materials and methods
Sample collection and handling
In late June 2015, blood samples from 2 cattle herds in Colorado containing cows that were diagnosed as infected with NJVS in the fall of 2014 were collected into serum separator tubes and sent overnight on cold packs to FADDL. Five animals in these herds had clinical signs of disease in the fall of 2014, and were confirmed to be infected with NJVS through laboratory testing by FADDL at that time. In case investigations of disease outbreaks, only affected animals were sampled, so only 2 of the individual cattle tested in 2014 were available for retest in 2015. In 2014, 1 herd had 2 cows that tested positive, 1 animal by detection of viral RNA via RT-rtPCR, with negative cELISA results, and 1 animal that tested positive by RT-rtPCR. The other herd had one cow with positive cELISA and CF results and negative RT-rtPCR results, and another animal that tested positive by RT-rtPCR. Two animals that were positive either via serology or RT-rtPCR (one from each farm) in the 2014 outbreak were also retested in 2015. The other 2015 follow-up blood samples were convenience samples of farms affected in the 2014 outbreak and whose owners agreed to participate in the study. All animals that were available from the herd were sampled in June 2015. In total, 108 samples from 2 farms were collected, 44 from farm 1 and 64 from farm 2.
The sample collection procedure was approved by the Plum Island Animal Disease Center’s Institutional Animal Care and Use Committee. Upon receipt, all samples were evaluated and entered into the local laboratory information management software for tracking. Samples were centrifuged at 554 × g for 10 min at 4°C (Jouan GR4a centrifuge, Thermo Scientific, Waltham, MA) and were stored at 4°C until tested. The initial testing was done within 1 mo of receipt (CF and cELISA), the virus neutralization (VN) was done in June 2016, and the novel ELISA was tested throughout the year, as time was available.
Each sample was tested using the NJVS antibody cELISA and CF assay. VN was completed on some samples, including the 2 animals retested between 2014 and 2015 to evaluate current antibody titers compared to 2014 test results. In addition, a newly developed double-sandwich VSV ELISA was used to evaluate 48 samples for IgM and IgG antibodies from all cELISA-positive and some -negative samples. Typically, the cELISA is used to screen for VSV infection by detecting all circulating VSV antibodies, and if positive, the CF assay is used to confirm recent and likely active infection based on the classical complement pathway, indicating the detection of IgM antibodies. The novel ELISA evaluates samples for the presence of both IgM and IgG antibodies against VSV individually.
Conventional assays
cELISA
The cELISA, which utilizes a recombinant nucleocapsid antigen and a polyclonal antibody, was used to test blood samples for the presence of antibodies, which aligns with procedures outlined by the World Organization for Animal Health (OIE).6,7,18 NUNC PolySorp 96-well plates (Thermo Scientific) were coated with 75 µL/well of NJVS recombinant antigen diluted in 0.05 M carbonate–bicarbonate, incubated at 4°C overnight, and subsequently blocked with phosphate-buffered saline (PBS) and 5% nonfat dry milk (NFDM) at room temperature for 30 min. Plates were washed 3 times with PBS and 0.05% Tween 20 (PBST), with additional wash steps following subsequent incubations with NJVS ascitic fluid and conjugate. The diluent used to prepare the serum, ascitic fluid, and conjugate dilutions was PBS and 1% NFDM, and the volume added per well for these reagents was 50 µL with an incubation at 37°C for 30 min. Test and control sera were added in duplicate along with a diluent blank consisting of PBS and 1% NFDM. Immediately following the serum incubation, polyclonal anti-NJVS ascitic fluid was added to all wells. An anti-mouse IgG conjugated to horseradish peroxidase was then added to all wells. Following the final wash step, substrate was added to all wells, and plates were allowed to incubate at room temperature for up to 15 min. TMB (3,3’,5,5’-tetramethylbenzidine) stop solution was then added to all wells. An ELISA reader set at a wavelength of 450 nm was used to determine optical density (OD) values, and mean OD values were calculated. Percent inhibition (PI) was calculated for all samples by dividing the mean OD of the sample by the mean OD of the blank, subtracting the resulting decimal from 1.0, and multiplying by 100%. PI ≥ 50% was considered positive (Berninger ML. Competitive enzyme-linked immunosorbent assay (cELISA) of serodiagnosis of vesicular stomatitis (VS) virus (New Jersey and Indiana-1 strains). [SOP-DS-0010.2; FADDL], 2015).
