
Abstract
Histomonas meleagridis is a flagellate protozoan organism that can cause severe necrotizing typhlitis and hepatitis in gallinaceous birds. Peafowl (Pavo spp.) have been shown to be susceptible to histomoniasis in experimental settings, but there are few reports of natural histomoniasis in this species. A retrospective study of the archived cases at 2 veterinary diagnostic laboratories in the United States yielded 5 cases of peafowl with gross and histologic findings characteristic of histomoniasis. Lesions included bilateral, transmural fibrinonecrotic typhlitis and multifocal necrotizing hepatitis with associated trophozoites morphologically consistent with H. meleagridis. There was no evidence of Heterakis gallinarum infestation in the studied cases. DNA was extracted from formalin-fixed, paraffin-embedded liver and ceca from all 5 cases and was analyzed using multiple sets of primers with subsequent sequencing and genotyping. Four samples were positive for H. meleagridis, and 1 sample was positive for both H. meleagridis and Tetratrichomonas gallinarum. These results confirm that peafowl develop clinical disease similar to that described previously in other gallinaceous birds infected by H. meleagridis. The role of T. gallinarum remains unknown and further research is necessary to elucidate its role, if any, in the pathogenesis of the observed lesions.
Keywords
Introduction
Histomonas meleagridis (phylum Parabasalia, class Tritrichomonadea, order Tritrichomonadida, family Dientamoebidae/Protrichomonadinae) is an anaerobic protozoan parasite existing in either flagellated or amoeboid forms.4,11 It has a worldwide distribution and transmission occurs primarily via ingestion of embryonated eggs of the cecal nematode Heterakis gallinarum containing H. meleagridis trophozoites or by ingestion of earthworms that have ingested nematode eggs.4,9 –11 The life cycle of the parasite H. meleagridis begins in the ceca of host birds infected with adult H. gallinarum worms. H. meleagridis infects the H. gallinarum ovary by an unknown mechanism so that trophozoite-laden H. gallinarum eggs are shed into the environment with the feces. H. gallinarum then develop into larvae containing H. meleagridis in the soil. When susceptible birds ingest H. gallinarum larvae from the environment, the H. meleagridis passengers are ingested as well. H. gallinarum will then travel through the gastrointestinal tract to the ceca, where the larvae will molt and release the trophozoites into the cecal lumen. 11 Free trophozoites can survive in the external environment for a few hours, but remain viable for up to 2 y within the nematode eggs. 9
Histomoniasis (blackhead disease, typhlohepatitis, infectious enterohepatitis, histomonosis) is characterized by cecal mucosal-to-transmural necrosis with formation of luminal cores. Hepatic lesions are variably present and consist of portal or multifocal-to-coalescing heterophilic granulomas and/or necrosis. 2 Necrotic lesions can also be present in the kidneys, bursa of Fabricius, spleen, air sacs, lungs, pancreas, and proventriculus. 14 Protozoal organisms presumably reach other organs through the portal system after cecal and hepatic infection has been established. 11 H. meleagridis virulence appears to be determined by the protozoal genetics, infectious dose, and influence of cohabiting cecal bacteria (Escherichia coli, Clostridium perfringens; Bacillus subtilis in turkeys) and protozoa (Eimeria tenella in chickens).1,5,11
H. meleagridis causes disease in most gallinaceous birds, including pheasants, chickens, turkeys, partridges, and quails. Turkeys are notoriously susceptible to infection, with outbreak mortality rates approaching 100%.4,11 Ring-necked pheasants are relatively resistant to disease and are considered a reservoir host for the pathogen. 10 Experimental infection of peafowl (Pavo cristatus) and other gallinaceous birds (chukar, guinea fowl, bobwhite quail, pheasants, and others) have been attempted and indicate variable susceptibility among species. 11 These experimental infections suggest that peafowl may have increased susceptibility to H. meleagridis compared with other gallinaceous birds. 9 There are few reports describing naturally occurring disease in peafowl, but commercially raised birds suffered outbreaks similar to those described in chickens and turkeys. 5 Our study describes the pathologic and molecular features of 5 cases of histomoniasis in peafowl.
