
Abstract
Neuroaxonal dystrophy (NAD) is a neurologic disorder of sheep characterized by accumulation of numerous axonal swellings (spheroids) in specific regions of the brainstem and spinal cord. Disruption of axonal transport, which is driven in anterograde and retrograde directions by the molecular motors, kinesin and dynein, respectively, is believed to contribute to spheroid development. Accordingly, we examined spheroids in ovine NAD cases immunohistochemically for kinesin and dynein and found both motor proteins, with dynein more strongly expressed than kinesin. Further investigations of the kinesin and dynein content of axonal spheroids in NAD, and other neurodegenerative disorders of domestic animals, could assist in better understanding the pathogenesis of these diseases.
Neuroaxonal dystrophy (NAD) denotes the occurrence of swollen axons (spheroids) in the nervous system. In humans, NAD has been subdivided into 3 types: physiologic, primary, and secondary. 8 Physiologic NAD occurs as part of the normal ageing process and is a common finding in certain regions of the brain, especially gracilis and cuneate nuclei. 8 Similarly, in all domestic animal species, numerous axonal spheroids are found in these relay nuclei with increasing age. There are usually no clinical signs in animals, 3 although physiologic NAD may account for some of the neurologic decline seen with ageing in humans. 8 In primary NAD, axonal spheroids are the principal pathologic feature, and their neuroanatomical distribution is concordant with the neurologic deficit. 8 Primary NAD has been reported in humans (as infantile, juvenile, and adult forms), sheep, horses, dogs, and cats. 13 Secondary NAD is a reactive process to another neurologic or non-neurologic disease, manifest as either a marked accentuation of physiologic NAD or widely distributed axonal spheroids in the brain contributing to the disease symptomatology. 8 It is in the context of primary NAD that the term is used in our paper.
A previous study of naturally occurring ovine NAD 9 found accumulation of markers of axonal transport—amyloid precursor protein, ubiquitin, neurofilament proteins, and synaptophysin—in axonal spheroids and suggested that disruption of this transport mechanism could contribute to spheroid development. In an attempt to further characterize the nature of the signature spheroids in ovine NAD, the spheroids were immunostained for the molecular motors of bidirectional axonal transport, kinesin and dynein.
Axons are often very long, and a variety of intracellular cargoes such as proteins and organelles required by axons and synaptic terminals for their normal function must be transported down the axon after synthesis in the neuronal cell body. Transport in the reverse direction from the periphery to the neuronal perikarya is also necessary to degrade misfolded proteins, clear protein aggregates, and provide a signaling mechanism from the synapse to the cell body. Long-distance transport in axons, both anterograde and retrograde, is driven by molecular motors of the kinesin and dynein families, respectively, that move along tracks of cytoskeletal filaments, termed microtubules. Any disturbance of axonal transport may have severe consequences for neuronal function and survival. 2
We collected brains from four 8-wk-old Merino 9 and two 2-wk-old Merino-Border-Leicester × Polled Dorset 5 lambs with clinical signs of ataxia, postural anomalies, paresis, paralysis, eventual permanent recumbency, and visual impairment. The brains were immersion-fixed in 10% buffered formalin, and coronal sections of prosencephalon, cerebellum, brainstem, and spinal cord were cut at 5-mm intervals and paraffin-embedded; 6-µm sections were then cut and stained with hematoxylin and eosin (H&E).
