
Abstract
Oropharyngeal and cloacal swabs have been widely used for the detection of H5N1 highly pathogenic avian Influenza A virus (HPAI virus) in birds. Previous studies have shown that the feather calamus is a site of H5N1 virus replication and therefore has potential for diagnosis of avian influenza. However, studies characterizing the value of feathers for this purpose are not available, to our knowledge; herein we present a study investigating feathers for detection of H5N1 virus. Ducks and chickens were experimentally infected with H5N1 HPAI virus belonging to 1 of 3 clades (Indonesian clades 2.1.1 and 2.1.3, Vietnamese clade 1). Different types of feathers and oropharyngeal and cloacal swab samples were compared by virus isolation. In chickens, virus was detected from all sample types: oral and cloacal swabs, and immature pectorosternal, flight, and tail feathers. During clinical disease, the viral titers were higher in feathers than swabs. In ducks, the proportion of virus-positive samples was variable depending on viral strain and time from challenge; cloacal swabs and mature pectorosternal feathers were clearly inferior to oral swabs and immature pectorosternal, tail, and flight feathers. In ducks infected with Indonesian strains, in which most birds did not develop clinical signs, all sampling methods gave intermittent positive results; 3–23% of immature pectorosternal feathers were positive during the acute infection period; oropharyngeal swabs had slightly higher positivity during early infection, while feathers performed better during late infection. Our results indicate that immature feathers are an alternative sample for the diagnosis of HPAI in chickens and ducks.
Introduction
Since its emergence in 1996 in Guangdong Province, China, 37 Asian H5N1 highly pathogenic avian Influenza A virus (HPAI virus) has spread and circulated widely around the world.13,29 Outbreaks of Asian H5N1 (hereafter, H5N1) influenza have resulted in the death of millions of birds either through culling or by infection, which has caused a massive impact to the poultry sectors of affected countries. H5N1 is a zoonotic virus that has an unusually high human case fatality, 35 leading to concerns that it could evolve into a virus with significant potential for human influenza pandemic.7,8
In contrast to low pathogenic avian Influenza A virus (LPAI virus) infection, in which replication is associated primarily with the mucosa of the gastrointestinal and respiratory tracts, 25 the H5N1 HPAI virus is found in multiple organs of infected birds.2,25,31 The case fatality in chickens is ~100%,9,11,14 while in ducks it is less severe and may not even induce clinical signs,2,11,27,31 although some viruses have been reported to cause high fatality in ducks.23,28 Findings have shown that the virus is able to replicate in feather structures of infected ducks and chickens, as indicated by either the isolation of virus or the presence of viral antigen in feather structures.2,14,38,39
Several diagnostic methods, such as virus isolation (VI) and antigen and RNA detection tests, have been used to detect H5N1 virus in infected birds.21,36 Oropharyngeal and cloacal swabs are the samples recommended for detecting avian influenza viruses,17,34,36 and have been widely used for avian influenza surveillance. Oropharyngeal swabs are reported as more reliable for H5N1 HPAI virus detection,2,30 whereas cloacal swabs are more suitable for waterfowl infected with LPAI. 10 A previous study in domestic ducks showed that H5N1 HPAI virus was able to be reisolated over a longer period from feathers than from oropharyngeal and cloacal swabs. 39 Similar results were found with H7N1 HPAI virus in chickens, where the viral titer in feather pulp was higher than in swabs. 4 For diagnostic purposes, feathers also have advantages over other sample types in that they are easy to collect, and their collection is minimally invasive.5,6 Earlier findings suggest that feathers are a potential candidate as an alternative sample for the diagnosis of H5N1 HPAI virus infection.
Although some field studies have reported the use of feathers for diagnosis of HPAI infection,19,20 most provide few details on the stage of growth or body location of the feathers used. To our knowledge, there are no studies that have examined in detail the relative diagnostic value of swabs and feathers from different body locations and of varying maturity in individual chickens and ducks. The present study aimed to address this deficiency by addressing 3 objectives: 1) to compare the relative proportion of virus-positive feathers, oropharyngeal swabs, and cloacal swabs from experimentally infected ducks and chickens; 2) to compare the viral titers in these sample types; and 3) to determine the number of feathers required to confirm infection in ducks.
Materials and methods
H5N1 HPAI virus strains
Our study used 3 different HPAI H5N1 virus strains. Two were Indonesian isolates: A/duck/Sleman/BBVW-1003-34368/2007 (IDN 34368; GenBank accession CY091949) and A/duck/Sleman/BBVW-598-32226/2007 (IDN 32226; GenBank accession CY091864), which belong to clades 2.1.1 and 2.1.3, respectively. 32 The third was a Vietnamese isolate: A/Muscovy duck/Vietnam/453/2004 (VN 453), of clade 1 lineage. All viruses were propagated twice in specific pathogen–free embryonated chicken eggs. a
Animals and animal husbandry
To ensure that the animal experiments conformed to national animal welfare legislation, all protocols implemented were approved by the Commonwealth Scientific and Industrial Research Organisation–Australian Animal Health Laboratory Animal Ethics Committee. Animal experiments were performed in physical containment level 3, using appropriate personal protective equipment, including breathing air hoods. Animal rooms were ventilated with high efficiency particulate air (HEPA)–filtered air, and the temperature was set at 22°C.
Commercially sourced, healthy 7–9-week-old Pekin ducks b (Anas platyrhynchos) and 4-week-old broiler chickens c (Gallus gallus) were used in our studies. Numbered leg bands were used to identify individuals. Commercial pellets and water were provided ad libitum for both species, with vegetables and bread also provided daily to the ducks. Wood shavings, which were regularly replaced, were laid down in a corner of each room to provide a dry retreat for the birds. The duck room contained a large plastic water-filled pond, which was cleaned daily.
Feathers
Feathers from 3 different body locations (tracts) were assessed in the study: pectorosternal (representing contour feathers), primary remiges (flight feathers), and rectrices (tail feathers). Because of the absence of mature feathers in the broilers, only immature stages of these 3 feather types were assessed. However, in the ducks, we had the opportunity to compare immature and mature pectorosternal feathers. Immature feathers were distinguished from mature feathers by the absence of a formed vane and the presence of vascular pulp within the pulp cavity. 12 Only immature feathers of the 2 remaining feather types (tail and flight) in ducks were available. Flight and tail feathers were collected only after euthanasia, to minimize the welfare impact to the birds.
Study protocol
Our study utilized experimentally infected birds that were part of separate pathogenesis 31 or immune-response studies, and therefore the study design was predicated on those studies. Thirteen or 14 ducks and chickens were inoculated with each of H5N1 HPAI virus strains IDN 34368 and IDN 32226, and 5 ducks and 5 chickens were inoculated with VN 453 (Table 1).
