
Abstract
Staphylococcus pseudintermedius is an opportunistic pathogen of dogs and cats. A high-resolution melting analysis (HRMA) protocol was designed and tested on 42 clinical isolates with known fluoroquinolone (FQ) susceptibility and gyrA codon 84 and grlA codon 80 mutation status. The HRMA approach was able to discriminate between FQ-sensitive and FQ-resistant strains and confirmed previous reports that the main mutation site associated with FQ resistance in S. pseudintermedius is located at position 251 (Ser84Leu) of gyrA. Routine, HRMA-based FQ susceptibility profiles may be a valuable tool to guide therapy. The FQ resistance-predictive power of the assay should be tested in a significantly larger number of isolates.
Staphylococcus pseudintermedius is an opportunistic, zoonotic pathogen of dogs and cats, and fluoroquinolone (FQ)-resistant strains are isolated with increasing frequency from infection sites.1,4,8 FQ resistance in S. pseudintermedius is predominantly mediated by a mutation located at position 251 of the gyrA subunit of DNA gyrase enzyme (Ser84Leu), together with a mutation located at position 80 of the grlA subunit of topoisomerase IV enzyme (Ser80Ile).3,7,10 Importantly, in S. pseudintermedius, drug efflux pumps are not responsible for conferring FQ resistance. 13 The standard antimicrobial disk susceptibility test 2 allows a phenotypic assessment of FQ resistance status. The molecular techniques currently used for the genetic characterization of FQ resistance usually require sequence-specific probes or post-amplification processing such as gel electrophoresis and DNA sequencing. 7 Thus, there is a need for reliable and accurate methods allowing fast identification of gyrA- and grlA-mediated FQ resistance in S. pseudintermedius strains isolated during routine veterinary clinical practice. High-resolution melting analysis (HRMA) offers a low-cost, closed-tube approach to amplicon analysis with the capacity for single-nucleotide discrimination and easy integration with real-time polymerase chain reaction (qPCR).9,13 This technique is increasingly used in microbiology for species identification, detection of resistance-carrying mutations, and genotyping.5,16,18 We screened 42 S. pseudintermedius strains for the presence of gyrA Ser84Leu mutation and grlA Ser80Ile mutation using specific HRMA protocols.
The strains used in our study were obtained as the sole or prevalent bacterial isolate from canine skin, external ear, or conjunctiva samples submitted for microbiologic examination to the Department of Veterinary Sciences and Public Health, University of Milan, from 2009 to 2013. Bacterial isolation was performed through cultivation on tryptone soy agar plates containing 5% sheep blood (Oxoid, Rodano, Milano, Italy) for 24 h at 37°C under aerobic conditions. Isolates were identified as S. pseudintermedius by morphology, Gram stain, catalase activity, growth on mannitol salt agar selective medium, and by the API-Staph identification kit (bioMérieux Italia, Campo a Ripoli, Firenze, Italy). The strains were tested for susceptibility to enrofloxacin and marbofloxacin by the disk diffusion method as follows. A 0.5 McFarland standardized suspension of each bacterial isolate was swabbed over the surface of a Mueller–Hinton agar plate (Oxoid), and paper disks containing either enrofloxacin or marbofloxacin (5 µg per disk) were placed onto the inoculated surface. After overnight incubation at 37°C, the diameters of the zones were measured, and the result was interpreted as susceptible, intermediate, or resistant according to the criteria recommended by the Clinical and Laboratory Standards Institute (CLSI). 2 The quality control (QC) strain used was S. aureus ATCC 25923. Following this testing, 16 S. pseudintermedius isolates were determined to be resistant to enrofloxacin and marbofloxacin; 25 isolates were sensitive to both FQs, and the 1 remaining (isolate 12) showed intermediate susceptibility (Table 1).
