
Abstract
Highly pathogenic avian influenza (HPAI) subtype H5N1 is an infectious systemic viral disease that results in high morbidity and mortality in poultry, and has been reported in a wide range of wild bird species during the last few years. An outbreak of HPAI H5N1 occurred in wild birds in Sweden in 2006 that affected several duck species, geese, swans, gulls, and raptors. Tufted ducks (Aythya fuligula) accounted for the largest number of positive cases and, therefore, were selected for more in-depth histologic and immunohistochemical evaluations. The main histologic lesions associated with the presence of avian influenza antigen were found in the brain, pancreas, and upper respiratory tract. Other tissues in which influenza antigen was variably found included liver, lung, adrenal glands, kidneys, and peripheral nerve ganglia. The current study describes the pathology and viral tissue targeting of H5N1 by using histology, polymerase chain reaction, and immunohistochemistry, and highlights the range and variation in the presentation of the natural disease in tufted ducks.
Introduction
Influenza A viruses (family Orthomyxoviridae) naturally infect a broad range of avian and mammalian species, including humans. Subtypes H5 and H7 have the ability to become highly pathogenic and cause generalized infection and high mortality in poultry. 23,28 Up until 2002, only sporadic cases of infection with highly pathogenic avian influenza (HPAI) had been reported in free-living wild birds. The HPAI cases were all associated with infections in domestic fowl, 23 except for an outbreak in common terns (Sterna hirundo) in South Africa in 1961, which was caused by H5N3 HPAI. 1 In 2002, HPAI virus of the subtype H5N1 spread and caused clinical disease and death among captive waterfowl and other wild birds in Hong Kong, 6 and is considered to be the first documented outbreak of an epidemic succession that affected many countries in Asia, Africa, and Europe (World Organization for Animal Health: Avian influenza. Facts and figures: H5N1 timeline. Available at: http://www.oie.int/Eng/info_ev/en_AI_factoids_H5N1_Timeline.htm. Accessed on April 1, 2009).
In 2005, an outbreak of H5N1 with mortality in several species of waterfowl, predominantly bar-headed geese (Anser indicus), occurred in Lake Qinghai in the Province of Qinghai in western China. 12 The lake is a breeding center for migrant birds from southeast Asia, Siberia, Australia, and New Zealand. 12 It is postulated that migrating wild birds could have spread the virus from east Asia, through southern Siberia, and further into Africa and Europe, which resulted in outbreaks in 2005 and 2006. 13 However, the theory regarding wild birds spreading the virus is debated and needs to be further confirmed. In February 2006, HPAI H5N1 outbreaks occurred in wild-living birds in many European countries simultaneously. The first confirmed case in northern Europe occurred in a mute swan (Cygnus olor) on the German island of Ruegen 27 in the Baltic Sea, situated about 400 km south of the Swedish coast, and, 3 weeks later, the first case of HPAI H5N1 infection was detected by the National Veterinary Institute (SVA; Uppsala, Sweden) and confirmed by the Community Reference Laboratory (CRL; VLA Weybridge, UK) in 2 tufted ducks (Aythya fuligula) from Oxelösund on the east coast of Sweden. During the following 8 weeks, the infection spread along the coast northward up to Stockholm and southward down to the Blekinge archipelago (between latitudes 55°N and 60°N), which involved a coastal area of approximately 900 km. During this outbreak, infection was confirmed in avian species of the taxonomical orders Anseriformes: tufted duck, scaup (Aythya marila), smew (Mergus albellus), goosander (Mergus merganser), mute swan, Canada goose (Branta canadensis); Charadriiformes: herring gull (Larus argentatus); Falconiformes: common buzzard (Buteo buteo); and Strigiformes: European eagle owl (Bubo bubo); and in a wild mink (Mustela vison). 29 In Sweden, as in Denmark, 2 tufted ducks accounted for the largest number of identified positive cases.
Disease caused by experimental HPAI H5N1 infection has been studied in many avian species, including swans, 3,8 geese, 3,17 passerines, and budgerigars (Melopsittacus undulatus), 7,20 Pekin ducks (Anas platyrhynchos domestica), 15,26 several wild ducks, 9 several gallinaceous species, 18 and others. Experiments indicate that pathologic changes and susceptibility vary between species and within species. 3,4,7–9,15,17,19 Information about the pathology of spontaneous infection by HPAI H5N1 in wild birds is scant, limited to a few species and/or small numbers of birds. Previous publications include descriptions in swans in Europe, 14,25 large-billed crows (Corvus macrorhynchos) in Japan, 24 magpies (Pica pica sericea) in South Korea, 11 tree sparrows (Passer montanus) in China, 10 wild waterfowl, a grey heron (Ardea cinerea), and a black-headed gull (Larus ridibundus), 6 as well as bar-headed geese 12 in China. The more limited descriptions are probably because of difficulties in obtaining fresh carcasses suitable for pathology investigations and because mortality caused by HPAI H5N1 in wild birds does not appear to occur in massive numbers, as is the case in outbreaks in poultry.