We performed a retrospective search for cases of histomoniasis in peafowl that were submitted for autopsy at the University of Georgia Athens Veterinary Diagnostic Laboratory (Athens, Georgia) and the Ohio Department of Agriculture Animal Disease Diagnostic Laboratory (Reynoldsburg, Ohio) between 2000 and 2015. All relevant information (clinical history, pathology report, and ancillary testing) was reviewed. Hematoxylin and eosin–stained slides from all cases were also reviewed. Tissues available varied among cases, but ceca and liver were examined in all cases.
DNA was extracted from formalin-fixed, paraffin-embedded liver and ceca using DNA extraction kits a per the manufacturer’s instructions. Extracted DNA from each tested case was analyzed by polymerase chain reaction (PCR) using 3 different sets of primers to amplify the internal transcribed spacer (ITS) region (PCR-1), the 18S rRNA gene (PCR-2), and a conserved region upstream of the β-tubulin gene (PCR-3; Table 1). PCR components included 1–2 μL of extracted DNA in a 25-μL reaction containing PCR beads b and 20 pM each of forward and reverse primers. Nuclease-free water was used as a negative control to detect contamination, and DNA isolated from a laboratory-propagated strain of H. meleagridis was included as a positive control. In PCR-1, the ITS region was amplified using Trichomonadidae-family wide ITSF and ITSR primers described in previous studies (Table 1). 8 Cycling parameters for the amplification were 1 cycle of 95°C for 1 min followed by 35 cycles of 94°C for 15 s, 48°C for 15 s, 72°C for 1 min, and a final extension at 72°C for 15 min. In PCR-2, the 18S rRNA gene of H. meleagridis was amplified using primers described previously (Table 1). 1 Cycling parameters for the amplification were as follows: 1 cycle of 95°C for 1 min; 35 cycles of 95°C for 15 s, 53°C for 15 s, and 72°C for 1 min; followed by final elongation step at 72°C for 10 min. In PCR-3, novel detection primers to amplify a region upstream of the β-tubulin gene identified by genetic sequencing of H. meleagridis are described (Table 1). Cycling parameters for the amplification were 1 cycle of 95°C for 1 min; 35 cycles of 95°C for 15 s, 45°C for 15 s, and 72°C for 1 min; followed by a final elongation step at 72°C for 10 min. Following amplification, all PCR amplicons from all 3 reactions were separated by gel electrophoresis using 1% agarose gel, stained, c and visualized with blue light. DNA was purified using extraction kits d and ligated into a vector c per the manufacturer’s instructions. The DNA transformation procedure was performed using competent cells e and 2 mL of ligation-reaction per the manufacturer’s instructions. Competent cells containing the vector with PCR product insert were detected by plating 50 μL of the transformation mixture on lysogeny broth (LB) agar plates supplemented with 100 mg/mL of carbenicillin. Two to 3 colonies, if available, were isolated and propagated in LB supplemented with 100 mg/mL of carbenicillin. Plasmid DNA was isolated using a kit f per the manufacturer’s instructions. Sequencing of the inserts was performed using pJET 1.2 forward and reverse sequencing primers at the Integrated Biotechnology Laboratories (University of Georgia, Athens, Georgia). Sequences obtained from our study and other related sequences were aligned using a multisequence alignment program. 15
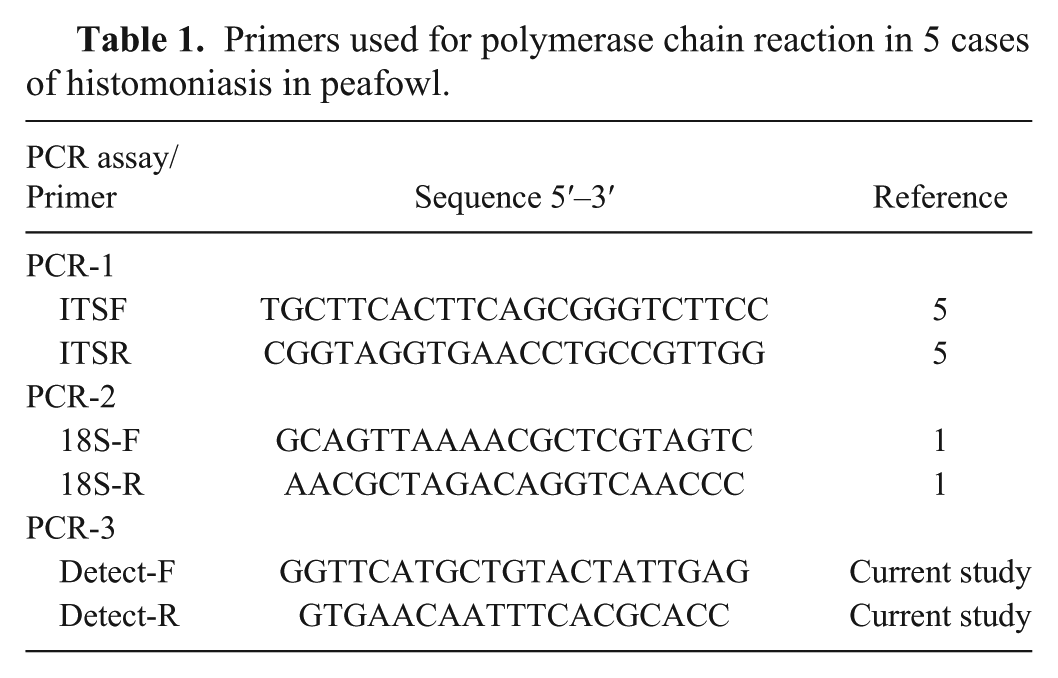
Primers used for polymerase chain reaction in 5 cases of histomoniasis in peafowl.
