
Abstract
Salmonella is one of the most common causes of foodborne illnesses in humans in the United States, and domestic poultry is considered an important source of this pathogen. Salmonella enterica subsp. enterica serovar Heidelberg is the fourth most commonly reported Salmonella from retail meats and food animals in the United States. We assessed the genotypes and antimicrobial resistance phenotypes of Salmonella Heidelberg isolated from various chicken and turkey hatcheries and breeder farms in the Midwest. The genotypes of 33 S. Heidelberg isolates from chickens (n = 19) and turkeys (n = 14) were compared using pulsed-field gel electrophoresis analysis. Cluster analysis of the fingerprints showed that the majority of the chicken isolates grouped together with 87% similarity; those from turkeys clustered with 88% similarity. Similarity between chicken and turkey isolates was also high (86%). Isolates from turkeys were generally more genetically diverse than those from chickens. Antimicrobial susceptibility analysis detected resistance to sulfisoxazole (36% of the isolates), streptomycin (33%), gentamicin (27%), tetracycline (24%), ampicillin and amoxicillin–clavulanic acid (15%), cefoxitin (12%), ceftriaxone and ceftiofur (12%), and chloramphenicol (9%). None of the isolates was resistant to azithromycin, ciprofloxacin, or nalidixic acid. Although the number of the isolates was limited in our study, we conclude that S. Heidelberg isolates from the same host generally clustered together and that a considerable number of the isolates were resistant to a number of antimicrobial agents.
Salmonella is a leading cause of foodborne illness in humans worldwide. 20 In the United States alone, more than one million foodborne illnesses are associated with nontyphoidal Salmonella spp. each year, 22 causing significant losses in medical care costs and decreased productivity. Salmonella enterica subsp. enterica serovar Heidelberg is one of the more important serovars that cause infections in humans, resulting in mild diarrhea to severe systemic illness.5,24,25 S. Heidelberg was the seventh most common serovar isolated from humans in 2011 and is believed to cause ~84,000 cases of salmonellosis in the United States annually. 16
Domestic poultry are known carriers of S. Heidelberg, a serovar that can cause clinical disease in young birds, although mature poultry often exhibit no clinical signs of infection.5,6 Poultry products can be contaminated with S. Heidelberg via 2 major routes: 1) meat products can become contaminated during processing 24 and 2) as a consequence of systemic infection, S. Heidelberg can invade the muscle of meat birds or infect eggs through the reproductive tissue of laying hens.12,13 Therefore, chicken breasts, ground turkey meat, and eggs are considered major sources of S. Heidelberg infections (CDC, 2013, National enteric disease surveillance: Salmonella annual report. Available at: https://goo.gl/tqA7WC). As well, case studies suggest that table eggs and chicken meat are the main sources of S. Heidelberg in humans in Canada and the United States.5,8 Notably, S. Heidelberg has a high potential of causing severe systemic disease in humans, unlike most other serovars associated with poultry. 8
In pregnant women and children younger than 5 y, ceftriaxone is used for the treatment of Salmonella infections.9,11 However, antimicrobial resistance has been increasing in S. Heidelberg, and this appears to be contributing to reduced effectiveness of therapeutic antimicrobials used in human health.7,10,11 Hence, it is essential to monitor and characterize antimicrobial resistance in S. Heidelberg from poultry to evaluate the potential impact of these strains on public and consumer health.
There is also a need to determine the genotypic relationship among S. Heidelberg strains isolated from poultry to devise adequate intervention strategies based on the prevalence in various poultry-associated sources of infection. One of the techniques for genotyping is pulsed-field gel electrophoresis (PFGE), a DNA-based typing method that has a high power of discrimination and reproducibility. 14 PFGE is currently considered the gold standard method for determining relatedness between foodborne bacterial pathogens and is used by PulseNet, the national molecular subtyping network for foodborne disease surveillance in the United States (https://goo.gl/fM3b08). Based on the aforementioned observations, our objective in this study was to determine the genotypic relatedness and antimicrobial resistance of S. Heidelberg isolated from various chicken and turkey farms located in the Midwestern United States.