Total number of birds and samples used in the comparison of different sample types for diagnosis of Asian H5N1 highly pathogenic avian Influenza A virus infection.*
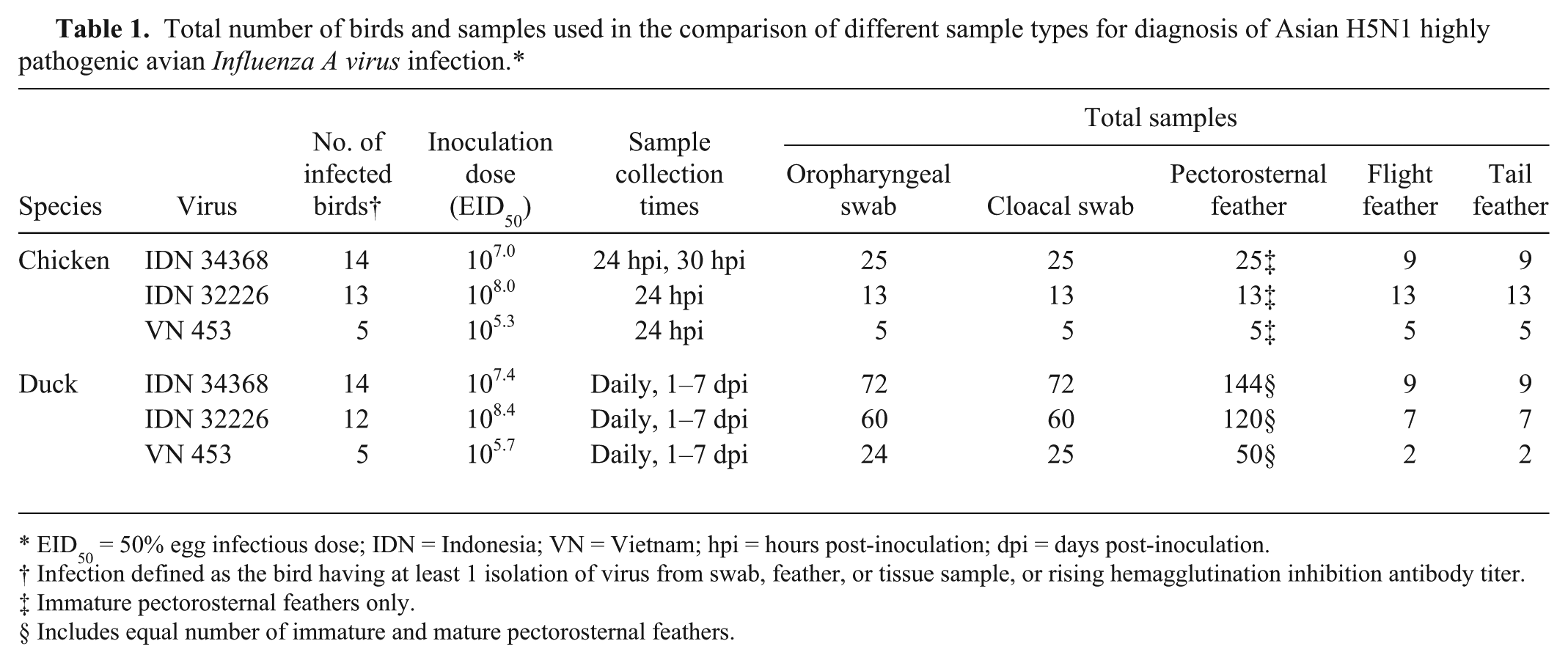
EID50 = 50% egg infectious dose; IDN = Indonesia; VN = Vietnam; hpi = hours post-inoculation; dpi = days post-inoculation.
Infection defined as the bird having at least 1 isolation of virus from swab, feather, or tissue sample, or rising hemagglutination inhibition antibody titer.
Immature pectorosternal feathers only.
Includes equal number of immature and mature pectorosternal feathers.
Ducks and chickens were inoculated by the intranasal, oral, and ocular routes with 0.5 and 0.2 mL, respectively, of diluted infective allantoic fluids, with viral doses as listed in Table 1. The birds were examined at least daily, and were euthanized as soon as moderate clinical signs were observed. In the Indonesian virus trials, 3 ducks in each group were euthanized on days 2 and 4, 2 ducks were euthanized on day 7 post-inoculation (dpi), and the remaining ducks were euthanized at day 18. In the Vietnamese virus trial, any surviving ducks were euthanized at 10 dpi.
Blood samples were collected from all chickens and ducks prior to challenge, and from surviving ducks at the termination of the trials. Oropharyngeal and cloacal cotton swabs d and pectorosternal feathers were collected prior to inoculation and daily after challenge. Flight and tail feathers were collected from most, but not all, birds after death. Because of the rapid time to death or euthanasia of chickens inoculated with IDN 32226 and VN 453, only 1 set of swabs and feathers was collected from each bird, at 24 hr post-inoculation (hpi). A second set of swabs and feathers was collected at 30 hpi from 11 chickens inoculated with IDN 34368, in addition to the initial set collected from the full group (14 chickens) at 24 hpi. A variable number of swab and feather samples was collected from each of the duck groups, depending on the initial size of the group and the number of ducks remaining on each sampling day. The total number of feather and swab samples used in the comparative study is summarized in Table 1.
To investigate the number of feather samples required to confirm infection in ducks, a further infection trial was performed. Nine 5-week-old ducks were inoculated with IDN 32226 (107.7 50% egg infectious doses [EID50] per duck), and 3 ducks were euthanized at 3, 4, and 5 dpi. Twenty immature pectorosternal feathers and 5 immature flight feathers were collected from each duck after death. Each feather was placed separately into 2 mL of transport medium consisting of phosphate buffered saline containing antibiotics (320 units/mL of penicillin, 320 µg/mL of streptomycin, and 50 µg/mL of gentamycin). In addition, lung, heart, and spleen samples were collected to confirm infection.
Processing of samples
Oropharyngeal and cloacal swabs were immersed in 3 mL of transport medium (as above), while feathers were placed into 2 mL of transport medium. Because of the varying length of feathers, the part of the feather vane exceeding the 5-mL tube e was trimmed with scissors and removed, leaving the calamus and the lower part of the rachis in the tube.
Feather, swab, and tissue samples were stored at −80°C. Thawed feather samples were ground in their 2 mL of transport medium in a mortar and pestle with purified sea sand, f poured back into the tubes, and then centrifuged at 1,000–1,300 × g at 4°C for 5 min. Results of virus testing of tissue samples have been reported previously, 31 and were used in our study only to confirm infection status of birds.