Reported ages of the retrieved cases ranged from 3 wk to 16 mo (Table 2). Gross findings were fairly consistent among the cases and consisted of bilateral necrotic cecal cores of white-to-tan, fibrinous-to-caseous material (4 of 5), marked cecal distention (4 of 5), with occasional cecal wall thickening and transmural necrosis (1 of 5; Fig. 1). Cases 3–5 had hepatomegaly and multifocal hepatic necrosis, as evidenced by multiple white-to-gray, 1–2 mm diameter, subcapsular foci that were distributed throughout the parenchyma. These foci occasionally had a central area of depression with a surrounding ring of hemorrhage. Histologically, all cases had fibrinonecrotic typhlitis and necrotizing hepatitis. The cecal mucosa was effaced by necrosis and hemorrhage with loss of stromal structures. The lamina propria was expanded by heterophils and macrophages, with fewer lymphocytes and plasma cells that also extended to the submucosa, muscle layers, and serosa (Fig. 2). Within these lesions were multiple, extracellular and intrahistiocytic, round, 10–20 µm diameter, periodic acid–Schiff (PAS) reaction positive protozoal trophozoites with central dark nuclei (Fig. 2 inset). Trophozoites were also present within intraluminal debris and infiltrating into less affected adjacent lamina propria and submucosa. Expanding the liver were multiple, extensive areas of necrosis characterized by hepatocytes with shrunken, hypereosinophilic cytoplasm and pyknotic nuclei that were surrounded by layers of epithelioid macrophages and heterophils with fewer lymphocytes and plasma cells and intralesional protozoa. Overall, there were fewer protozoal organisms in the liver than in the ceca. Protozoal organisms were also less frequently associated with overt necrosis. Two juvenile birds (cases 1, 2) lacked gross hepatic lesions and had less severe histologic changes in the liver compared to the older birds (Figs. 3, 4). Hepatic lesions in these 2 juvenile birds were primarily portal. Less frequent changes included necrotizing airsacculitis with intralesional protozoa (2 of 5), areas of pulmonary necrosis surrounded by epithelioid macrophages and multinucleate giant cells (necrogranulomas) with intralesional fungal hyphae (1 of 5), and ingluvial candidiasis (1 of 5).