The current study describes the pathology and distribution of virus in tissues of tufted ducks naturally infected with HPAI H5N1 virus in Sweden. The purposes of the study were to obtain a better understanding of the disease in a frequently affected wild bird species and to discuss the diagnostic implications.
Materials and methods
Outbreak
Postmortem examination and sampling. During the avian influenza outbreak in Sweden in 2006, routine necropsies and sampling for the possible presence of H5N1 were carried out on 83 tufted ducks in biosafety level 3+ facilities at SVA. Cloacal and/or tracheal swabs were obtained for polymerase chain reaction (PCR) screening for influenza virus from all birds. At the beginning of the outbreak, only cloacal swabs were used, because they were considered to be the most reliable samples for influenza screening (based primarily on what was known about low pathogenicity avian influenza). Cloacal swabs were obtained by inserting a cotton swab through the vent opening and swabbing the mucosal lining. Two weeks after the first positive bird was identified, tracheal swabs were substituted for cloacal swabs, because this was considered to be a more reliable sampling method for HPAI. Tracheal swabs were obtained by inserting a cotton swab through the glottis and swabbing the inside of the trachea. Selected tissue samples were stored frozen (–20°C) or fixed in 10% neutral buffered formalin for histopathology and immunohistochemistry (IHC). The number of tissue samples varied depending on the condition of the bird (degree of autolysis and amount of scavenging). Body condition was estimated based on the amount of body fat and musculature.
PCR analyses for influenza virus. RNA from tissue samples, tracheal swabs, and cloacal swabs was extracted by using a virus mini extraction kit a in an extraction robot. b Extracted RNA was analyzed for the presence of influenza A viruses by a real-time reverse transcription (RT) PCR assay (matrix [M] gene 22 modified by the CRL). All positive cases were further examined with an H5-specific RT-PCR (KHA, hemagglutinin [HA] gene). 21 Both PCR assays were performed according to the recommendations from the CRL. The positive H5 PCR products, covering part of the HA gene, including the H0 cleavage site, were sequenced, and the cleavage site was analyzed to determine the pathogenicity of the viruses.
Of the 83 sampled tufted ducks, 37 tested positive for HPAI. Three birds (2 of which were not included in the present study) were negative on PCR analysis of the initial tracheal swab but had similar encephalic and pancreatic histologic lesions as positive birds. Therefore, PCR analysis was conducted on brain samples from these birds by using the same method as described for the swabs.
Sample selection
For the present study, 20 tufted ducks were selected of the 37 that tested positive for the presence of HPAI viral RNA. Inclusion criteria for the study were low degree of autolysis, detection of HPAI nucleic acid by PCR in at least one sample (swab or tissue), and availability of brain and at least one other organ for histopathologic investigation. Of the 20 birds included in the study, 2 were sampled by cloacal swabs, 17 were sampled by tracheal swabs, and 1 was sampled by both cloacal and tracheal swabs for PCR analysis (Table 1). The selected ducks included 9 females, 8 males, and 3 whose sex was not recorded at necropsy. Eleven were adults, and 2 were juveniles, and, in 7, the estimated age was not recorded. Thirteen ducks were found dead, and 7 ducks were euthanized because of severe neurologic signs, including head tilt, circling, loss of balance, and drooping wings. Tissues available for histopathology from the selected ducks included brain (n = 20), lungs (n = 17), upper respiratory tract (n = 17), air sacs (n = 7), heart (n = 14), liver (n = 17), spleen (n = 14), kidneys (n = 15), pancreas (n = 14), adrenal glands (n = 10), intestines (n = 14), gonads (n = 10), and ventriculus (gizzard) and proventriculus (n = 9). Brain, lung, liver,
Diagnostic tests for avian influenza in 20 selected tufted ducks. *
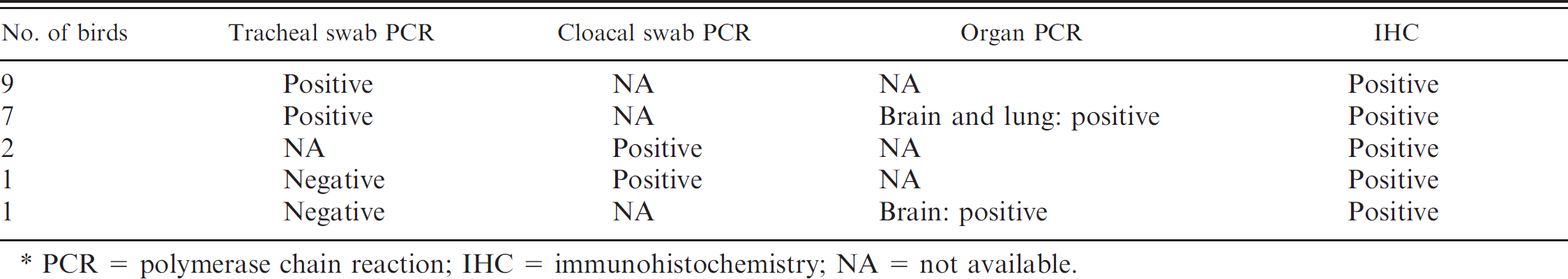
PCR = polymerase chain reaction; IHC = immunohistochemistry; NA = not available.