Virus and antibody detection
Virus isolation and titration was conducted in Vero (African green monkey kidney) cells as described elsewhere. 2 Vero cells were seeded into 96-well, flat-bottomed tissue culture plates g in Eagle minimal essential medium (EMEM) with 10% fetal calf serum (FCS), 100 units/mL of penicillin, 100 µg/mL of streptomycin, 0.8 mM glutamine, 0.01 M HEPES (N-2-hydroxyethylpiperazone-N-2-ethanesulfonic acid), and 2.5 µg/mL of amphotericin B. Briefly, 100 µL of each sample was diluted into 900 µL of phosphate buffered saline A (PBSA), and 10-fold dilutions made from 10−1 to 10−7. One hundred microliters of each dilution was dispensed into the Vero cell seeded tissue culture plates in 4 replicates. The first concentration included in the titration was 100 (neat) for swab samples and, because undiluted feather samples occasionally showed toxicity in Vero cells, 10−1 for feather samples. The plates were maintained at 37°C for 5 days in a humidified incubator with 5% CO2. Wells were observed for the presence of cytopathic effect, and titers were calculated according to the Reed–Muench formula. 16 Because of the different starting dilutions, the minimum detectable titers were 10−0.33 and 100.67 TCID50/0.1 mL (TCID50 = 50% tissue culture infective dose) for swabs and feather samples, respectively.
In the infection trial conducted to determine the proportion of infected feathers in ducks, VI was also conducted in embryonated chicken eggs following the method of the World Organization for Animal Health. 36 Briefly, 0.2 mL of undiluted sample suspension was inoculated into the allantoic cavity of each of three 9–11-day-old embryonated chicken eggs. The eggs were incubated for 5 days at 37°C before testing the allantoic fluid of dead or moribund embryos using a hemagglutination (HA) assay. 36 Sera from the birds were tested using the hemagglutination inhibition (HI) assay as reported elsewhere. 36 Antibody results were used to confirm the infection status of birds (see below).
Data analysis
No influenza HI antibodies were detected in any of the ducks or chickens prior to challenge, and no influenza virus was detected in any of the prechallenge samples. Infection in the inoculated birds was defined as the bird having at least 1 isolation of virus from swab, feather, or tissue sample, or rising antibody titer. Birds that did not fit this definition were considered not infected and were removed from the dataset.
Kolmogorov–Smirnov normality test was used to analyze sample distribution. Because many of the datasets were not normally distributed and the sample size of some groups was small, nonparametric tests were used to analyze the data. Data were evaluated using Mann–Whitney U test for group comparisons. To increase the robustness of statistical analysis, virus titers in chickens and ducks were each combined according to sample types to increase the number of samples. The difference was considered statistically significant if the P value was <0.05. Statistical analysis was performed using commercially available statistics software. h Calculation of 95% confidence intervals was based on the formula of Wilson (1927) as described elsewhere. 26
Results
The clinical and pathological outcomes of infection for the trials using the Indonesian viruses have been described elsewhere, 31 and the clinical and pathological signs for the Vietnamese virus were similar to those described previously for this virus. 2 All 3 viruses caused severe disease in chickens, with all birds having died or been humanely euthanized by 30 hpi. The Indonesian viruses caused no clinical signs in ducks, apart from 1 duck that died at 5 dpi with IDN 34368. 31 Four of 5 ducks infected with the Vietnamese virus developed disease, and were euthanized for welfare reasons, and the remaining duck was euthanized healthy on day 10. One duck (no. 27), inoculated with IDN 32226, was considered not infected and thus was not included in the analysis as it had not seroconverted by 7 dpi when it was euthanized, yielded no virus-positive swabs or feathers on any sampling occasion, and was negative on viral detection tests on tissues. 31 All other chickens and ducks in the study either seroconverted or yielded at least 1 virus-positive sample or tissue.
Chickens
In chickens, all sample types have very high frequency of positives
The oropharyngeal swabs, cloacal swabs, pectorosternal feathers, flight feathers, and tail feathers from almost all chickens infected with all three H5N1 HPAI viruses were positive by VI in cells at the 1 (IDN 32226, VN 453) or 2 (IDN 34368) sampling occasions before their death (Table 2; Supplementary Table 1; available at http://vdi.sagepub.com/content/by/supplemental-data). The only virus-negative samples were cloacal swabs from 3 (of 14) IDN 34368–infected chickens at 24 hpi and the tail feather sample from 1 (of 13) IDN 32226–infected chicken.
Virus isolation from swabs and feathers collected from infected chickens and ducks after inoculation with three Asian H5N1 highly pathogenic avian Influenza A virus (HPAI virus) strains.
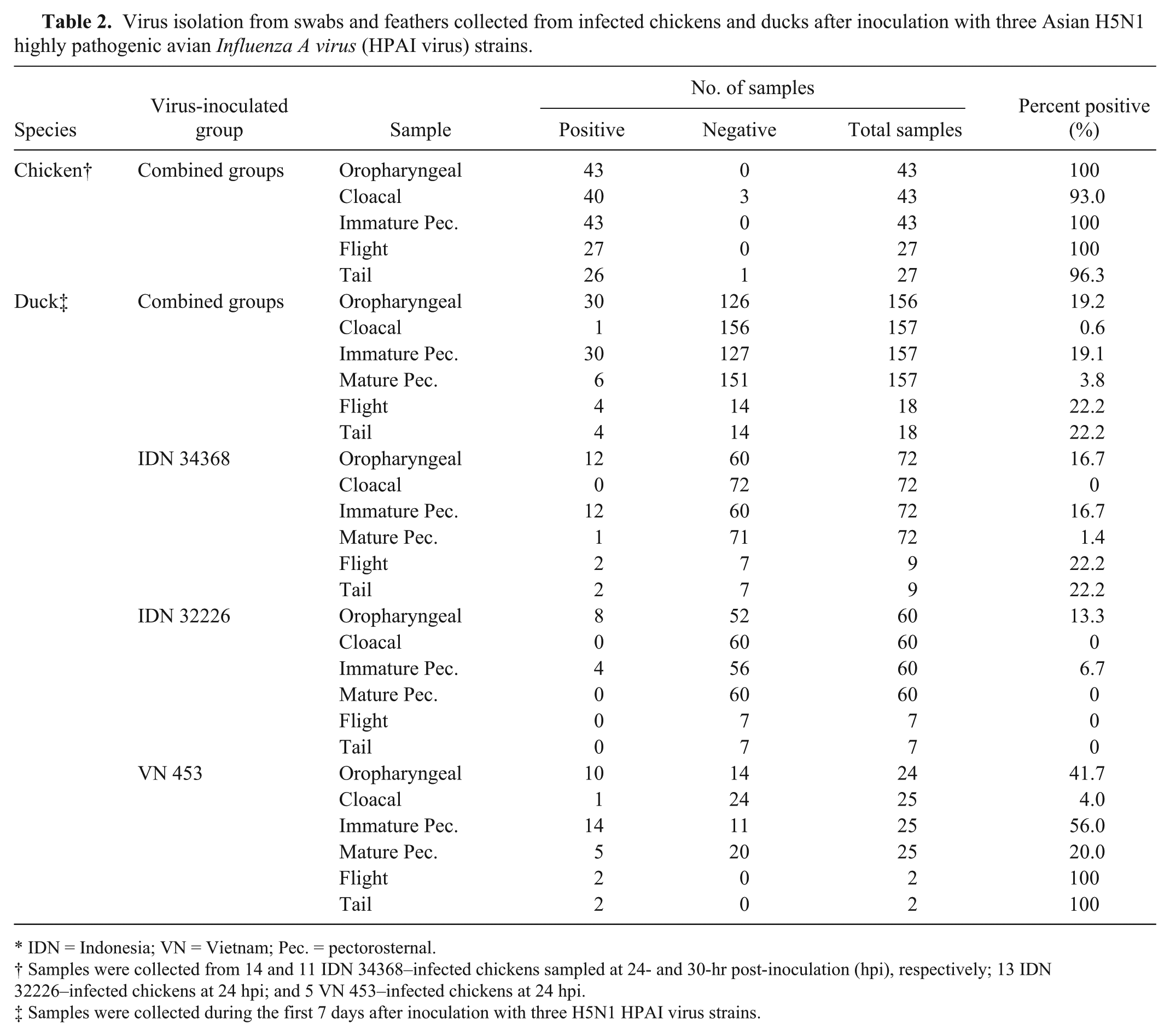
IDN = Indonesia; VN = Vietnam; Pec. = pectorosternal.