To achieve our objective, 33 putative S. Heidelberg isolates were obtained from poultry laboratories located in the Midwest. The isolates were submitted between April and July of 2015 from chicken (n = 19) and turkey (n = 14) farms. The isolates were recovered from different sample types, including environmental booties (29 of 33), hatchery debris (2 of 33), and cloacal swabs (2 of 33). The isolates were subcultured on trypticase soy agar (TSA) plates and incubated at 37°C for 24 h. Pure colonies were selected and tested by polymerase chain reaction (PCR) to confirm their identity as S. Heidelberg. For this purpose, DNA was obtained using a QIAamp DNA mini kit (Qiagen, Valencia, CA) and then quantified using a NanoDrop ND-2000 spectrophotometer (Thermo Scientific, Waltham, MA). Primers specific to the Salmonella genus (F: ATCGCTGACTTATGCAATCG, R: CGGGTTGCGTTATAGGTCTG) and to serovar Heidelberg (F: TGTTTGGAGCATCATCAGAA, R: GCTCAACATAAGGGAAGCAA) were used in the PCR analysis.2,21 The PCR products were analyzed using gel electrophoresis, and the size of the amplicons was determined by comparison with a DNA marker (All purpose Hi-Lo DNA marker, Bionexus, Oakland, CA).
To determine the genetic relatedness of the isolates, PFGE analysis was performed according to the CDC protocols with slight modification (https://goo.gl/fM3b08). Briefly, isolates (n = 33) were grown at 37°C overnight on TSA plates and transferred to a cell suspension medium (100 mM Tris–100 mM ethylenediamine tetra-acetic acid [EDTA], pH 8.0) to achieve an optical density (OD610) of 1.3–1.4. Plugs were prepared by adding and gently mixing 400 µL of 1% agarose gel (SeaKem LE agarose, Lonza, Rockland, ME) to 400 µL of each cell suspension. The mixtures were dispensed to plug molds. (Bio-Rad, Hercules, CA). Lysis of the cells was performed by incubating the plugs in 5 mL of cell lysis buffer (50 mM Tris:50 mM EDTA [pH 8.0]:1% sarcosyl) and proteinase K (20 mg/mL) at 55°C for 2 h. The plugs were then washed twice with preheated Tris–EDTA buffer (55°C) and 3–4 times with preheated clinical laboratory reagent water (CLRW). For restriction digestion, the plugs were incubated with XbaI (10 U/mL) for 1.5–2 h at 37°C. The plugs were then loaded into the wells of the gel, and electrophoresis was conducted using a PFGE unit (CHEF-DR variable angle system, Bio-Rad) at 6 V for 18–19 h. S. Braenderup H9812 was used as a control strain and as an additional marker to facilitate comparison between gels. The gels were then stained for 30 min in ethidium bromide (10 mg/mL), destained in CLRW for 20 min (3–4 times), and photographed under ultraviolet light.
The PFGE fingerprint images were analyzed using Bio-Numerics software (Applied Maths, Austin, TX) to generate dendrograms that highlighted similarities among the fingerprints. Similarity and cluster analysis were performed using the Pearson correlation coefficient with 2% optimization and 1% position tolerance and the unweighted pair group method with arithmetic means algorithm. Furthermore, jackknife analysis was used to determine if the fingerprints could be assigned accurately to each host group (chicken or turkey).