CF assay
A modified CF test was used to detect early antibodies against VSV.2,18 The test differs from the OIE-prescribed terrestrial diagnostic manual in that it uses bovine serum rather than rabbit serum, and uses mouse brain–derived antigen. Test, positive control, and negative control sera were diluted 1:5 in veronal-buffered saline (VBS), heat inactivated at 56°C for 30 min to destroy any complement in the sera, and 8 serial 2-fold dilutions were prepared in 96-well “U” bottom plates. NJVS and VSIV1 antigens were produced from brains of mice inoculated with NJVS and VSIV1. The normal antigen was also produced from mouse brains. Antigens diluted in VBS, along with complement diluted in VBS and 5% calf serum (modifying factor), was added to the plates to increase test sensitivity in cattle and swine. Appropriate controls for the serum, antigens, complement, and sheep red blood cells (RBCs) were included in the assay. Plates were initially incubated at 37°C for 3 h. A suspension of 2.8% sheep RBCs was diluted 1:2 in hemolysin (rabbit anti-sheep RBC antibody) for a final dilution of 1.4%. The 1.4% solution was incubated at 37°C for 15 min and then added to all plates. Final incubation occurred at 37°C for 30 min. The plates were centrifuged at low speed to visualize the presence or absence of hemolysis. A positive reaction occurred when there was 0% hemolysis. The endpoint titer was the last dilution exhibiting 0% hemolysis (Berninger ML. Complement fixation test for detection of antibodies to vesicular stomatitis virus. [SOP-DS-0011.02; FADDL], 2016).
VN assay
The VN assay used is a modification from the test described in the OIE terrestrial diagnostic manual in that it does not use the Vero cell line, has a more robust viral titer range, and includes an inconclusive titer range. 18 VN was done to determine the neutralizing antibody titers in serum samples.14,17,18 Sera were heat inactivated at 56°C for 30 min and serial 2-fold dilutions prepared; the virus was diluted to contain 794–2,512 TCID50/25 µL (a viral concentration of 102.9–103.4). The VN assay consists of an initial incubation of equal volumes (25 µL/well) of serum and virus, for a total volume of 50 µL/well, at 37°C for 60 min, followed by the addition of 150 µL/well of Mengling-Vaughn porcine kidney (MVPK) cells from a 2 × 105 cells/mL suspension. Following a 3-d incubation at 37°C, the presence or absence of a cytopathic effect was determined. Controls in the assay included positive and negative control sera, back titration of the working stock of virus to verify the appropriate virus concentration, and a cell control. The Spearman–Kärber method was used to determine 50% endpoint titers for the serum and virus. 4 Serum titers <1:8 are negative, titers ≥1:8 and <1:32 are inconclusive, and titers ≥1:32 are positive (Berninger ML. A virus neutralization test for the detection of antibodies to vesicular stomatitis virus (VSV), serotypes New Jersey and Indiana (subtypes 1, 2, and 3). [SOP-DS-0009.02; FADDL], 2015).
Novel assay
Double-sandwich IgM-IgG ELISA
A newly developed double-sandwich IgM-IgG ELISA is being validated by FADDL and was evaluated in our study for agreement against the conventional cELISA and the CF assay. Affinity-purified polyclonal antibodies to bovine IgM heavy chain (µ) and IgG, heavy and light chains (H+L), were coated overnight at 4°C on Immulon 96-well plates (Thermo Scientific). Plates were blocked with 5% NFDM and washed with PBST. Additional reagents were diluted in 1% NFDM, and incubations occurred for 45–60 min at room temperature, with 3 washes between steps. Bovine field and control sera were added to each of the IgM- and IgG-coated wells, followed by the addition of NJVS binary ethylenimine-inactivated (BEI) antigen. Next, mouse-produced monoclonal NJVS antibody was added and detected with anti-mouse IgG conjugated to horseradish peroxidase. Finally, TMB peroxidase substrate was added and, after ~2 min, TMB stop solution was used. Plates were read on a spectrophotometer at 450 nm, and the OD of the sample was divided by the negative control to obtain the signal-to-noise (S:N) ratio. Samples with S:N >2.2 were considered positive for IgM antibody; S/N >1.2 was considered positive for IgG.