Samples were collected from 14 and 11 IDN 34368–infected chickens sampled at 24- and 30-hr post-inoculation (hpi), respectively; 13 IDN 32226–infected chickens at 24 hpi; and 5 VN 453–infected chickens at 24 hpi.
Samples were collected during the first 7 days after inoculation with three H5N1 HPAI virus strains.
Viral titers in chickens vary among sample types
Viral titers, measured in Vero cell cultures, varied among the different sample types (Fig. 1). Titer results were analyzed by individual virus groups and by data combined from the 3 virus groups. For IDN 34368, the data from the 11 chickens remaining at 30 hpi were used in the combined data analysis because the clinical stage at that time point was equivalent to that of the other 2 viruses at 24 hpi. 31 The lowest median titer (100.50 TCID50/0.1 mL) was found in cloacal swabs of the IDN 34368–infected group at 24 hpi (Supplementary Fig. 1; available at available at http://vdi.sagepub.com/content/by/supplemental-data), whereas the highest median titer (106.50 TCID50/0.1 mL) was found in both flight and tail feathers of the VN 453–infected group (Fig. 1).
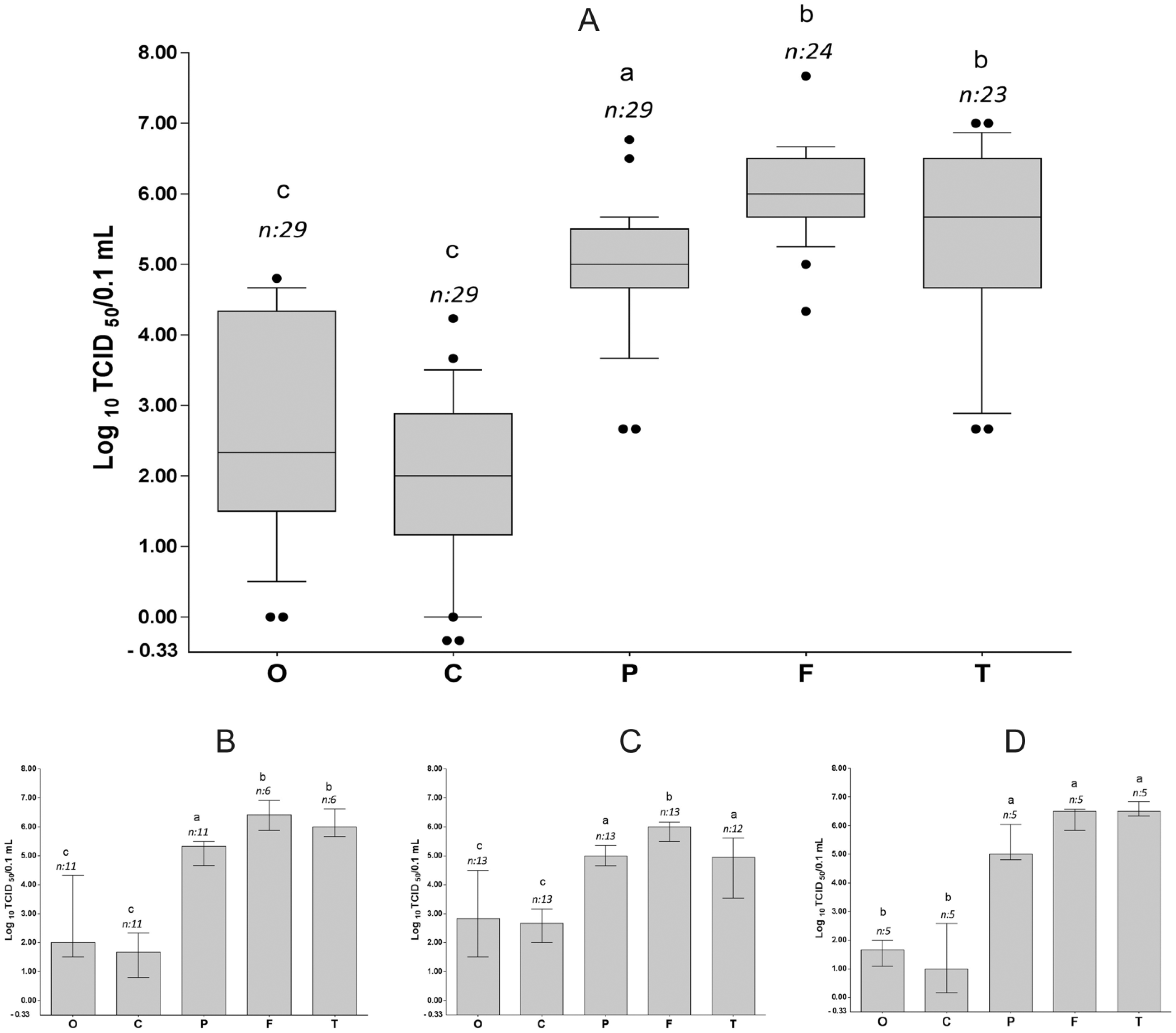
Viral titers in swabs and feathers of chickens inoculated with Asian H5N1 highly pathogenic avian Influenza A virus strains: combined data (
In all 3 individual trial datasets and in the combined dataset, the highest median virus titers were found in feathers (Fig. 1). All 3 feather types contained significantly higher virus content than either oropharyngeal swabs or cloacal swabs (P < 0.05). Among the types of feather, flight and tail feathers both had significantly higher median viral titers than pectorosternal feathers in the combined dataset and in some of the individual virus groups (Fig. 1). Between the 2 swab types, there was no significant difference in viral titer between oropharyngeal and cloacal swabs for any of the individual viruses or for the combined dataset (Fig. 1).