Comparison between antimicrobial disk susceptibility testing for enrofloxacin and marbofloxacin with the results obtained by DNA sequencing and high-resolution melting analysis (HRMA) for gyrA and grlA mutation status of Staphylococcus pseudintermedius isolates.*
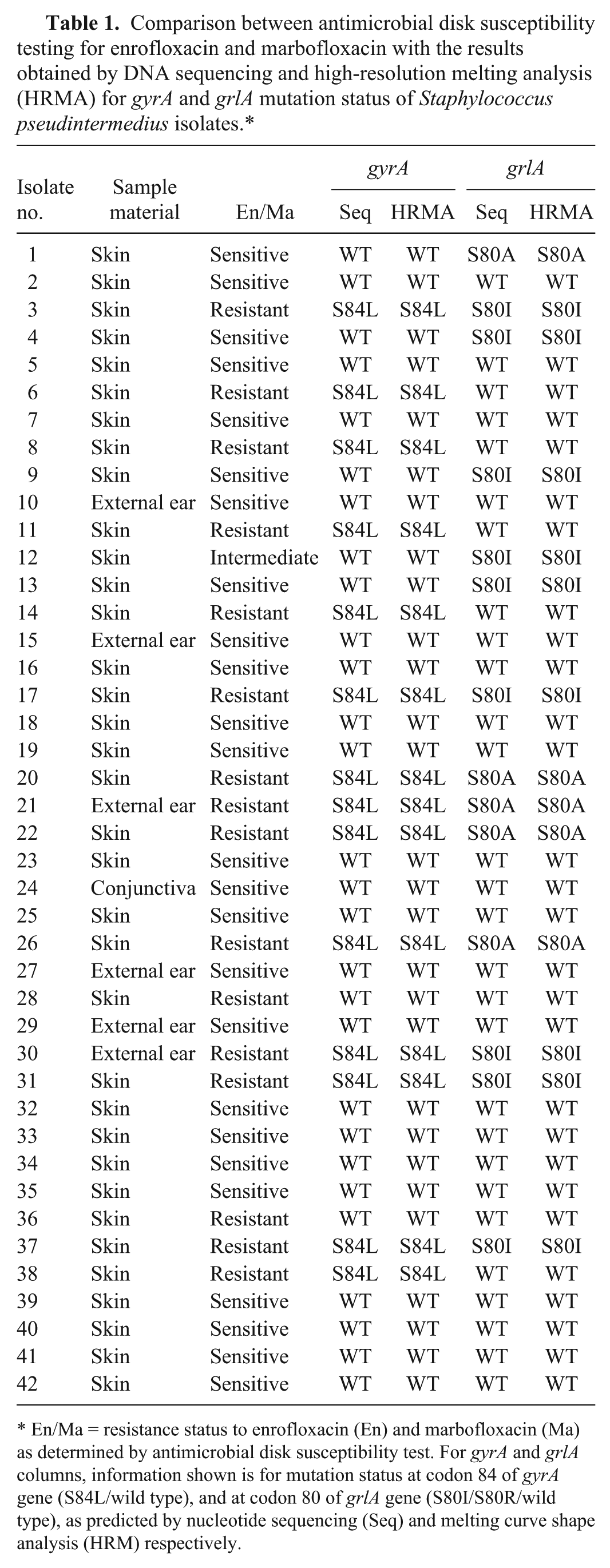
En/Ma = resistance status to enrofloxacin (En) and marbofloxacin (Ma) as determined by antimicrobial disk susceptibility test. For gyrA and grlA columns, information shown is for mutation status at codon 84 of gyrA gene (S84L/wild type), and at codon 80 of grlA gene (S80I/S80R/wild type), as predicted by nucleotide sequencing (Seq) and melting curve shape analysis (HRM) respectively.
The coding sequences of S. pseudintermedius gyrA and grlA genes were obtained from NCBI (http://www.ncbi.nlm.nih.gov; accession NC_014925.1). Degenerate primer sequences available in the literature for gyrA and grlA amplification in staphylococci were adapted to S. pseudintermedius. GyrAsmod forward primer 5’-ATGAGTGTTATCGTATCTCGTGC-3’ and GyrAasmod reverse primer 5’-CCATCGAACCGAAGTTACCTTG-3’ were used for the amplification of a gyrA 262-bp fragment. 7 GrlAsmod forward primer 5’-AATACGTATGATAAACATTTTCG-3’ as reported in the literature 7 and GrlA2 reverse primer 5’-AGCACGTGACGTAATTTCCAGTCTT-3’(designed on NC_014925.1 GenBank nucleotide sequence, gene coordinates 249–274) were used for the amplification of a grlA 137-bp fragment.