The resistance of the isolates to antimicrobials was determined using Sensititre NARMS gram-negative plates (Thermo Fisher Scientific) that contained 14 different antimicrobials that target gram-negative bacteria. 1 The antimicrobials were: cefoxitin (0.5–32 mg/mL), azithromycin (0.12–16 mg/mL), chloramphenicol (2–32 mg/mL), tetracycline (4–32 mg/mL), ceftriaxone (0.25–64 mg/mL), amoxicillin–clavulanic acid 2:1 (1/0.5–16/8 mg/mL), ciprofloxacin (0.015–4 mg/mL), gentamicin (0.25–16 mg/mL), nalidixic acid (0.5–32 mg/mL), ceftiofur (0.12–8 mg/mL), sulfisoxazole (16–256 mg/mL), trimethoprim–sulfamethoxazole (0.12/2.38–4/76 mg/mL), ampicillin (1–32 mg/mL), and streptomycin (2–64 mg/mL). Escherichia coli ATCC25922 was used as a control strain. The minimum inhibitory concentration, defined as the lowest concentration of an antimicrobial that produced no visible growth, was recorded for each antimicrobial. For interpreting resistance, the breakpoints adopted by the National Antimicrobial Resistance Monitoring System (NARMS) guidelines, were used (CDC, 2010, National Antimicrobial Resistance Monitoring System: enteric bacteria. Human isolates final report, 2010. Available at: https://goo.gl/qd3FvG). Isolates that exhibited resistance to more than 2 classes of antimicrobials were considered multidrug resistant (MDR). 19
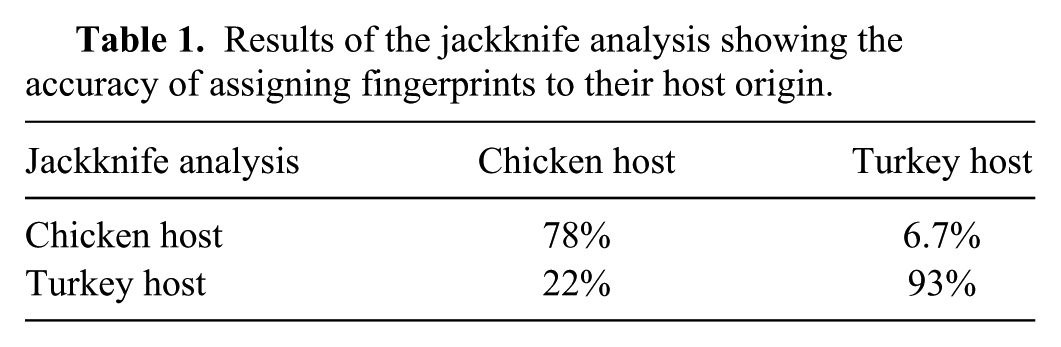
Our results showed that 10 of the 19 PFGE fingerprints of Salmonella isolated from chickens grouped together with 87% similarity (cluster A); 6 of 14 turkey strains clustered together with 88% similarity (cluster B; Fig. 1). Clusters A and B shared a relatively high similarity (86%). However, isolates from turkeys were generally more genetically diverse than those from chickens. For example, 2 clusters, composed of 2 turkey isolates each (NS014 and NS015; NS021 and NS022), were 74% and 63% similar to the clusters A and B, respectively (Fig. 1). The highest similarity (99%) between chicken and turkey isolates was observed between the fingerprint of 1 chicken isolate (NS031) and 2 turkey isolates (NS033; NS034; Fig. 1). In contrast, the fingerprint of another chicken isolate (NS011) shared a comparatively low similarity (50% and 12%, respectively) to all other fingerprints (Fig. 1). Furthermore, the dendrogram displayed 3 main clusters that shared a relatively low similarity (12%; Fig. 1). Although the reasons that influenced this similarity are not clear currently, it was notable that each of the clusters was composed of fingerprints from both turkey and chicken isolates. Regardless, the general grouping of the isolates according to the host of origin was further confirmed using the jackknife analysis that showed that chicken and turkey isolates could be assigned to chicken and turkey groups with 78% and 93% accuracy, respectively (Table 1).

A dendrogram showing the similarity between the pulsed-field gel electrophoresis patterns of the Salmonella isolates. Similarity and cluster analysis were performed using the Pearson correlation coefficient with 2% optimization and 1% position tolerance and the unweighted pair group method with arithmetic means algorithm.
Results of the jackknife analysis showing the accuracy of assigning fingerprints to their host origin.