Viral titers in chickens increase with progression of disease
The more prolonged disease course in the IDN 34368–infected group gave us the opportunity to evaluate the change of titer during different stages of disease in chickens, with samples collected in early (24 hpi) and established (30 hpi) clinical disease. In all sample types except oropharyngeal swabs, the median titers at 30 hpi were significantly higher than in samples collected 6 hr previously (Supplementary Fig. 1; P < 0.05).
Ducks
Similar analyses using data from the individual virus-infected groups and the combined dataset were conducted on samples from infected ducks. As the infection period was longer than in chickens, samples were collected up to 7 dpi. For some analyses, data from up to 7 days were pooled.
In ducks, frequency of positives in all sample types is low
The overall proportion of virus-positive samples from ducks was markedly lower than in chickens (Supplementary Table 2; available at http://vdi.sagepub.com/content/by/supplemental-data). To allow clearer comparisons between sample types, the data from up to 7 days was pooled for each virus group and for the combined dataset (Table 2). The number of samples that gave a positive VI result was low for all sample types. From a total of 157 cloacal swabs collected over 7 days from the 3 virus groups, only 1 sample was positive (in VN 453 group at 2 dpi), and only 30 of 156 (19%) oropharyngeal swabs were positive. The overall percentage of virus-positive feather samples varied: 4% (6/157) for mature pectorosternal feathers, 19% (30/157) for immature pectorosternal feathers, and 22% (4/18) for both flight and tail feather samples. Frequently, individual ducks that were virus positive in 1 sample type were virus-negative in other sample types collected at the same time.
Sample type with highest frequency of positives is dependent on virus isolate and time of collection
The type of sample that gave the overall highest frequency of virus-positive results varied between the different virus groups. In the IDN 34368– and VN 453–infected groups, flight and tail feathers were superior to other samples, whereas with the IDN 32226 group oropharyngeal swabs were superior for positivity (Table 2). However, some of these data are based on small sample sizes, so interpretation must be guarded. Within feather types, there was again variation among the different virus groups. For both the IDN 34368 and VN 453 groups, VI-positive results were obtained with all feather types, although the number of positive mature pectorosternal feather samples was very low. For the IDN 32226 group, immature pectorosternal feathers were the only feather samples to yield positive results.
Virus positivity of the different sample types was assessed for each virus group over time (Fig. 2; Supplementary Table 2). More positive samples were detected in either swabs or feathers on 2–4 dpi than on the other days during the experiments. Early in the infection (1–3 dpi), higher proportions of oropharyngeal swabs were positive compared to feathers in the 2 groups inoculated with Indonesian viruses. During late infection (after 4 dpi) feathers were the only positive samples in all 3 virus groups. Virus was able to be isolated from feathers until 5–7 dpi in all groups (Fig. 2; Supplementary Table 2). No positive results from any sample type were obtained on the terminal days of the infections: from day 8 to 18 for IDN 34368, day 6 to 18 for IDN 32226, and on day 7 for VN 453.
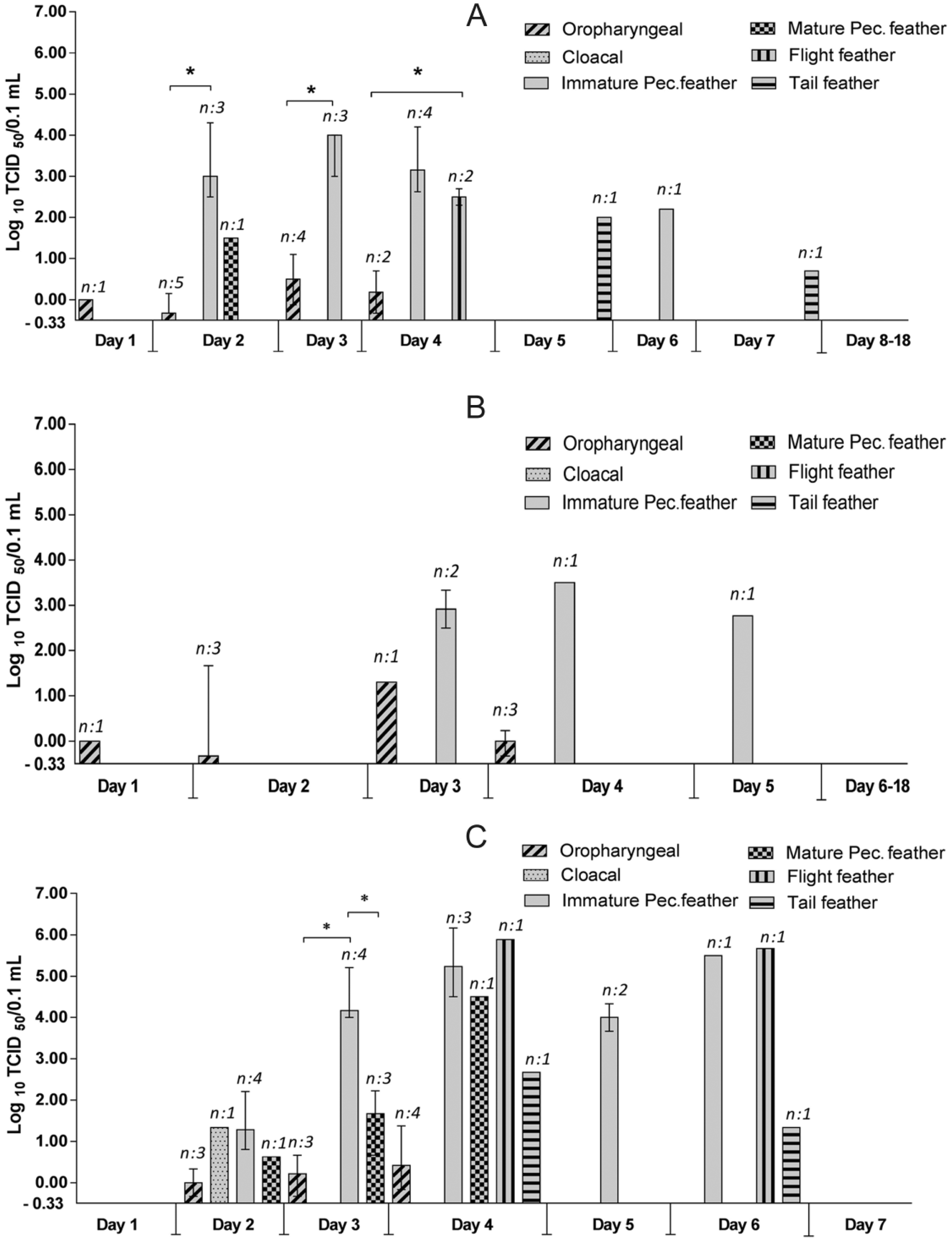
Mean viral titers over time of virus-positive swabs and feathers of ducks infected with IDN 34368 (
Viral titers in duck feathers are generally higher than in swabs
The viral titers of the different sample types were compared. When data from 7 days and all virus groups was combined, all feather types had a significantly higher median titer than oropharyngeal swabs (P < 0.05; Fig. 3A). Similarly, within the individual virus groups, most feather types had significantly higher titers than oropharyngeal swabs, with the exception of tail feathers in both the IDN 34368 and VN 453 groups where the difference was not significant (Fig. 3B–D).