For target sequence analysis, each S. pseudintermedius isolate was tip-sampled from a single colony and placed in a 500-µL vial containing 50 µL of water. Samples were then subjected to 3 alternating cycles of 5 s at 95°C and 5 s at −80°C to achieve bacterial cell lysis and DNA release. The bacterial lysates were then subjected to conventional PCR amplification in a gradient thermocycler (Mastercycler, Eppendorf, Hamburg, Germany). The PCR reactions were carried out in duplicate in a total volume of 20 µL containing 1 µL of bacterial lysate, 1× Taq buffer containing 1.5 mm MgCl2 and 200 µm dNTP, 1.25 U of GoTaq DNA polymerase (Promega Italia, Milano, Italy), and 500 nm of GyrAsmod and GyrAasmod primers. The thermal profile for the amplification was 94°C for 90 s; 35 cycles of 94°C for 45 s, 51°C for 30 s, 72°C for 90 s, and a final elongation step at 72°C for 300 s. The same protocol was applied for the identification of grlA but using the GrlAsmod and GrlA2 primer pair. Amplification products were run on 2% agarose gel and gel-purified (QIAquick gel extraction kit, Qiagen, Hilden, Germany). The purified amplicons were then sequenced using standard technology (Prism 310, Applied Biosystems, Foster City, CA). The obtained sequences were aligned using ClustalW software (www.clustal.org/clustal2) to the expected sequence for gyrA and grlA genes of S. pseudintermedius.
For the gyrA gene, 14 FQ-resistant isolates carried the already described C to T transition at position 251 leading to substitution from Ser to Leu at codon 84. The remaining 2 FQ-resistant isolates, all of the 25 FQ-sensitive isolates, and the FQ-intermediate isolate showed no mutation at position 251. None of the isolates carried any other mutations in the amplified target sequence (Table 1). For the grlA gene, 5 FQ-resistant isolates, 3 FQ-sensitive isolates, and the FQ-intermediate isolate carried the already reported G to T transversion at position 239 leading to the Ser80Ile mutation. Four FQ-resistant isolates and one FQ-sensitive isolate carried a different nucleotide transversion from A to C at position 238 leading to a Ser to Arg mutation of the codon 80. The remaining 7 FQ-resistant isolates and 21 FQ-sensitive isolates showed absence of the above mutations (Table 1). Taken together, 9 FQ-resistant isolates carried both the C to T mutation at position 251 of the gyrA gene and either the G to T mutation at position 239 (5 strains) or the A to C mutation at position 238 (4 strains) of the grlA gene.
The same primer pairs used for conventional PCR were then used for qPCR and HRMA (Eco real-time PCR system, Illumina, San Diego, CA). Each amplification reaction contained 1 µL of bacterial lysate, 7.5 µL of reaction mix (SsoFast EvaGreen, Bio-Rad Laboratories), 300 nm of each primer, and H2O to reach a total volume of 15 µL. The thermal protocol for the PCR was 95°C for 600 s, 40 cycles of 95°C for 5 s, and 51°C for 5 s. The melting protocol was 95°C for 15 s, 55°C for 15 s, ramping to 95°C with fluorescence data acquisition at each 0.1°C increment, and a final step at 95°C for 15 s. Sample melt curves were subjected to normalization and temperature shifting. The difference plot for each sample was generated for subsequent grouping analysis. A mixed template of FQ resistant and FQ-sensitive (1:1) strains was used as control to check sequence specificity of the melting profile. When analyzing the melting curve shape obtained through HRMA, the amplicons containing mutations were characterized by a different melting temperature (Tm) with respect to the corresponding wild-type (WT) amplicons. Graphically, the different Tm of a DNA double helix corresponds to a shift of fluorescence, which occurs in a different way between the amplicons based on the nature of the polymorphism. In the present case, the melting curves having different shapes reflected the WT or mutated gyrA/grlA status of the corresponding isolates.