kidney, pancreas, and intestines were also stored at −20°C for possible additional virologic analyses.
Histopathology and IHC
After formalin fixation, the tissue samples were processed routinely, sectioned at 4–5 μm, and stained with hematoxylin and eosin. Duplicate sections for IHC were mounted onto glass slides treated with adhesive, c depar-affinized in xylene, and rehydrated. Endogenous peroxidase activity was blocked with 3% hydrogen peroxide for 7 min. The sections were treated with proteinase K d for 6 min and with 2% bovine serum albumin e for 20 min. They were then incubated at room temperature for 60 min with a commercial anti-influenza A nucleoprotein primary monoclonal antibody, f diluted 1:200. The detection was conducted with labeled streptavidin-biotin. g The color was developed with aminoethylcarbazole substrate. h Sections were counterstained with hematoxylin, coverslipped, and examined microscopically. Each immunostain included a positive reference control (pancreas from a swan that was confirmed positive for H5N1) and a negative control (pancreas of a healthy chicken). Each section was also accompanied by a primary antibody-omitted control. The presence of viral antigen was described for each tissue. The intensity and extension of the immunostaining was assessed semiquantitatively (negative, mild, moderate, or marked).
Results
Gross pathology
Most of the ducks were in good body condition (n = 14), 3 were in poor condition, and 1 was emaciated. Body condition had not been recorded in 2 ducks. Few gross lesions were noted. The most prominent gross lesions were mildly to moderately congested lungs (n = 8), red-brown mottling of the pancreas (n = 6; Fig. 1), and moderately enlarged spleens (n = 3). The gastrointestinal tract was empty in 5 birds, whereas the rest of the birds had small amounts of digesta in the intestines.
Histopathology and IHC
The main histologic lesions associated with the presence of avian influenza antigen were found in the brain, pancreas, and upper respiratory tract sections. All 20 birds were positive by IHC in at least 1 tissue.
Brain and other neural tissue. Most birds (18/20) had a nonsuppurative encephalitis or meningo-encephalitis. There were multifocal areas of gliosis, neuronophagia, and occasional lymphocytes, as well as mild-to-moderate perivascular cuffs, with a predominance of macrophages and lymphocytes in both the gray and white matter in most regions of the brain. Neurons within these areas had degeneration or necrosis. Abundant viral antigen was present in the nucleus and cytoplasm of neurons, in macrophages, and in glial cells, mostly in association with inflammatory lesions (Fig. 2). In 2 ducks, there was positive influenza staining of ganglion cells peripheral to the adrenal gland.
Upper respiratory tract. In the nasal mucosa, a mild intraepithelial infiltration of heterophils, especially in the lamina propria and deeper parts of the epithelium, was observed in several ducks (12/17). There were occasional degenerate epithelial cells in the inflamed areas, but necrosis or severe changes were rare. Blood vessels often were congested and had swollen endothelial cells. Fibrin thrombi were variably present in small vessels in the lamina propria mucosae. Influenza viral antigen was detected both in the superficial and deeper parts of the nasal epithelium (8/17; Fig. 3), as well as in the endothelium and media of some of the smaller vessels in the mucosa (2/ 17). The trachea of 3 ducks had a mild infiltration of heterophils and lymphocytes in the submucosa, but no viral antigen was detected.