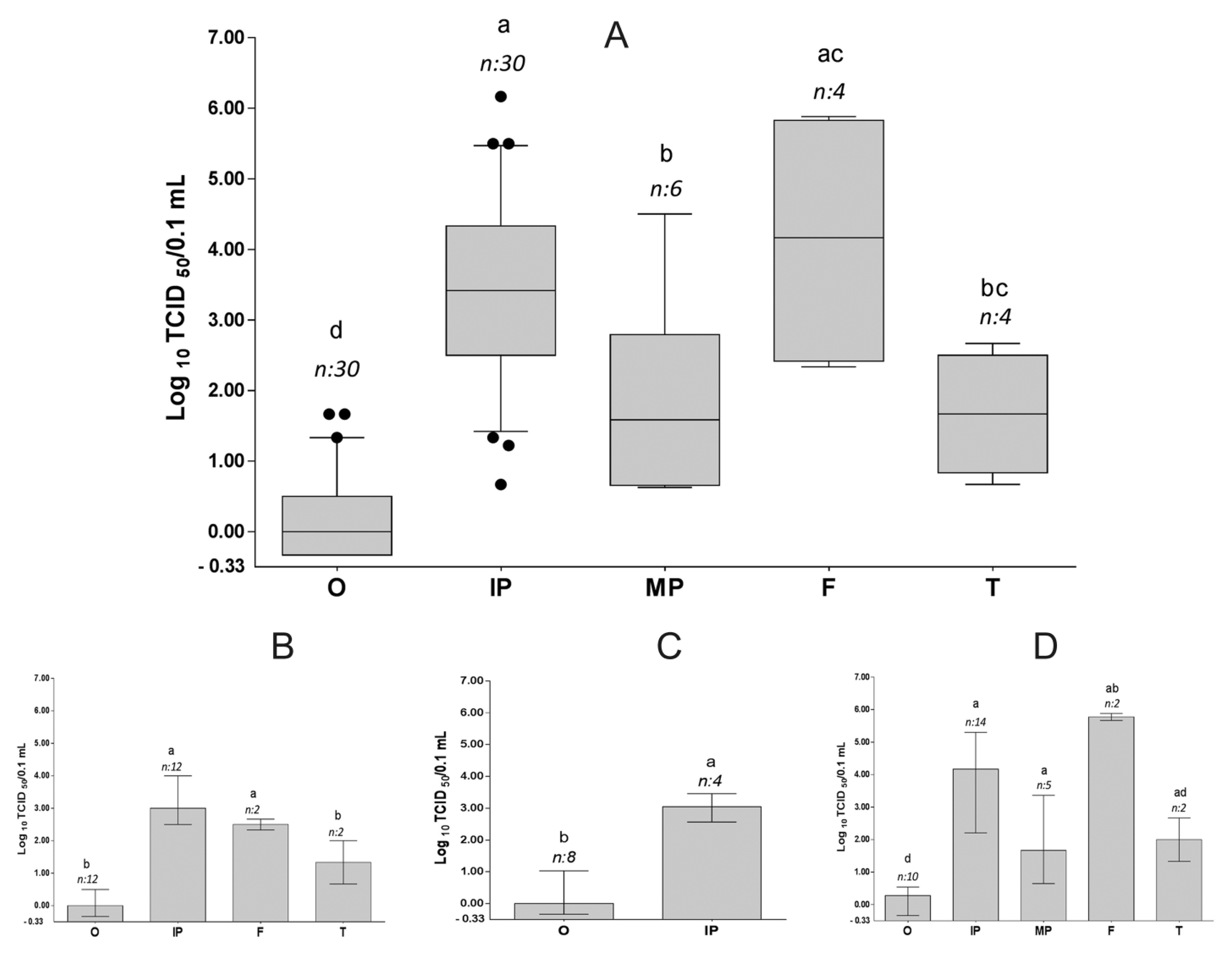
Viral titers of virus-positive swabs and feathers of ducks inoculated with three Asian H5N1 highly pathogenic avian Influenza A virus strains: combined data (top) and individual virus groups (bottom). Data from 7 days was pooled. Cloacal swabs were not included because only 1 sample from 1 virus group was positive. In the IDN 34368 and IDN 32226 groups, some sample types were not included because an insufficient number of samples were virus positive. For the top chart: the solid line inside each box is the median value; the lower and upper line of each box indicates the interquartile range; the bar shows the 10th–90th percentiles; and dots indicate data outside the 10th and 90th percentiles. For the bottom charts: each bar indicates the median viral titer with error bars showing the interquartile ranges. Kolmogorov–Smirnov test was used to determine sample distribution. Results were analyzed statistically using Mann–Whitney U test to evaluate the group differences. Letters above each box indicate significant differences (P < 0.05) between each dataset (same letter = no significant difference). Sample size (n) is indicated above each bar. O = oropharyngeal; IP = immature pectorosternal; MP = mature pectorosternal; F = flight; T = tail.
Within feather types, immature pectorosternal feathers and flight feathers tended to have higher virus titers than tail feathers and mature pectorosternal feathers, with significant and nonsignificant differences among these comparisons in the combined dataset and individual virus groups (Fig. 3). For some feather types in the individual virus groups, there was insufficient positive data to allow statistical analysis to be conducted. Statistical analysis was also not conducted on cloacal swabs because only 1 sample from all 3 virus groups was positive.
The number of feathers from ducks required to obtain a positive result
The results described above indicated that although duck feathers have relatively high titers of virus, the number of feather samples that test positive by VI is relatively low. This is in contrast to infected chickens, where nearly all feather samples from all 3 virus groups contained detectable virus. This raised the question of how many feather samples would be needed to confirm infection in ducks. To help answer this question, a separate infection study was conducted in ducks, using IDN 32226. This virus was selected because, of the 3 viruses used in the initial trials, it produced the lowest proportion of positives in feathers. Thus, the number of feathers required for diagnosis with this virus would likely be an overestimate of the number required for many other H5N1 viruses. In addition, this virus is representative of the most prevalent HPAI present in Indonesia. 32
Twenty immature pectorosternal and 5 immature flight feathers were collected after euthanasia from 3 IDN 32226–infected ducks on 3, 4, and 5 dpi, to test for virus by culture in embryonated eggs and Vero cells. All ducks were confirmed as infected by the isolation of virus from tissues taken at autopsy (data not shown).