For immunohistochemical detection of motor proteins, rabbit polyclonal antibodies to dynein (Santa Cruz Biotechnology, Santa Cruz, CA) and kinesin (Thermo Scientific, Rockford, IL) were used in a standard streptavidin–biotinylated immunoperoxidase technique. The epitope recognized by the former antibody was amino acids 4320–4644 of the dynein heavy chain of rat origin; the latter antibody detects total levels of endogenous kinesin protein, the immunogen being the stalk domain of human kinesin expressed in (aa 331–906) Escherichia coli. In order to exclude the possibility that some of the nerve fiber swellings were dendritic rather than axonal, microtubule-associated protein (MAP-2) immunohistochemistry was performed with a monoclonal antibody to MAP-2 (Sigma-Aldrich, St. Louis, MO). In brief, sections were dewaxed using xylene and rehydrated through alcohols. Antigen retrieval was performed using EDTA (pH 8). Slides were allowed to cool and washed twice in phosphate-buffered saline (PBS; pH 7.4), then endogenous peroxidase activity was quenched. Nonspecific proteins were blocked using normal horse serum for 20 min. Dynein was applied at 1 in 4,000, kinesin at 1 in 3,200, and MAP-2 at 1 in 1,000 at room temperature overnight. The following day, the sections were given 2 washes in PBS and then a biotinylated anti-rabbit secondary (Vector Laboratories, Burlingame, CA) was applied to both dynein and kinesin for 60 min at room temperature. Following 2 PBS washes, the slides were incubated for 1 h at room temperature with a streptavidin–conjugated peroxidase tertiary (Pierce Biotechnology, Pasadena, CA). Sections were visualized using 3,3’-diaminobenzidine tetrahydrochloride (DAB), washed, counterstained with hematoxylin, dehydrated, cleared, and mounted on glass slides. Control slides were stained in parallel for all of the antibodies used. These controls included positive controls with known lesions (diffuse axonal injury in a 6-wk-old Merino lamb resulting from traumatic brain injury produced by a humane stunner; Fig. 1), minus primary antibody controls, and isotype antibody controls for the mouse monoclonal antibodies. In normal brain sections from an age-matched, clinically normal lamb, no axonal spheroids immunopositive for these motor proteins were found. Accordingly, these antibodies to kinesin and dynein have been validated for diagnostic use in the brains of sheep.
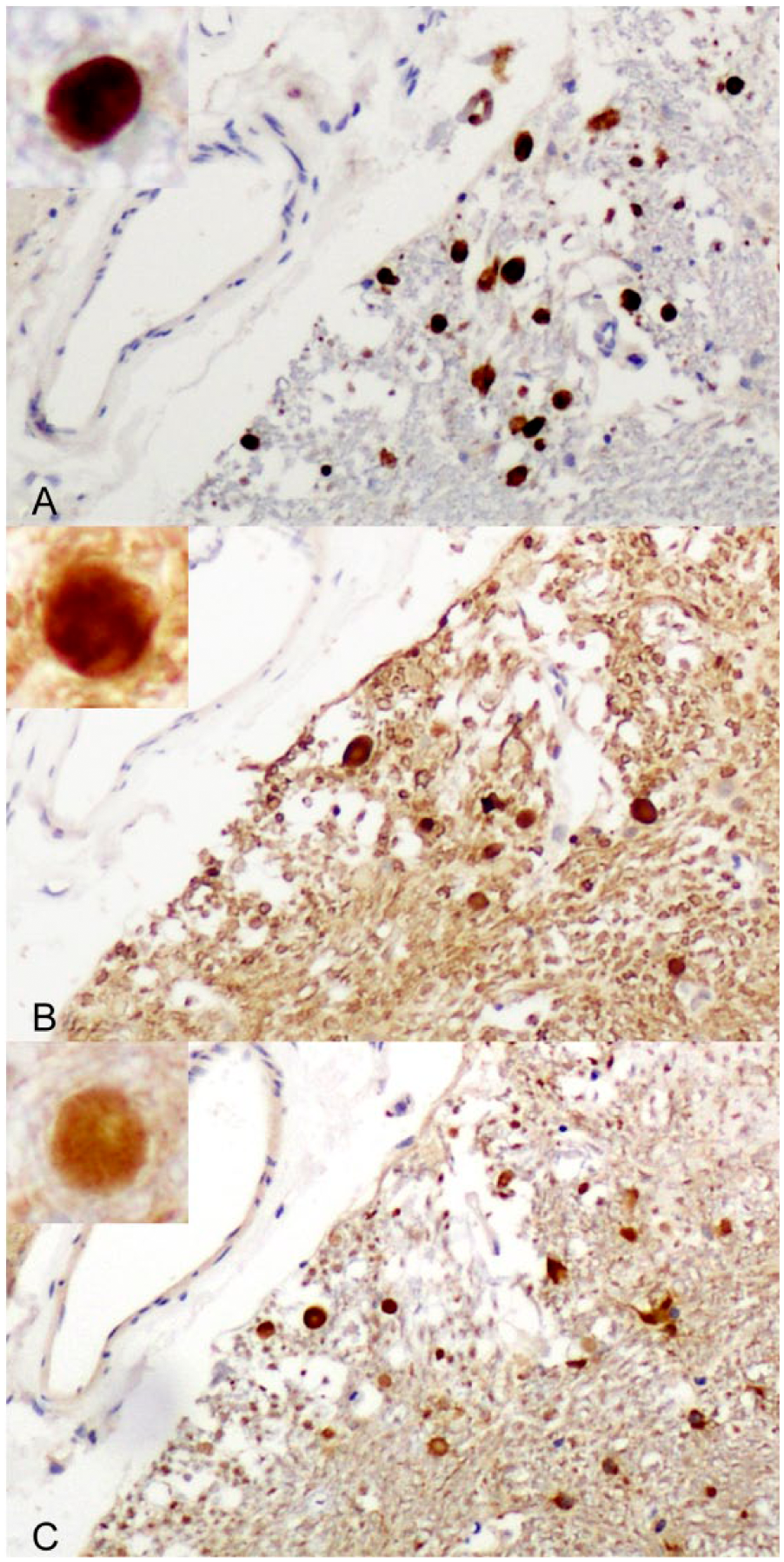
Positive control brain from a 6-wk-old, traumatically head-injured sheep showing axonal spheroids in the external cuneate nucleus immunostained for amyloid precursor protein (