A shift of the temperature towards a lower value was detected in the same 14 isolates carrying the mutation from C (characterized by 3 hydrogen bonds) to T (characterized by 2 hydrogen bonds) at position 251 of the gyrA gene (Fig. 1; Table 1). Regarding grlA, a shift was detected in the 9 isolates carrying the mutation from G to T at position 239 leading to Ser to Ile amino acid change. An alternative curve shape was detectable in 5 isolates reflecting the nucleotide transversion from A to C at position 238 leading to Ser80Arg mutation. The remaining 28 isolates showed the WT shape curve, reflecting absence of mutation in the target codon (Fig. 2; Table 1).
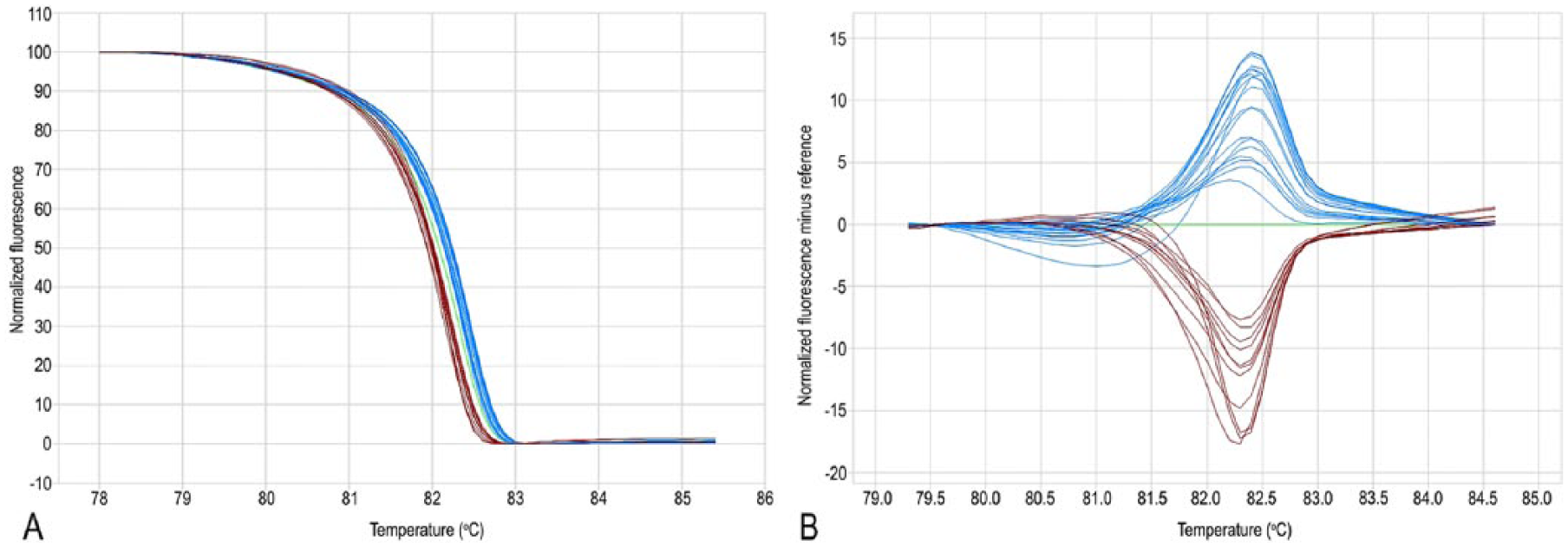
Real-time PCR–high-resolution melting analysis of Staphylococcus pseudintermedius gyrA locus.