Lungs and air sacs. The lungs had marked congestion and hemorrhages. Fibrin thrombi were occasionally observed in smaller vessels and capillaries. Inflammatory changes were observed in the lungs of more than half of the cases (11/17) and ranged from mild infiltration of heterophils in parabronchi to severe inflammation, with necrofibrinous exudate and sloughed necrotic epithelial cells into the lumen of parabronchi, atria, and air capillaries. Parabronchial epithelium was occasionally hyperplastic, and there were large numbers of heterophils and moderate numbers of mononuclear cells expanding the interstitium. Viral antigen, which was localized to air vesicles and squamous epithelium lining parabronchi, was only observed in 18% of the birds (3/17). In secondary bronchi, infiltration of lymphocytes and mononuclear cells was occasionally observed in the lamina propria.
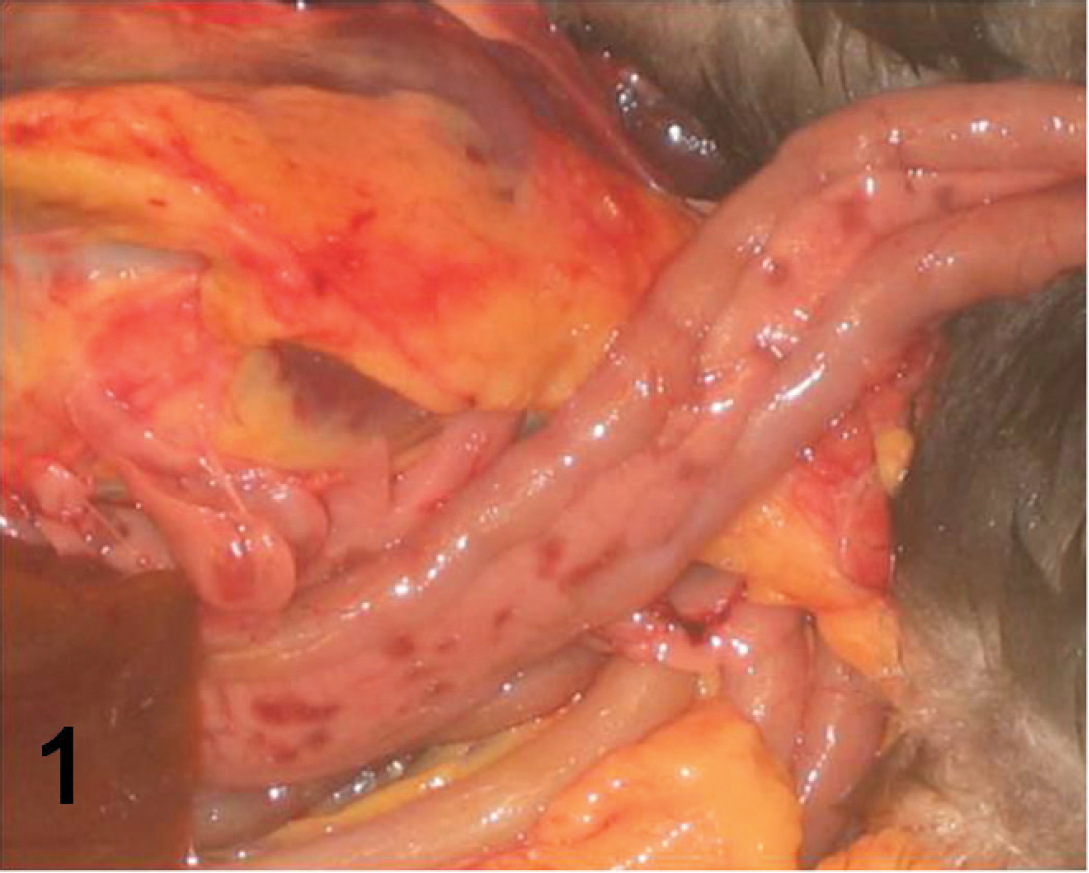
Pancreas of tufted duck (Aythya fuligula), showing multifocal areas of hemorrhage.

Brain: influenza viral antigen (red stain) in nucleus and cytoplasm of neurons. Influenza A immunohistochemistry. Hematoxylin counterstain. Bar = 50 μm.

Nasal mucosa: influenza viral antigen in superficial epithelium and submucosa. Immunohistochemistry. Hematoxylin counterstain. Bar = 50 μm.

Liver: area of hepatic necrosis with influenza viral antigen in nucleus and cytoplasm of hepatocytes. Immunohistochemistry. Hematoxylin counterstain. Bar = 50 μm.
Fibrin and cell-rich exudates were seen in the lumen of these airways. Positive IHC staining was not observed in the secondary bronchi or in any of the air sacs that were examined. Inflammatory lesions were not observed in the air sacs.