Statistical analysis
Herd immune status
Summary statistics were calculated to describe the general demographic characteristics of the 2 farms. Frequency, seroprevalence, and 95% confidence intervals (95% CIs) were calculated at the farm level and the overall sample size level based on the designation of each assay as positive or negative for each assay type, including the novel assay system for detection of both IgM and IgG antibody.
Test comparison
The kappa statistic was calculated to assess the agreement beyond chance between the conventional assays and the IgM-IgG ELISA for both of its potential antibody isotypes. The prevalence and bias indices and the corrected kappa were also calculated. 3 A kappa measure of <0 indicates less than chance agreement, 1–20 = slight agreement, 21–40 = fair agreement, 41–60 = moderate agreement, 61–80 = substantial agreement, and 81–99 = almost perfect agreement. 20 The results were captured and analyzed using Excel (Microsoft, Redmond, WA).
Results
Immune status
Two farms in Colorado were sampled in 2015, one dairy and one beef. Only cows were sampled on both farms, and no other domestic animal species were present on the farms. The average age on the beef farm and dairy farm was 6.1 y and 4 y, respectively. The beef farm had 137 cows that were all pregnant and lactating (Table 1). The dairy farm had 4,000 head of cattle. The farm sizes had not changed since the fall of 2014, when the initial VSV diagnosis was made. An initial VSV diagnosis was made in November 2014 for farm 1 and September 2014 for farm 2; clinical disease was seen on both farms at the time of laboratory confirmation.
Demographic summary statistics of the sampled Colorado cattle farms, June 2015.

Numbers in parentheses are standard deviations.
Using the cELISA, the seroprevalence of persisting antibodies in farm 1 cattle was 18% and in farm 2 cattle was 36%. Using the CF assay to measure recent exposure, seroprevalences were 2% and 8%, respectively. The results indicate the presence of early antibodies in 7 of the animals, and samples were collected prior to the identification of positive herds or clinical signs of disease in these areas at that time (Table 2).
Summary statistics and kappa comparison of serologic tests used to measure vesicular stomatitis virus IgM and IgG antibodies.
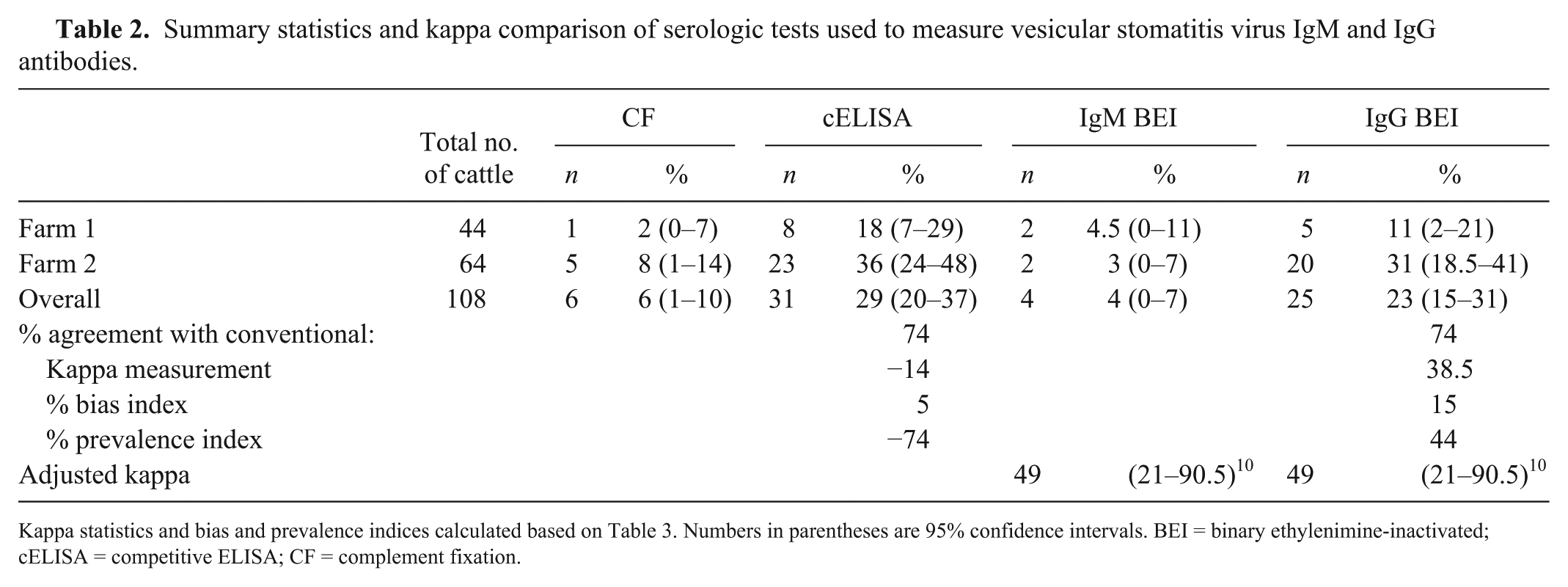
Kappa statistics and bias and prevalence indices calculated based on Table 3. Numbers in parentheses are 95% confidence intervals. BEI = binary ethylenimine-inactivated; cELISA = competitive ELISA; CF = complement fixation.