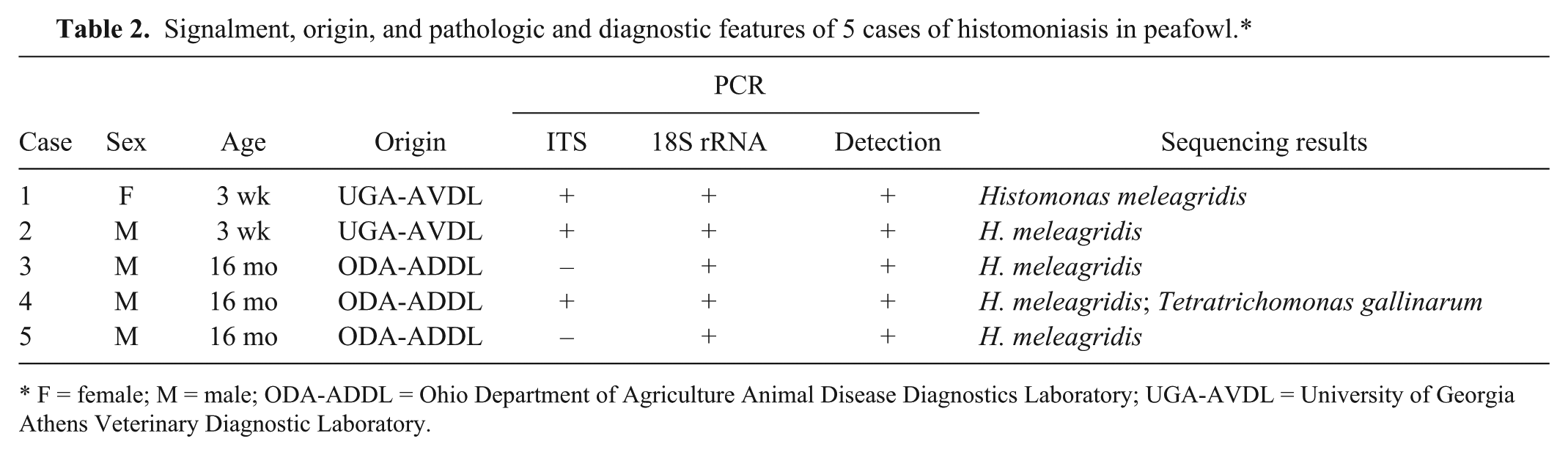
Signalment, origin, and pathologic and diagnostic features of 5 cases of histomoniasis in peafowl.*
F = female; M = male; ODA-ADDL = Ohio Department of Agriculture Animal Disease Diagnostics Laboratory; UGA-AVDL = University of Georgia Athens Veterinary Diagnostic Laboratory.
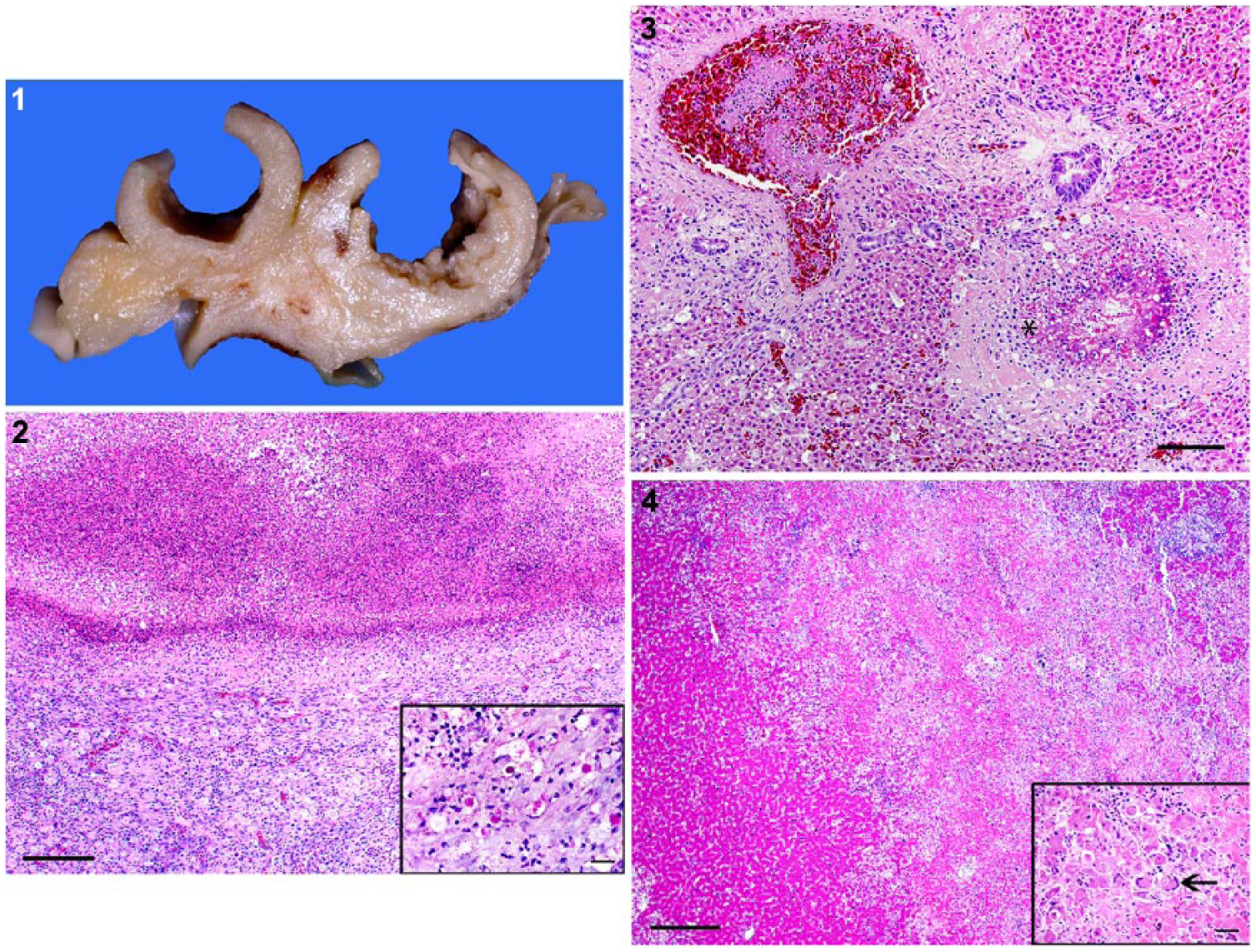
Histomoniasis in peafowl (Pavo cristatus).
Reported ancillary tests included fecal flotation and fecal smears, as well as aerobic, anaerobic, and Salmonella spp. bacterial culture. No birds were positive on fecal smear for coccidia or other protozoa. E. coli and C. perfringens were isolated from both the intestine and liver in case 4. No birds were positive for Salmonella spp. Only case 3 was reported to have received treatment prior to death, including metronidazole, fenbendazole, and enrofloxacin.
All tested birds were positive on at least 2 of the 3 PCR reactions (Table 2). The ITS region did not amplify in cases 3 and 5, whereas the 18S rRNA region and novel detection primers described to be specific for H. meleagridis did amplify appropriate sequences in all 5 cases. The remaining 3 cases were positive using all 3 primer sets. Case 4 was positive by sequencing for both H. meleagridis and T. gallinarum.
There is some evidence that young turkeys and chickens are thought to be more susceptible to infection with H. meleagridis, although mature turkey flocks can also experience severe disease. 9 A similar age-related trend is suggested by our data set, wherein the age range of affected peafowl varied from juvenile to young adult birds. However, given the small number of birds in our study, it is difficult to ascertain the extent of this correlation. The gross and histologic changes in the current cases were consistent among the samples and were compatible with previous descriptions of histomoniasis in other gallinaceous birds.4,9,11 Concurrent infections with other pathogens, similar to our observations, have also been described previously and may be related to concurrent immunosuppression in affected birds. 