The high similarity of the strains within and between hosts was not surprising because it has been previously shown that 107 clinical and agri-food isolates of S. Heidelberg from Canada displayed limited genetic diversity, 3 corroborating our PFGE analysis. In another study, 6 the majority of 49 S. Heidelberg isolates from different animal sources clustered into 3 groups that were potentially related with a difference of only 2–3 bands among genotypes of these strains. However, 7 isolates exhibited unique patterns with >3 band difference among them. 6 The isolates in this previous study 6 were randomly selected from a historical collection (1991–2013) and were retrieved from a variety of hosts, including cattle, chickens, horses, and turkeys. In comparison, our isolates were collected over a significantly shorter period of time, solely from turkeys and chickens, and likely reflect a different geographical area (Midwest). Regardless, it should be noted that high similarity between the PFGE fingerprints in our study does not necessarily reflect similarity in important phenotypic traits, such as antimicrobial resistance (see below), that might be associated primarily with extrachromosomal genetic material. 3
Collectively, 14 of 33 isolates tested (42%) were classified as MDR (Table 2). Notably, NS011 and NS029 were resistant to 6 antimicrobials each; NS011 displayed resistance to 3 additional antimicrobials. Furthermore, NS012, NS015, and NS025 were each resistant to 5 antimicrobials (Table 2). One (3%) isolate (NS001) was resistant to 1 antimicrobial; 16 (48%) isolates were not resistant to any of the antimicrobials tested. The highest resistance (36%) was observed against sulfisoxazole (12 of 33) followed by streptomycin (33%; 11 of 33), gentamicin (27%; 9 of 33), tetracycline (24%; 8 of 33), ampicillin and amoxicillin–clavulanic acid (15%; 5 of 33), cefoxitin, ceftriaxone, and ceftiofur (12%; 4 of 33), and chloramphenicol (9%; 3 of 33). There was no resistance against azithromycin, ciprofloxacin, and nalidixic acid. Eight (24%), 5 (15%), and 7 (21%) isolates were resistant to tetracycline, ampicillin, and gentamicin, respectively. A relatively large number of isolates was resistant to sulfisoxazole (36%) and streptomycin (33%), whereas none of the isolates displayed resistance to nalidixic acid (Table 2).
Antimicrobial susceptibility of Salmonella isolates.*
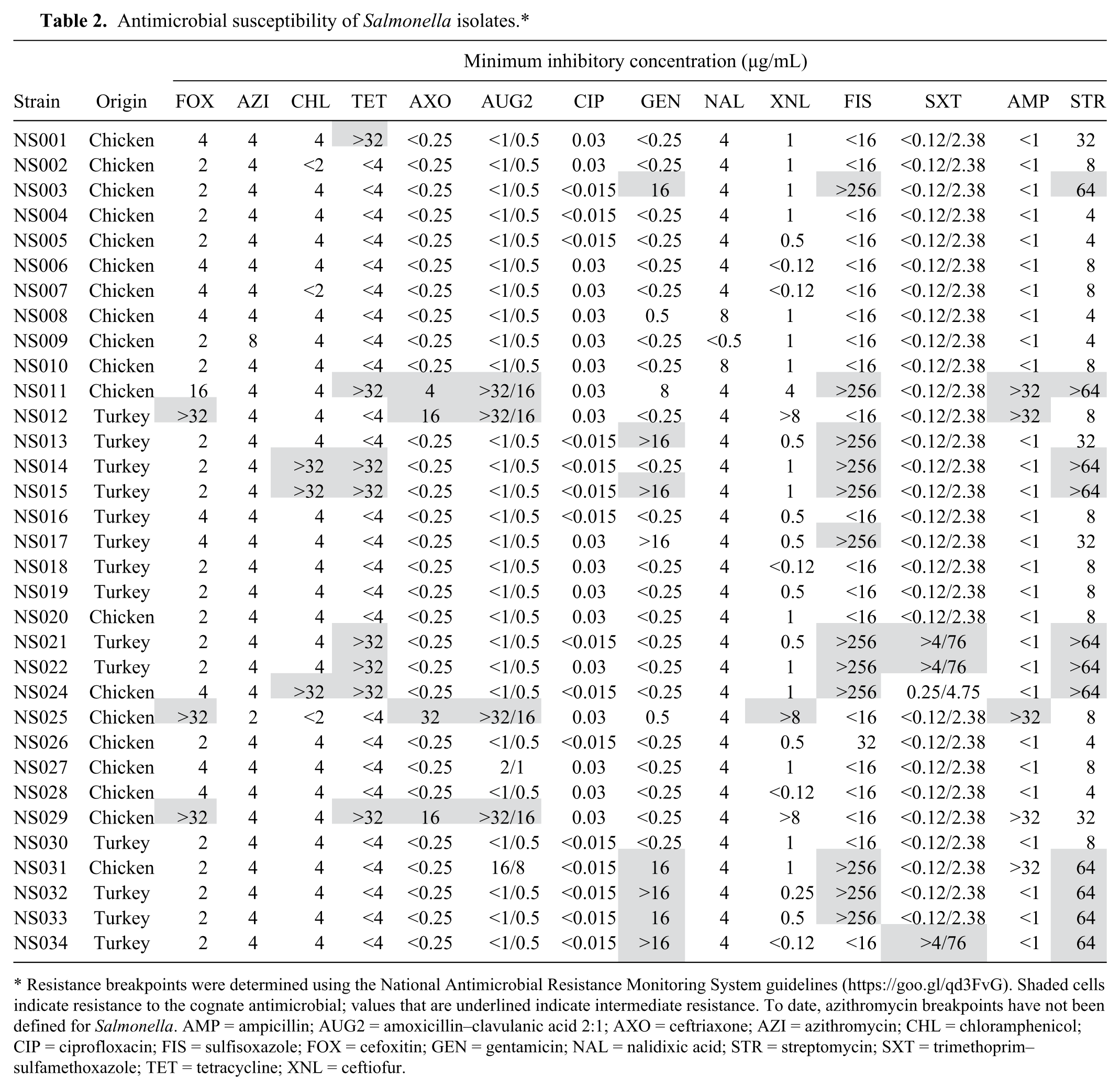