Combining data from 9 ducks at the 3 time points, 10% of pectorosternal and 22% of flight feathers were positive on VI in eggs (Table 3). No feathers of either type from 1 duck at 3 dpi (no. #89) or 1 duck at 5 dpi (no. 95) were positive in either culture system (Supplementary Table 3; available at http://vdi.sagepub.com/content/by/supplemental-data). Of the remaining ducks, flight feathers had higher values for positivity than pectorosternal feathers in all but 1 duck. Both feather types had highest positivity values on 5 dpi (Table 3). The highest proportion of positives was from flight feathers of duck 92 at 5 dpi, when 3 of 5 feathers (60%) were virus positive (Supplementary Table 3).
Proportion of virus isolation–positive feathers in ducks inoculated with an Asian H5N1 highly pathogenic avian Influenza A virus strain (IDN 32226).*

IDN = Indonesia; dpi = days post-inoculation.
Data from 3 ducks at each time point.
Using methodology of Cannon and Roe (1982), as described elsewhere, 26 we calculated the number of feathers needed in a sample to correctly diagnose a duck as positive for HPAI infection, based on prevalence of positive feathers ranging from 5% to 60% (Supplementary Table 4; available at http://vdi.sagepub.com/content/by/supplemental-data). According to this method, and using prevalence of 5% and 20%, respectively, 24 pectorosternal and 6 flight feathers were required per duck to ensure a confidence level of 0.7. For a confidence interval of 0.95, this was 59 and 14 feathers per duck for pectorosternal and flight feathers, respectively.
Discussion
The accurate diagnosis of HPAI infection is essential to enable rapid and appropriate control measures to be undertaken. A range of diagnostic tests is available to detect H5N1 HPAI viruses in samples from infected birds, but a key component of effective diagnosis is the choice of sample to be tested. We evaluated the use of feathers for the diagnosis of H5N1 HPAI in ducks and chickens of known infection status, and compared feathers to oropharyngeal and cloacal swabs, which are the conventional samples used for H5N1 diagnosis. We used VI in Vero cell cultures to determine the proportion of samples that were virus positive and the titer of virus in each of these sample types. In addition, we estimated the number of feathers that would be required to confirm infection in ducks.
The potential use of feathers for the detection of H5N1 HPAI virus has been investigated previously. Virus was reisolated from feathers in an infection study using 2 H5N1 HPAI viruses in ducks and geese, 39 and virus was also detected in feathers of infected wild swan, 40 and of chickens, quail, turkey, pheasant, and chukar partridge. 14 H5N1 HPAI virus was still infective in feathers of infected ducks stored for >5 months at 4°C. 41 Similarly, feather pulp of infected chickens was a potential sample for detection of H7N1 HPAI virus. 4 However, little was known about the type and stage of growth of feathers that can be used as a sample for detection of H5N1 HPAI virus in both chickens and ducks. Moreover, there was no information on the use of feathers for diagnosis of Indonesian H5N1 HPAI viruses, which reportedly cause less severe disease in ducks than other strains of the virus. 2 Therefore, in our study, chickens and ducks were infected with 2 Indonesian H5N1 HPAI viruses of different subclades. A Vietnamese H5N1 HPAI virus was used as a comparison, as the virus has been reported to cause severe disease in ducks. 2 Different types of feathers were evaluated to assess the advantages and disadvantages of feathers as diagnostic samples. In addition, 2 stages of growth (immature and mature) of 1 feather type (pectorosternal) in ducks were compared.
In chickens infected with either of the Indonesian viruses or the Vietnamese virus, clinical signs occurred rapidly and were severe, resulting in death or humane euthanasia within 30 hpi. All sample types (oropharyngeal swabs, cloacal swabs, pectorosternal feathers, flight feathers, and tail feathers) had very high positivity. From a total of 183 samples collected from 32 H5N1-infected chickens, only 4 samples were virus negative (3 cloacal swabs and 1 tail feather sample). The viral titers varied among the sample types, with the highest titers found in feather samples and the lowest titers in cloacal swabs. We determined that the titer of virus in all feather types and in cloacal swabs increased as the disease course progressed. Overall, we demonstrated that in chickens, H5N1 HPAI diagnosis can be readily achieved using a variety of sample types and that feathers provide a very suitable alternative sample to the routinely collected oropharyngeal and cloacal swabs. The ease of collection of feathers, the lack of need to purchase swabs (which is an important consideration in some countries), and the higher viral titers found in feathers provide some advantage over the conventional sample types.
The clinical outcome and the frequency of virus-positive samples of H5N1 HPAI virus infection in ducks in our study was markedly different to that in chickens, particularly with the 2 Indonesian viruses. All but 1 of the 26 ducks infected with the Indonesian viruses remained healthy for the duration of the study. Although 4 of 5 ducks infected with the Vietnamese virus developed moderate disease signs and had to be euthanized from 3 to 6 dpi, the disease course was more protracted than and not as severe as in chickens. A striking difference between infected chickens and ducks was the paucity of virus-positive samples collected from ducks, even from the more severely affected Vietnamese virus group. From a total of 663 swab and feather samples collected during acute infection from 31 ducks infected with the three H5N1 viruses, only 75 samples were positive on VI. This low sample positivity from ducks known to be infected with H5N1 virus demonstrates clearly that the diagnosis of H5N1 HPAI in ducks of unknown infection status is a major challenge.
Cloacal swabs were the sample type with the lowest positivity from ducks, with only 1 of 157 swabs being virus positive. This suggests that H5N1 HPAI virus replicates poorly in the intestinal tract of ducks, as has been reported by others.3,24 This contrasts to LPAI viruses in ducks, which replicate in the intestinal tract and associated tissues, resulting in the shedding of LPAI viruses from the cloaca and the ready detection of virus in cloacal swabs. 33 With the H5N1 HPAI virus strains in our study, testing of cloacal swabs from ducks was clearly unproductive, and we suggest that this is likely to be the case for the diagnosis of H5N1 HPAI in the field.
The overall percentage of virus-positive results in oropharyngeal swabs and 3 types of feather samples (immature pectorosternal, flight, and tail) from ducks was ~20%. The type of sample that gave the highest positivity varied slightly among the 3 virus groups and varied over time. In general, oropharyngeal swabs gave superior detection early in the course of infection (1–3 dpi) whereas feathers were the only sample type that yielded a positive result after 4 dpi, with feather samples from some ducks remaining positive up to 7 dpi. Others have also reported that H5N1 virus was only detected in swabs in the early stages of infection in ducks15,18 and that virus can be detected in feathers up to 8 dpi 38 or 10 dpi 1 . In contrast to the relatively low proportion of positive samples from ducks tested in our study, another study found that a high proportion of swab, and wing, breast, and tail feather samples from 4-week-old ducks from 3–10 dpi were virus positive by real-time reverse transcription polymerase chain reaction. 1 However, that study found similar low rates of positivity to our study when feather and swab samples from older ducks (24-week-old) were tested. In that study, the H5N1 virus used appeared to be more pathogenic in ducks and this may explain its difference with the Indonesian viruses we used in our study, which were generally asymptomatic in ducks.