Liver. Influenza antigen was found in the nuclei and cytoplasm of hepatocytes in 3 of 17 ducks (Fig. 4). These livers had random, focally extensive areas of hepatic necrosis. Moderate numbers of heterophils and lymphocytes were associated with the necrosis.
Pancreas. Pancreatic lesions were dominated by severe, acute, multifocal-to-coalescing coagulative necrosis of the acini (11/14). The necrotic areas were often associated with hemorrhage and were surrounded by varying amounts of mixed inflammatory cells. When multiple samples of pancreas from single individuals were studied, the extension of the necrosis varied from just detectable to more than 50% of the section. Necrohemorrhagic lesions in the pancreas were observed in all but one of the tufted ducks (19/20), but viral antigen was only found in 6 ducks (43%). The antigen was present in the nuclei and cytoplasm of acinar cells, mostly in a rim of cells that bordered the necrotic areas (Fig. 5).
Urinary system. A mild, interstitial infiltration of mononuclear cells was noted in the kidney of 1 of 14 ducks, and focal degeneration and necrosis of renal
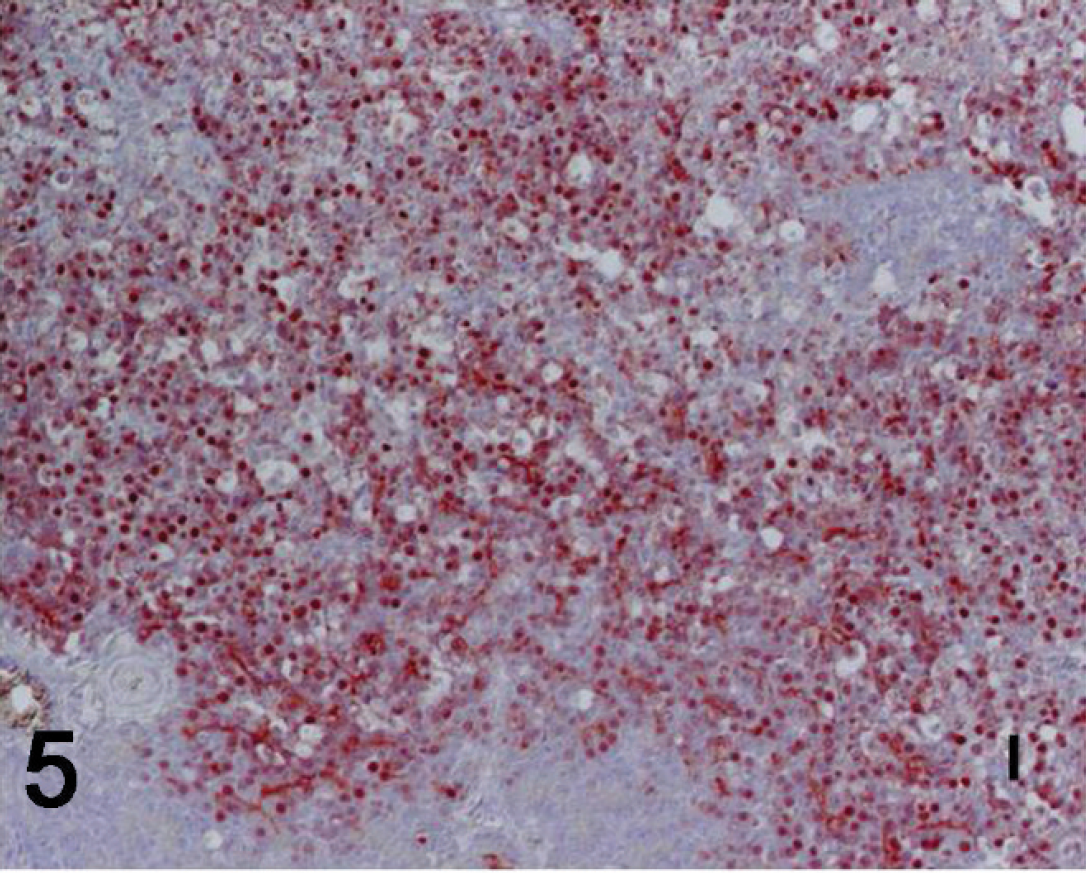
Pancreas: abundant influenza viral antigen in nucleus and cytoplasm of acinar cells bordering an area of necrosis. Immunohistochemistry. Hematoxylin counterstain. Bar = 50 μm.

Kidney: influenza viral antigen in nuclei of tubular cells. Some of the fine granular staining in the cytoplasm and the luminal portions of the tubular cells is nonspecific. Immunohistochemistry. Hematoxylin counterstain. Bar = 50 μm.