Axonal spheroids were observed at all levels of the cerebrum, cerebellum, brainstem, and spinal cord, but were more frequently seen in gray matter, with nuclear aggregates bilaterally symmetrical in distribution. The spheroids were most abundant in cuneate and gracile nuclei, but were also found in the oculomotor nucleus, roof nuclei of the cerebellar peduncles, lateral geniculate nucleus, and, less commonly, in other neuroanatomic sites involved in proprioception and vision such as the lateral geniculate body, anterior colliculus, and adjacent portions of the optic tract.
In H&E-stained sections, axonal swellings (spheroids) were variable in appearance and ranged from 4–50 µm diameter. Most were round-to-ovoid and, where a spheroid was in continuity with a non-distended portion of axon, a “tail” was evident. Spheroids varied in staining intensity from strongly to palely eosinophilic; some showed a more deeply stained central core or, less commonly, the periphery was more deeply staining than the center. Some spheroids contained nonstained clefts or vacuoles. MAP-2 immunoreaction was not found at spheroid bodies.
In brain regions in which axonal spheroids were abundant, such as medullary gracile and cuneate nuclei (cuneate immunoreactivity is shown in Figs. 2, 3), there was strong dynein immunopositivity of most spheroids, the majority being uniformly immunolabeled. However, as in H&E-stained sections, some had a more deeply immunostained core (Fig. 2A); a few had stronger peripheral staining or were, occasionally, immunonegative. Some dynein-immunoreactive spheroids exhibited many nonstaining clefts (Fig. 2A) or were multivacuolated or multiloculated. Kinesin immunoreactivity of axonal spheroids was consistently much weaker than dynein, most frequently evident as peripheral immunostaining with a poorly stained central region (Fig. 3B). Kinesin immunonegative spheroids were more common than those immunostained for dynein. Immunohistochemical staining of spheroids for both motor proteins was distinguished from neuronal staining by reference to duplicate H&E-stained sections of the same region and by morphologic characteristics such as size and shape. In the former, nonstaining clefts and deeper staining central cores were evident and, in the latter, there was a nonstaining nucleus. Nevertheless, a number of neurons were weakly immunopositive to both motor proteins. This pattern of immunoreactivity was found in the brains of all 6 lambs examined and in all neuroanatomic sites where axonal spheroids had aggregated. The variable immunohistochemical staining of axonal swellings in the ovine NAD cases included in our study might suggest that they were at different stages of temporal evolution and/or of different protein composition and resembled the morphology of spheroids in H&E-stained sections.