We commonly found individual ducks that were virus positive in 1 sample type and negative in other sample types collected at the same time, and the sample type that was positive in that individual changed on different sampling days. Thus, we suggest that when veterinarians are faced with ducks of unknown infection status and of unknown duration of infection, a combination of oropharyngeal swab and feather samples would maximize the probability of virus detection. However, even with this combination of sample types, it must be recognized that the likelihood of an infected duck testing negative in both samples remains a strong possibility.
As with the chicken groups, feather samples from the duck groups tended to have higher median viral titers than oropharyngeal swab samples. Within the different feather types, immature pectorosternal feathers and flight feathers generally had higher median titers than tail feathers or mature pectorosternal feathers. Considering that immature pectorosternal feathers also had the highest positivity in some of the virus-infected duck groups, it would appear that this feather type would be the sample of choice for diagnosis in ducks. In addition, it is practical to collect this feather type from live birds with minimal discomfort, whereas collection of the other potential sample candidate, flight feathers, may cause undue damage to the bird because of flight feathers’ large size and rich vascularization, which can result in bleeding on plucking. Flight feathers are better collected after death to prevent these welfare concerns.
Considering the comparatively high viral titers in duck feathers but the overall low frequency of virus-positive samples, we asked the question of how many feathers would need to be collected from an infected duck to confirm infection. We focused on immature pectorosternal feathers and flight feathers because these 2 feather types showed most promise as alternative diagnostic samples. Using a virus that had provided a relatively low positivity in the initial trials, we found that ~24 pectorosternal feathers and 6 flight feathers would need to be tested per duck to obtain a positive diagnostic result with a confidence level of 0.7 (Supplementary Table 4). The number of feathers required for diagnosis naturally increased as the confidence level increased.
The high titers of H5N1 virus in feathers of both ducks and chickens reflect the tropism of the virus for feather tissues.38,39 H5N1 viral nucleoprotein antigen has been detected principally in vascular structures of the feather pulp of H5N1-infected chickens 14 and in feather epithelium in H5N1-infected ducks. 39 In ducks, we were able to compare virus content in mature and immature pectorosternal feathers. In all 3 virus groups, immature feather samples had a higher proportion of positive results than mature feathers, and the median viral titer of immature feathers in the combined dataset was higher than that of mature feathers. In mature feathers, the main feather structures are non-living tissues, and the blood vessels are atrophied at this stage of feather development. 22 Thus, virus is likely less able to spread to and replicate in mature feathers compared to immature feathers. Further studies to investigate the distribution of virus within feather structures would increase understanding of the effect of HPAI infection on feathers and their potential use as diagnostic samples.
In this study, we tested 3 H5N1 HPAI viruses, all of which are known to replicate systemically when infecting ducks and chickens.2,31 It is likely that other HPAI viruses would also exhibit tropism for feather structures, and therefore would be detectable in feather samples. 4 However, for LPAI viruses, which generally do not replicate systemically, swabs would remain the preferred sample for detection of infection.
We used VI in Vero cell cultures as the diagnostic test to determine presence of H5N1 virus and to measure the concentration of virus in the samples. In this study, we needed an assay that was quantifiable, practical, inexpensive, and comparable across different sample types. Also, we needed a test that was not too sensitive to contaminants. Although a molecular test may have provided a more sensitive assay, we were concerned that molecular assays would detect irrelevant viral RNA, for example, from contamination of samples with inactivated virus present in the environment. In addition, viral RNA is highly persistent in birds during convalescence, 30 and for this reason, we believe that polymerase chain reaction tests are not accurate for detection of current infection status in individual birds. Popularity of use in the field was not a consideration for the choice of detection assay, bearing in mind that we needed to compare sample types, not assay procedures. For these reasons, we decided that isolation of infectious virus was the more robust and defendable assay to use for this purpose.
In conclusion, we have demonstrated that feathers can be used as effective samples in the diagnosis of Indonesian and Vietnamese H5N1 HPAI viruses in ducks and chickens. Feathers had significantly higher titers than either oropharyngeal or cloacal swabs in both species. In chickens, H5N1 virus could be detected in all swab and feather samples from nearly every bird so if circumstances warranted, feathers could provide an additional or alternate sample type to be collected from suspect infected birds. In ducks, only a small proportion of all sample types was virus positive, even with the Vietnamese H5N1 virus that caused moderately severe disease in the birds. In particular, it was clear that cloacal swabs are of little use in the diagnosis of H5N1 infection in ducks. Because of the generally low proportion of virus-positive samples in ducks, we suggest that multiple sample types comprising oropharyngeal swabs and pectorosternal or flight feather samples should be tested to maximize the likelihood of obtaining a virus-positive result. However, veterinarians should be aware that even with this approach, it is possible that the infection status of individual H5N1-infected ducks may be incorrectly assigned. For this reason, we also suggest that samples be taken from several ducks of the flock to maximize the chance of virus detection at a flock level.
Footnotes
Acknowledgements
We are grateful to the staff of the AAHL Animal Studies team (Geelong, Victoria, Australia) for assistance with animal trials and to Dr. Frank Wong for critically reviewing the manuscript.
a.
Australian SPF Services, Woodend, Victoria, Australia.
b.
Triple S Farming Pty Ltd., New Gisborne, Victoria, Australia.
c.
Steggles, Bannockburn, Victoria, Australia.
d.
Eurotubo cotton swab, Deltalab, Rubi, Spain.
e.
5-mL yellow cap sterile container, Sarstedt Australia Pty. Ltd, Mawson Lakes, South Australia, Australia.
f.
Merck Pty Ltd., Victoria, Australia.
g.
NuncF96 MicroWell plates, Thermo Fisher Scientific Australia Pty Ltd., Victoria, Australia.
h.
GraphPad Prism 5, GraphPad Software Inc., La Jolla, CA.
Authors’ contributions
H Nuradji contributed to design of the study; contributed to acquisition, analysis, and interpretation of data; and drafted the manuscript. J Bingham contributed to conception and design of the study, and contributed to acquisition, analysis, and interpretation of data. S Lowther and H Wibawa contributed to acquisition and analysis of data. A Colling contributed to analysis of data. NT Long contributed to acquisition of data. J Meers contributed to design of the study, and contributed to analysis and interpretation of data. All authors critically revised the manuscript; gave final approval; and agree to be accountable for all aspects of the work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was partially funded by the Australian Centre for International Agricultural Research (ACIAR), project number AH/2004/040. H. Nuradji was supported by a John Allwright Fellowship funded by ACIAR.