The sample of farm 1 captured 32% of the herd population; 8 of 44 (18%) animals were seropositive by cELISA, suggesting a history of inapparent infections. The sample size of farm 2 was small compared to the herd size (2%); however, 23 of 64 (36%) cattle analyzed were positive by cELISA.
Two cattle from farm 1 had clinical signs of disease (erosions and drooling) when tested in November 2014. A swab of dental pad erosions was submitted from one of the animals, and the swab was positive for NJVS by RT-rtPCR. Serum submitted from the other cow was positive for VSV antibodies on both the cELISA and CF in 2014; the CF titer was 1:80. The RT-rtPCR test conducted on a probang oropharyngeal sample from this cow was negative. The June 2015 blood samples for both cattle were negative on cELISA and CF. The samples were then evaluated via VN, and the 2 samples had neutralizing antibody titers of 1:45 and 1:260. The latter sample was the one with paired serology from the fall of 2014 in which both the cELISA and CF assays were positive via conventional testing.
Test comparison
There was 74% agreement between both the IgM-IgG ELISA and the original cELISA and the CF assay. The kappa values were −14 and 38.5 prior to adjusting for bias and prevalence, respectively. Once a prevalence -and bias-adjusted kappa (PABAK) was calculated, it reflected moderate agreement 20 between the traditional and the IgM-IgG ELISA (PABAK = 49; Tables 2, 3). A larger sample size would improve the precision of the CI and would reveal if there were differences in the assay’s abilities to detect antibodies.
Concordance of traditional versus novel assay results for the detection of vesicular stomatitis virus IgM and IgG antibodies using a novel antigen.
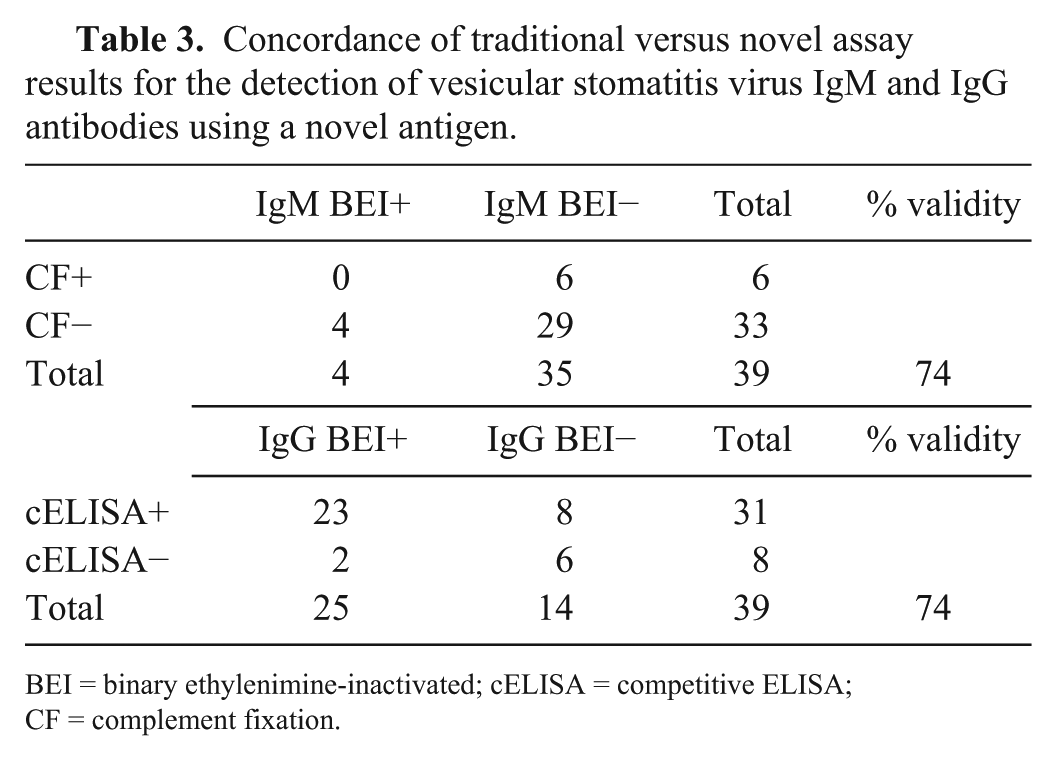
BEI = binary ethylenimine-inactivated; cELISA = competitive ELISA; CF = complement fixation.