Ovary: influenza viral antigen in cytoplasm and nucleus of follicular cells. Immunohistochemistry. Hematoxylin counterstain. Bar = 50 μm.

Small intestine: marked staining of the submucosal (Meissner) plexus because of the presence of influenza viral antigen. Note the absence of antigen in the intestinal mucosa. Immunohistochemistry. Hematoxylin counterstain. Bar = 50 μm.
tubular epithelium was noted in one other bird. Viral antigen was found in these 2 ducks in renal tubular cells and in cellular debris in the tubular lumina (Fig. 6).
Spleen. A few spleens (3/14) had mild depletion of lymphocytes in the periarterial lymphatic sheaths and mild diffuse infiltration of heterophils. Influenza antigen was not found in any of the investigated spleens.
Heart. Mild lesions, which ranged from vacuolation of myocardial cells to mild multifocal myocardial fiber necrosis with infiltration of predominantly lymphocytes and macrophages, were observed in half of the cases (7/14). However, viral antigen was not found in any myocardial tissue.
Adrenal glands. Lesions associated with the presence of influenza antigen were found in the adrenal glands of 2 of 10 ducks. One duck had a moderate number of small necrotic foci in the corticomedullary border, with infiltration of a few heterophils, whereas the other duck only had a mild diffuse infiltration of lymphocytes. Antigen was localized to the nuclei of some medullary and cortical cells.
Genital system. Viral antigen was found in the ovarian follicular cells of 1 of 7 female ducks (Fig. 7). There were no lesions associated with the presence of virus. Viral antigen was not detected in the testicles from 3 studied ducks.
Intestines. Neither viral antigen nor histologic lesions were found in the intestinal epithelium except for mild inflammatory reactions associated with parasite infection in 8 ducks. Virus was detected by IHC in the submucosal (Meissner) plexus (Fig. 8) of 3 of 14 ducks and in both the submucosal plexus and the myenteric plexus in 1 duck. No inflammation was associated with the presence of virus in the plexa.
Proventriculus and ventriculus (gizzard). Viral antigen was demonstrated in the nuclei and cytoplasm of cells adjacent to the luminal surface of the proventriculus of 1 of 9 ducks. The cells were not further identified but probably were epithelial cells from the mucosal surface.
Other tissues and organs. Several tissues and organs had minor lesions that were not associated with the presence of viral antigen. These were dominated by mild parasitic lesions in nasal mucosa, lungs, kidney, intestines, adrenal glands, and liver. In the nasal mucosa, chitinized exoskeleton remnants (probably mites) were observed in 1 duck, and lesions similar to those seen around these parasites were observed in 3 more birds. In 2 ducks with inflammatory changes in the lung, microfilariae were found in the lumen of small blood vessels. Eight ducks had parasites in the intestine (trematodes and unidentified parasite remains). A granuloma with trematode eggs was found in the kidney of 1 duck, and trematodes were found in the adrenal glands of 2 ducks.
Discussion
In the current study, tufted ducks naturally infected with HPAI H5N1 revealed a range of mild-to-severe, acute-to-subacute inflammatory lesions that affected between 1 and 6 organs per bird, which confirmed the within-species variation of tissue tropism in H5N1 infections. These variations are likely to be more marked in natural than experimental disease. In naturally infected free-living birds, the clinical and pathologic picture of viral infection is influenced by several factors, such as the age of the bird, the amount and routes of viral exposure, the presence of concomitant infections, the levels of immunity acquired during previous exposure to influenza viruses, 8 and the time course of the infection.
The most consistent lesions in the tufted ducks were multifocal nonsuppurative encephalitis and multifocal necrotizing pancreatitis. This is in agreement with natural H5N1 infection in other wild bird species 6,11,12,14,24,25 and with experimental infection of wild birds, including tufted ducks. 9 The lesions found in the brain and pancreas were similar to those described in other natural and experimental infections. However, in the present study, histologic lesions were not always associated with the presence of viral antigen as might have been expected. In general, the most strongly IHC-positive cells were devoid of light microscopic evidence of cell injury and were located peripherally or adjacent to areas of necrosis. In contrast, necrotic cells were devoid of viral antigen. This may indicate that the preservation of virus in the tissue may be limited to a short period. Viral antigen may thus be difficult to detect in necrotic tissue and in cases with a longer time course of infection. The high number of ducks with encephalitis, in association with high levels of virus as detected by IHC, suggests that the virus is highly neurotropic, as previous studies showed. 4,9,17,25 Signs of neurologic disturbance were, in fact, the main observed clinical signs in infected birds. It is likely that the encephalitis, in combination with an inability to feed or drink, was the ultimate cause of death in most of the birds. Neurotropism of the virus was also observed in the peripheral nervous system, with detection of virus in the submucosal and myenteric plexa of the intestine and in ganglion cells. These findings in the peripheral nervous system are not well documented in wild birds with natural H5N1 infection but have been described in several experimentally infected wild bird species, such as wood ducks, 4 Canada geese, 17 and tufted ducks. 9
Rhinitis, with the presence of viral antigen in nasal epithelium, was another common finding in the tufted ducks. This has rarely been described in naturally infected birds. Rhinitis was not observed in naturally infected mute and whooper swans, 25 whereas experimentally infected mute swans 8 and Pekin ducks 15 had signs of rhinitis. Descriptions in wild birds may be underrepresented, because this tissue is not often investigated in routine postmortem examinations. The presence of mites in the nasal cavity of some of the tufted ducks may account for some of the inflammatory changes in their nasal epithelium. However, the presence of viral antigen in most of the ducks with inflammatory changes suggests that the virus is indeed related to the inflammation. It could be speculated that primary parasitic lesions in the mucosa predispose to nasal HPAI infection.