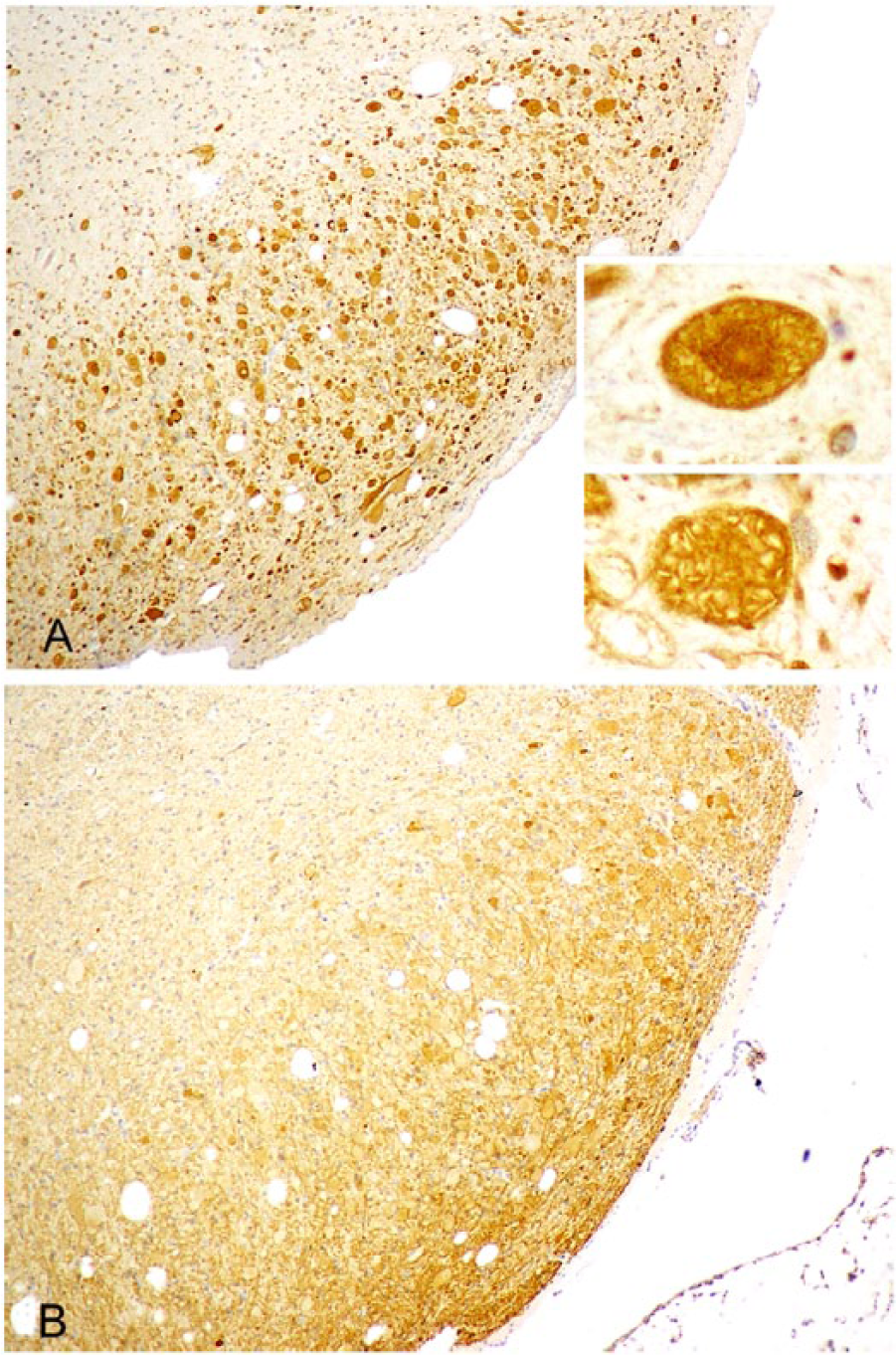
Right cuneate nucleus of an 8-wk-old Merino lamb.
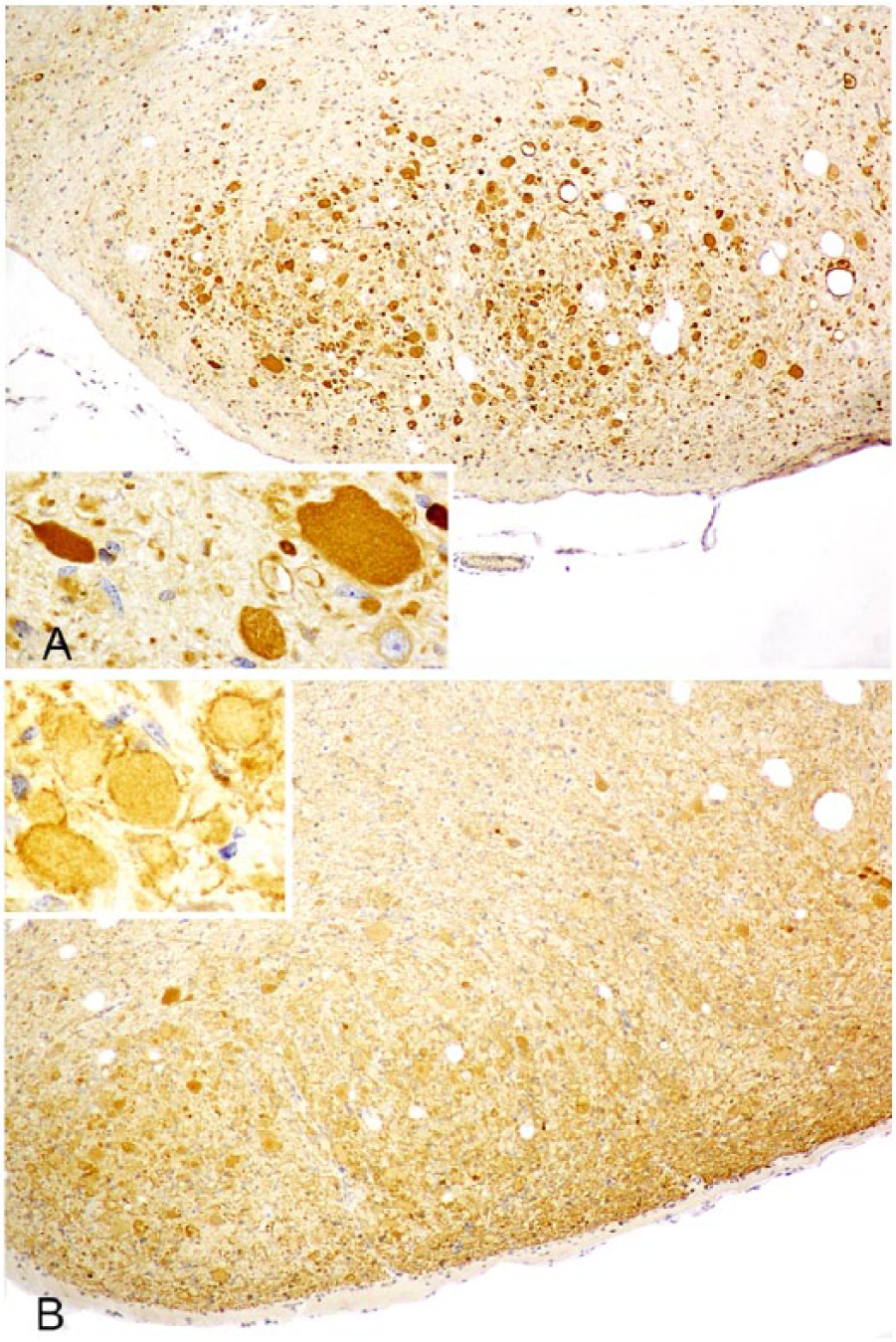
Left cuneate nucleus of an 8-wk-old Merino lamb.
In our study, axonal spheroids were consistently more strongly immunopositive to dynein than kinesin; differential expression of these motor proteins in spheroids has also been reported in some human neurodegenerative diseases. 6 In human motor neuron disease, for example, spheroids show strong kinesin immunopositivity, whereas dynein immunoreactivity is scant or absent. In these cases, it was suggested that kinesin might be selectively absorbed to spheroids because their contents had a greater affinity for kinesin than dynein. 14
The exact mechanism of axonal spheroid formation is unresolved, and there is still some doubt as to whether spheroids are the cause, or a consequence, of axonal transport failure.1,4,7 However, it is known that, once axonal spheroids have taken shape, they can further sequester other elements, including kinesin and dynein, 7 which might explain the spheroidal expression of these motor proteins in our NAD cases.
There is increasing evidence implicating disruption of axonal transport in the pathogenesis of a range of human neurodegenerative disorders, including Alzheimer’s disease, frontotemporal dementia, motor neuron disease, amyotrophic lateral sclerosis, and Parkinson and Huntington diseases, although defects probably occur in different components of the transport mechanism, and diverse neuronal populations are injured. Mutations in human genes encoding motor proteins provide more direct evidence that derangements in axonal transport contribute to neurodegeneration, supported by studies in transgenic mouse models.10,12
There are a number of different mechanisms by which axonal transport can be disrupted. Mutations in the motor proteins, kinesin and dynein, may impede axonal anterograde transport and recycling of synaptic metabolites, respectively. Activated kinases may damage kinesin and dynein; mitochondrial injury produced by oxidative stress may lead to energy deprivation; excess intracellular calcium may activate calpain proteolytic degradation of microtubules; and damage to transported materials may inhibit their attachment to motor proteins. Disrupted axonal transport may also cause accumulation of autophagosomes, which move bidirectionally along microtubules, and predispose to neurodegeneration. 11
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
We thank SA Pathology, Adelaide, for funding support.