11 Hepatic lesions in our study were not as severe as those seen in turkeys, which was also noted in previous experimental histomoniasis in peafowl. 9 These lesions tend to be portal at early stages of infection, but become more extensive as the disease progresses. 4 Although no information on clinical course was available from our cases, the fact that the hepatic lesions in the 2 juvenile birds (cases 1, 2) were less severe and primarily portal when compared to the older birds (cases 3–5) may imply that the juvenile birds may have died at an earlier stage of infection. There are reports of H. meleagridis genotype 2 causing more severe cecal changes without concurrent hepatic lesions, implying that affected tissues may depend directly or indirectly on genetically modified parasitic virulence factors. 3
There was no evidence of H. gallinarum infestation in any of the examined cases in our study, which is also consistent with previous experimental infections in peafowl. 9 To our knowledge, H. gallinarum infection without concurrent H. meleagridis infection has not been attempted or described in peafowl and, therefore, it is unknown how susceptible peafowl are to H. gallinarum infection. Species differences in susceptibility have been described in other gallinaceous birds, which could explain our findings. 16 Alternatively, severe pathologic changes in the ceca caused by H. meleagridis infection may provide an inhospitable environment for H. gallinarum.9,16 In such cases, where H. gallinarum is unable to complete its life cycle, the magnitude of clinical disease in susceptible birds caused by H. meleagridis should depend on the number of infected eggs present in the soil or shed by carrier species (chickens and pheasants) in contact with peafowl. Other forms of transmission, such as cloacal drinking and reverse peristalsis, ingestion of insects, and through human handlers have been documented in poultry, 11 but it is unknown if any of these alternative routes are relevant to the disease spread in our cases.