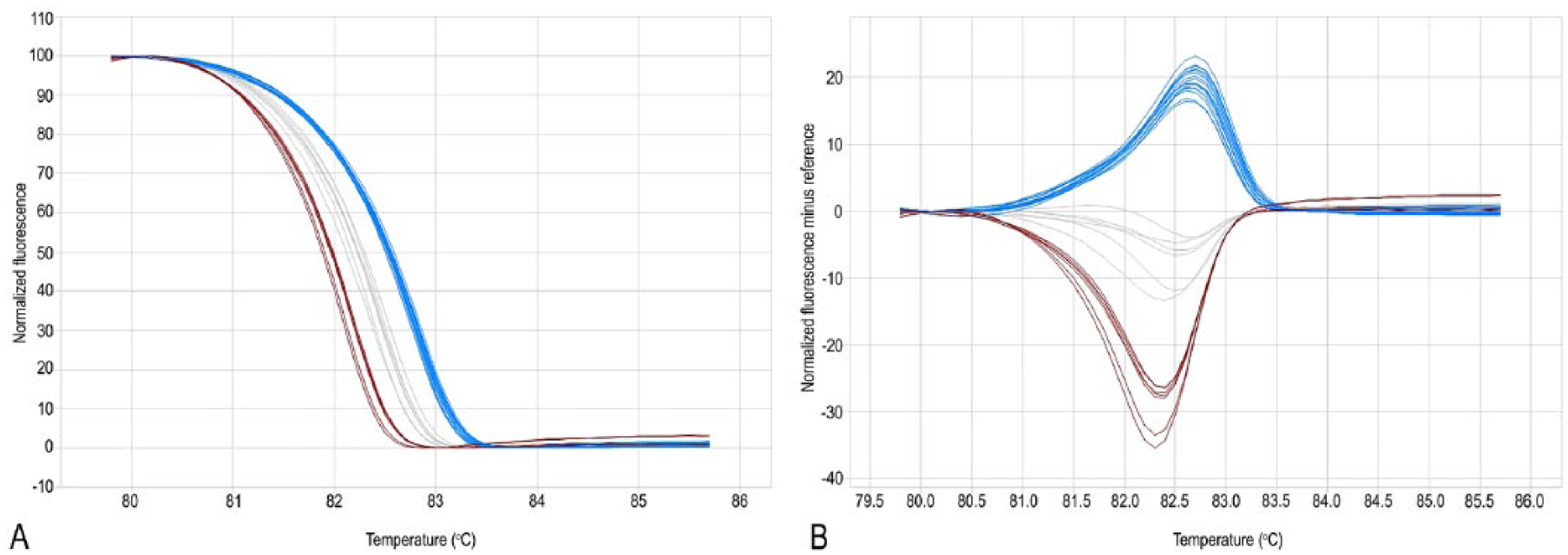
Real-time PCR–high-resolution melting analysis of Staphylococcus pseudintermedius grlA locus.
When comparing the results of the antibiogram with the results of DNA sequencing of both mutation hotspots, a close correlation between sensitivity or resistance to FQs and gyrA mutation was seen: 14 resistant isolates carried the Ser84Leu mutation of gyrA; 25 sensitive isolates carried the WT codon of gyrA; 1 intermediate isolate carried only the Ser80Ile mutation of grlA gene; finally, only 2 resistant isolates carried the WT codon for both gyrA and grlA genes (Table 1). Accordingly, the same 14 isolates assessed as resistant by DNA sequencing were also detected by the HRMA method through discrimination of curve shapes linked to Ser84Leu mutation status of gyrA. Only 2 resistant isolates showed no mutations for gyrA and grlA. In particular, no mutations were found in gyrA amplicons other than C251T in 14 resistant isolates. It is conceivable that different mutations in gyrA or other relevant genes may confer FQ resistance to S. pseudintermedius. On the other hand, our research was aimed only to provide a proof-of-concept regarding the predictive power of gyrA C251T mutation screening for FQ assessment, and performing a comprehensive molecular analysis of resistant isolates was outside the scope of our study.
The mutations found in grlA amplicons seemed to not correlate with FQ resistance, with the exception of the intermediate strain that carried only the grlA mutation, but not the gyrA mutation, which could explain partial resistance. This is in accordance with previous studies reporting that intermediate-resistant S. pseudintermedius have mutations in grlA alone. 11
In our study, the qPCR-HRMA protocol targeting the C251T mutation of the gyrA gene was able to discriminate among FQ-sensitive and FQ-resistant strains on the basis of the corresponding melt curves with complete agreement with data obtained by DNA sequencing of the same gene as a reference technique and by FQ susceptibility testing. Nucleotide polymorphisms not related to resistance status can affect interpretation of HRMA results. In our study, the targets for gyrA and grlA gene amplification were selected on the basis of mutation scanning data available in the literature. 3 In detail, the nucleotide sequences within primer annealing sites involved only mutations leading to amino acid substitutions in FQ-resistant S. pseudintermedius. With one exception noted below, no silent mutations, or mutations causing amino acid substitutions in both FQ-resistant and FQ-susceptible strains, have been described. A silent mutation has been described previously at the nucleotide position 159 of the grlA gene sequence. 3 However, this potential mutation should not interfere with the HRMA because the mutation lies within the annealing sequence of the grlA forward primer, making it irrelevant for the HRMA.