Despite the presence of congestion and inflammatory changes in the lungs of many of the tufted ducks, mild antigen staining was only found in the lungs of 3 ducks. Viral antigen was not found in the remainder of the respiratory tract (trachea and air sacs). It is probable that, at the time of death, the virus was no longer detectable in the lungs. Some of the inflammatory changes may be attributed to possible concomitant infections. The low number of antigen-positive lungs is in agreement with descriptions in naturally infected mute and whooper swans in which only vascular endothelium in the lung of 2 of 18 swans contained viral antigen, 25 as well as in magpies in which only pulmonary endothelial cells were positive. 11 In general, the lower respiratory tract seems to be less affected in naturally infected wild birds 11,24,25 than in poultry 16 and experimentally infected birds. 9 However, in experimentally infected tufted ducks inoculated intratracheally and intra-esophageally, viral antigen was found in epithelial cells of the bronchus, parabronchus, and atrium, as well as in air capillaries. 9 The route of administration (often intranasal and/or intratracheal) and the viral dose in experimental infections probably increase the likelihood of finding viral antigen in both the upper and lower respiratory tract compared with wild birds in which the route of infection and the virus dose can vary from case to case. Thus, in the naturally infected tufted ducks included in the current study, the virus seems either to be cleared from the lungs fairly rapidly or to be more rhinotropic than pneumotropic.
The heart is often described as one of the main targets for H5N1 viral replication, with the presence of viral antigen in myocytes in association with myocardial necrosis in both naturally 5,11,24,25 and experimentally 9,15 infected birds. Half of the tufted ducks in the current study had lesions in the heart, but viral antigen could not be detected, which indicated that the virus was no longer present or that the changes were not caused by viral infection. Localization of viral antigen to organs, such as liver, kidneys, and adrenal glands, was observed in only a few birds. Although the virus has pantropic potential, other factors seem to affect the distribution of virus and lesions in different organs.
Although the number of organs and the severity of lesions varied among individuals, the disseminated and multifocal distribution of lesions in the tufted ducks suggests that the virus is spread hematogenously. However, there was no evidence of actual vascular damage. Viral antigen was only found in the endothelium and tunica media of a few blood vessels in the nasal mucosa of 2 ducks. This is in agreement with experimentally infected mute swans in which antigen was found in submucosal arteries of the nasal concha in only 3 of 14 swans with signs of rhinitis and which also had large amounts of antigen in other tissues. 8 Experimentally infected Pekin ducks also had signs of rhinitis but had no lesions or antigen in the endothelium, 16 and endotheliotropism was not detected in experimentally infected tufted ducks. 9 The previous studies are in contrast to experimentally infected black swans in which the virus led to a vasculotropic infection, with the presence of viral antigen in the endothelial cells in most visceral organs and the brain. 3
The tufted ducks included in the current study were tested by tracheal and/or cloacal swabs. When considering the very small amounts of antigen visible by IHC in the lower respiratory tract and none visible in the intestinal epithelium, the question arises as to where the virus is derived in the PCR-positive tracheal and cloacal swabs. It was shown experimentally that detection of virus in pharyngeal swabs varied between species–-tufted ducks, common pochards (Aythya ferina), and mallards (Anas platyrhynchos) excreted more virus than common teals (Anas crecca), Eurasian wigeons (Anas penelope), and gadwalls (Anas strepera)–-and among individuals within species. 9 It was further proposed that the virus likely originated from lungs, air sacs, or both. In the naturally infected tufted ducks in the present study, viral antigen was observed by IHC in the lungs of only 3 ducks, even though 16 of 18 tracheal swabs were PCR positive. Possibly the virus was being shed from the nasal cavity. Cloacal excretion of virus was uncommon in the experimentally infected tufted ducks but, when present, was proposed to originate from pancreas, liver, or both. 9 Only 3 of the tufted ducks included in the present study were tested by cloacal swab, and these were positive. It seems likely that the virus originated from the liver or the pancreas in these animals as proposed in experimentally infected tufted ducks. 9 In addition, viral excretion may have originated from the kidneys in 1 bird that had moderate amounts of viral antigen in renal tubular epithelium, something that was not observed in experimentally infected tufted ducks. 9 In a future study, it would be interesting to analyze a larger number of tissue samples with PCR to compare and correlate PCR and IHC results, because this was not within the scope of the present study.