Discussion
Collection of samples did not occur until late June 2015, and the 2015 VSV outbreak did not begin until July. Farm 1 animals were sampled on arrival in Larimer County, CO in June 2015. The animals had been in Elbert County at the time of disease diagnosis in the fall 2014 outbreak. Larimer County had its first 2015 case diagnosed July 15–17 (Pelzel-McCluskey A. 2015 vesicular stomatitis virus situation report. Fort Collins, CO: SPRS USDA, 7/17/2015. Available at: https://goo.gl/XnxfPN), and Elbert County did not have its first 2015 case diagnosed until late October 2015 (Pelzel-McCluskey A. 10/21/2015, https://goo.gl/FFA464). Farm 2 was also sampled in June 2015 in Morgan County, CO. Cases were not identified in Morgan County until October 2015 (Pelzel-McCluskey A. 10/14/2015, https://goo.gl/mnMjL8), and disease was originally diagnosed on this farm in September 2014 (Fig. 1).
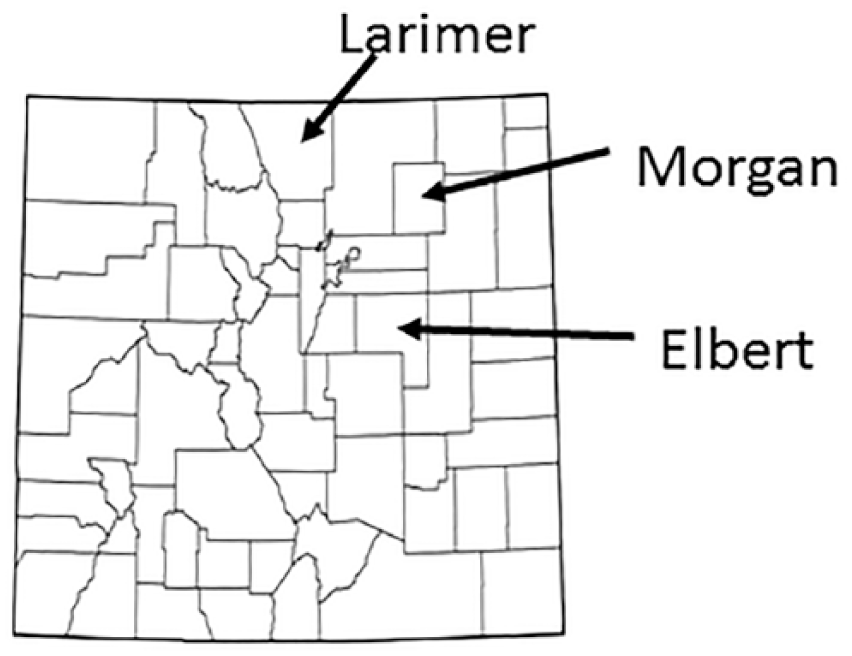
Map of Colorado counties with Larimer, Morgan, and Elbert counties identified.
At the time of sampling, it had been 7 mo since the diagnosis on farm 1, and many animals remained seropositive. Interestingly, the 2 animals that had clinical disease in fall 2014 were seronegative by cELISA when retested in 2015, but maintained a titer when evaluated by VN. This is true even for the animal that was originally found positive by CF and cELISA in the fall of 2014. This may be associated with the analytical sensitivity of the cELISA and fluctuating antibody titers, but it is imperative that a larger sample size be used to evaluate this hypothesis.