Resistance breakpoints were determined using the National Antimicrobial Resistance Monitoring System guidelines (https://goo.gl/qd3FvG). Shaded cells indicate resistance to the cognate antimicrobial; values that are underlined indicate intermediate resistance. To date, azithromycin breakpoints have not been defined for Salmonella. AMP = ampicillin; AUG2 = amoxicillin–clavulanic acid 2:1; AXO = ceftriaxone; AZI = azithromycin; CHL = chloramphenicol; CIP = ciprofloxacin; FIS = sulfisoxazole; FOX = cefoxitin; GEN = gentamicin; NAL = nalidixic acid; STR = streptomycin; SXT = trimethoprim–sulfamethoxazole; TET = tetracycline; XNL = ceftiofur.
The highly resistant NS011, NS029, and NS025 isolates were from chickens, whereas only isolates NS012 and NS015 were from turkeys. One chicken isolate was resistant to gentamicin, whereas 7 turkey isolates were resistant to this antimicrobial. However, 12 chicken isolates and 4 turkey isolates were pan-susceptible to all antimicrobials.
Our results are comparable to those reported earlier in which 28% of S. Heidelberg isolates from animal and food sources were found resistant to antimicrobials. 15 Furthermore, in 2007, up to 38% of S. Heidelberg isolates from retail ground meat were resistant to antimicrobials. 17 Additionally, in our study, all of the chicken isolates were susceptible to azithromycin, ciprofloxacin, nalidixic acid, and trimethoprim–sulfamethoxazole. Similarly, other studies have reported no resistance to nalidixic acid.17,18,25
Approximately 86% of Salmonella species isolated from chickens and quails in Italy were resistant to tetracycline. 4 Contrary to our observations, tetracycline resistance was higher in turkey-, than in chicken-associated isolates in Canada. 23 In our study, 42% (14 of 33) of the isolates were classified as MDR. Although more MDR isolates were obtained from turkey samples in comparison to those from chickens (57% and 32%, respectively), this difference was not statistically significant (p < 0.05). In comparison, in Italy, 4 9% of the overall poultry isolates were MDR, and 11% of the isolates from chicken and quails were MDR The differences in antimicrobial resistance phenotypes between the aforementioned studies can be attributed to: 1) potential differences in farming practices (e.g., conventional, free-range, organic farming), which have been changing over a period of time, 2) variations in the methods adopted to test antimicrobial resistance, and 3) regulations that govern the use of antimicrobials in animals in different countries. The role of these factors in the development and spread of resistance to antimicrobials requires further analysis.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by University of Minnesota Foundation internal funding.