Thus, our work provides a proof-of-concept of a HRMA-based approach for rapid (i.e., within 3 h after isolation of bacterial colonies) identification of gyrA-linked FQ resistance in S. pseudintermedius clinical isolates at the molecular level, without the need to perform DNA sequencing or to use mutation-specific probes. We also confirmed previous reports that, in S. pseudintermedius, the main mutation site associated with FQ resistance is at position 251 (Ser84Leu) of gyrA as detected in 14 of 16 of the tested resistant isolates. The HRMA method of detecting the gyrA mutation hotspot is especially relevant when considering that efflux pumps seem not to be involved in FQ resistance of S. pseudintermedius. 15
This study was performed on a limited number of FQ-resistant S. pseudintermedius isolates (16) that were obtained from canine clinical samples. The number of screened isolates and the rate of FQ resistance across the study period were comparable to susceptibility studies already performed in S. pseudintermedius.10,12 As well, the number of resistant isolates was the same as used in a previous study demonstrating a high correlation between HRMA and a reference assay for the detection of linezolid resistance in Staphylococcus epidermidis. 5 Thus, the methodology described herein has the potential to be used for assessing FQ resistance linked to S. pseudintermedius, although more resistant strains should be included in a further evaluation to determine the predictive value of this assay in the veterinary clinical routine.
Two FQs currently licensed for use in dogs in Italy (i.e., Baytril, Bayer, Milano, Italy; Marbocyl, Vétoquinol Italia, Bertinoro, Italy; https://goo.gl/LxPSkX) were selected in our study to test bacterial resistance. Enrofloxacin is the most commonly used FQ in veterinary medicine and a primary antimicrobial agent to screen FQ resistance in staphylococci.6,10 Marbofloxacin is a third-generation FQ that is also widely used in veterinary clinics and in the laboratory assessment of FQ resistance. 14 The additional use of a less potent quinolone such as nalidixic acid in our study could have given more information on the resistance profile of the strains, in particular with regard to the intermediate isolate. However, nalidixic acid is not widely used in the routine treatment of canine infections.
In clinical practice, treatment of canine infections is often based on the empirical use of antibiotics without previous assessment of antimicrobial susceptibility. Improper use of antibiotics against bacterial pathogens can increase the risk of treatment failure in dogs, and also enhance the possibility of transmission of antibiotic-resistant bacteria to humans that are in contact with the animals.6,7 The development of susceptibility tests suitable for use in routine diagnostic laboratories may help increase the effective use of antibiotics and reduce the spread of FQ resistance. The Ser84Leu change in gyrA has been reported to be the most widespread mutation among S. pseudintermedius exhibiting high FQ resistance, a finding that was confirmed also in our study. 11 The analysis of gyrA mutation status through HRMA can offer some practical advantages when compared to other available molecular techniques, such as DNA sequencing and PCR-based restriction fragment length polymorphism analysis.5,14,17 First, both amplification and analysis are performed in the same tube as a closed system, thus greatly reducing the risk of contamination. Second, HRMA is not particularly laborious and avoids delicate steps (e.g., restriction digestion). Third, the HRMA assay can generate results in a shorter time (2–3 h) compared to the other techniques including the disk diffusion method.
Footnotes
Acknowledgements
We thank Dr. Fabrizio Fabbrini, Clinica Veterinaria Papiniano, Milan, Italy, for providing most of the clinical samples that were the sources of the S. pseudintermedius isolates.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was financially supported by Programma dell’Università per la Ricerca 2008–2009 grants of M Mortarino and PA Martino.