VSV-seropositive animals are known to have fluctuating IgG titers,8,9,16 and the IgG titers of the samples in our study may have fallen below the threshold of the cELISA. It is possible that, as titers persist and rise, if tested again, these animals would be positive on the cELISA, or due to the non-clinical nature of the animals, the levels of persistent antibodies were low. A single test may not be reliable for identifying historically seropositive animals, although herd seroprevalence clearly persists, with an estimated 18% of the farm 1 herd being seropositive.
The presence of early antibodies detected by CF and IgM antibodies detected by the novel ELISA indicates potential re-exposure to VSV, or persistence of IgM. This may be the result of re-exposure associated with viral RNA fragments maintained in lymphoid tissue post-infection 8 or the development of VSV quasispecies in the infected animal. 9 Animals in 1 county broke with disease 2–3 wk after sample collection in 2015, whereas the animals in the other county did not develop disease for >3 mo. Therefore, the early antibody and IgM levels potentially could indicate an upcoming outbreak. There may be potential serosurveillance capability for early detection of an outbreak, but the practical application of such surveillance in areas where epizootics occur regularly would need to be validated and the reliability of the surveillance confirmed.
In evaluating the novel double-sandwich IgM-IgG assay for VSV, agreement of 49% was considered fair overall, and the 95% CIs for the novel assay provided surety of some agreement overall. It is not clear whether cELISA or CF is a better assay than the novel double-sandwich IgM-IgG assay at this point, but it is clear there is discordance between the results that should be further elucidated. The novel assay’s detection of IgM antibodies did not concur with any CF detection of early antibodies, given that it was negative for IgM when CF was positive for 6 samples and that CF was negative when the novel ELISA was positive for the detection of IgM in 4 samples. However, it is important to note that CF-positive samples had titers ≤1:10. These animals would not be considered positive in an outbreak-free zone unless the CF titers were ≥1:40. Additionally, the only 2 samples that were tested by both IgM-IgG and VN that were found positive by IgM were also positive by VN, with the highest titers found overall (1:2,050 and 1:510, respectively); a third sample with a VN titer of 1:510 was also positive for IgG. There was greater concordance among the IgG results with the cELISA; the overall agreement was still 74%, the same as with the IgM results compared to CF (Table 3). Further analysis with a larger sample size should be used to compare these assays more robustly.
Samples were collected from herds in June 2015 that had known disease during the 2014 VSV outbreak. The 2015 collection was done prior to VSV detection in the counties of the sampled animals. In VSV-infected herds, many animals will seroconvert, and few will develop clinical signs of disease. 11 Thus, one would expect that the results would reveal long-term (IgG) antibodies from these herds in the 2015 samples. Of the 108 animals tested, 31 (29%) were seropositive on the cELISA for non-neutralizing antibodies, which are likely IgG. Interestingly, neither of the 2 cattle that exhibited a clinical and serologic response in the previous season was positive on the conventional tests 1 y later, and they were only detected to have an antibody titer based on the VN (these samples were not analyzed by the IgM-IgG ELISA). This is supportive of the concept of fluctuating antibody titers seen in endemic areas, such as Central America, 12 which are thought to play a greater role in endemic areas than in epidemic areas, such as the United States. Yet, our study provides suggestive evidence that fluctuating titers are occurring in U.S. cattle herds. Serial titers would have to be drawn from known-positive animals over a period of time to fully elucidate these findings. Nonetheless, it is clear that a large proportion of cattle in infected herds maintained an antibody response, and that seroprevalence persists at a herd level.
The kappa statistic estimates the expected agreement between measurements beyond chance alone. However, kappa measurements can be impacted by the prevalence of the effect being measured, as was seen here. Further, the kappa statistic can be biased, as seen when the difference in the discordant cells of the 2-by-2 table comparisons are not equal to one another. 3 It was necessary to adjust the kappa for prevalence and bias in the analysis. The agreement of the IgM-IgG ELISA with the conventional assays revealed correlation and could be pursued to replace the cELISA and CF assays to allow for improved testing.
Footnotes
Acknowledgements
We thank the participating farmers and producers. We thank Dr. Luis Rodriguez of the Foreign Animal Disease Research Unit of the Agricultural Research Service and Sabrina Swenson of the National Veterinary Services Laboratories of the Animal Plant Health Inspection Service of the U.S. Department of Agriculture for their input into the results and findings.
Authors’ note
Additional materials and information may be made available upon request.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
Funding was provided via the fiscal year 2015 Veterinary Service’s Grassroots micro-grant.