Many of the phylogenetic studies examining H. meleagridis isolates and other parabasalid protozoa have focused on the internal transcribed spacer 1–5.8S rRNA–internal transcribed spacer 2 (ITS1-5.8S-ITS2) regions of the ribosomal gene, which are noncoding sequences with less conservation pressure, leading to high genetic variability.1,5,8,17 Multiple sequence variants, however, can exist within a single isolate, necessitating the need for C-profiling or sequencing to accurately characterize H. meleagridis strains. 8 Subtyping into 2 phylogenetically distinct genotypes has been done using sequences at the 18S rRNA and rpb1 loci, but these analyses still require deep sequencing. 1 The novel detection primers described in our study were designed to obtain a uniquely sized amplicon of H. meleagridis that would allow diagnosticians to distinguish H. meleagridis isolates from other parabasalid organisms without relying on sequencing. In the small sample size in this study, PCR results correlated with sequencing results. Sequencing was still necessary to distinguish the single case of dual infection of H. meleagridis and T. gallinarum (case 4). It is difficult to interpret the presence of T. gallinarum genetic sequences in this case. Further research would be necessary to determine a possible role of T. gallinarum in the development of the observed lesions in these cases or a potential H. meleagridis/T. gallinarum coinfection. In an early pilot study, our novel primers were able to detect T. gallinarum infection in a subset of peafowl cases originally diagnosed as histomoniasis (data not shown). Although the ability of T. gallinarum to cause disease in the lower intestinal tract of birds is debatable given inconsistent experimental results, 12 reports of naturally occurring T. gallinarum infection in gallinaceous and anseriform birds have been described.5,6,13 Our results, however, may imply that coinfections with H. meleagridis and T. gallinarum or with T. gallinarum alone may be more common than currently expected.
Differentiating H. meleagridis from other parabasalid organisms or even from other kingdoms of pathogenic agents may not be precise or accurate using only routine histologic examination. It has long been considered difficult to distinguish H. meleagridis on light microscopy from fungal organisms, necessitating the use of special stains such as PAS or Grocott methenamine silver. 11 It may be also difficult to distinguish H. meleagridis and T. gallinarum on cecal smears, which was one of the standard means of detection of Histomonas for many decades. 5 For these reasons, routine histopathology and immunohistochemistry should be interpreted in conjunction with PCR using species-specific primers for H. meleagridis as a more accurate and specific detection tool in cases of histomoniasis.2,7 Specific primers and in situ probes have already been developed for both H. meleagridis and T. gallinarum for this purpose in other avian species.2,7 Analysis of pooled ceca and liver samples by PCR specific for the ITS region has reported higher rates of detection compared with cloacal swabs and PCR of other organs. 5 Distinguishing between the 2 organisms has implications in understanding transmission and, therefore, in biosecurity and disease prevention as well.
Further directions for this work include evaluating our novel detection PCR primers on a wider set of samples and avian species to establish their sensitivity and specificity. Investigating the relationship between H. meleagridis and T. gallinarum may also further elucidate the interaction of these organisms in instances of potential coinfections. Because most peafowl are raised on soil that likely contains H. gallinarum or its eggs, it may be a protective measure for these birds to not be housed with chickens or pheasants that may carry H. meleagridis, or to raise birds indoors to prevent contact with contaminated soil.
Footnotes
Authors’ contributions
LL Clarke contributed to conception and design of the study; contributed to acquisition, analysis, and interpretation of data; and drafted the manuscript. RB Beckstead contributed to design of the study and to analysis and interpretation of data. JR Hayes contributed to design of the study and to acquisition and interpretation of data. DR Rissi contributed to conception and design of the study and to acquisition, analysis, and interpretation of data. All authors critically revised the manuscript; gave final approval; and agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
a.
DNA extraction mini kit, Qiagen, Valencia, CA.
b.
Ready-to-go PCR beads, GE Scientific, Piscataway, NJ.
c.
SYBRsafe, Thermo Fisher, Waltham, MA.
d.
GeneJet gel extraction kit, Thermo Fisher, Waltham, MA.
e.
EZ competent cells, Qiagen, Valencia, CA.
f.
GeneJet plasmid miniprep kit, Thermo Fisher, Waltham, MA.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.