It is interesting to note that in the current study, 2 birds were PCR negative in tracheal swabs but were later confirmed to be infected by HPAI by PCR of cloacal swabs or of brain tissue and thus were included in the study. Similar findings were found in other Swedish wild bird species tested with tracheal swabs. Of 45 HPAI-positive birds (including the tufted ducks in the present study) that were also analyzed histologically, 10 were PCR negative in tracheal swabs. These latter birds had histopathologic changes consistent with HPAI infection, including encephalitis and, in some cases, pancreatitis. They, therefore, were retested by PCR with brain and lung tissue and, subsequently, 5 of these were positive (unpublished data). Thus, PCR of tracheal swabs failed to detect at least 11% of positive cases. The failure to detect positive birds by using tracheal swab analysis only as well as the inter- and intraspecies variations of lesions and distribution of virus imply that sampling for HPAI should include both tracheal and cloacal swabs, and that the highest sensitivity of detection is achieved by testing brain and lung tissue. When considering the fact that nasal epithelium often contained viral antigen, oropharyngeal swabs should perhaps also be included. It should be noted that analysis of swabs alone is a screening method and not a completely reliable diagnostic method.
The acute nature of lesions in naturally infected tufted ducks and the high number of positive tufted ducks compared with other affected bird species observed during the influenza outbreak in Sweden suggest that subclinical infections with a carrier state are probably rarely encountered among tufted ducks. Therefore, tufted ducks found dead or with neurologic signs should be sampled and necropsied because they can serve as “indicator species” for HPAI. Tufted ducks are probably particularly sensitive to HPAI, as suggested in experimental infections, 9 but other factors, such as natural behavior of the ducks, population density, environmental factors, amount of viral exposure, level of viral replication in tissues, and concomitant infections, may also affect the clinical and pathologic picture of avian influenza infections in free-living birds. 15 The Swedish outbreak index cases came from a specific area during special climatic and environmental conditions (cold weather with large amounts of ice cover along the coast and a limited amount of open water) where larger than normal numbers of tufted ducks aggregated in a limited amount of space, which may have created a higher risk of efficient spreading of the virus. To assess how sensitive tufted ducks are to infection with H5N1 in a natural setting, it would have been interesting to sample asymptomatic individuals from areas where positive ducks were found. However, this was not possible during the outbreak. Ancillary diagnostic tests, such as parasitology and bacteriology, would also have been beneficial to assess possible concomitant infections, but these were not conducted because of constraints when working with a biosafety level class-3 pathogen. The description of the pathology in a natural H5N1 infection as described in the current study can serve as a complement to and in comparison with experimental studies. The current study also provides information on variations in presentation of the disease and on confounding factors that should be kept in mind when dealing with natural viral infections and samples obtained from the field.
Acknowledgements
The authors thank Jonas Malmsten and the technicians at the postmortem room for help with necropsies; Ewa Westergren, Lotta Ottander, Susanne Andersson, Gudrun Andersson, and Maria Lögdahl for technical laboratory help; and Thijs Kuiken for help with developing the IHC method. The authors are also grateful to all who helped collect the birds, especially to L. E. Carlsson, who also provided photographs and video films of affected birds. This study was funded by The Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning (FORMAS 221–2007–1177).
Footnotes
a.
MagAttractr Virus Mini M48 Kit, Qiagen GmBH, Hilden, Germany.
b.
Magnatrix 8000+ 96-well extraction robot, NorDiag ASA, Oslo, Norway.
c.
VECTABONDTM, Vector Laboratories Inc., Burlingame, CA.
d.
Proteinase K Dako S3004, Dako Denmark A/S, Glostrup, Denmark.
e.
Bovine Albumin-BSA-Cohn Fraction V-A4503, Sigma-Aldrich Sweden AB, Stockholm, Sweden.
f.
mAbs AI-84, European Veterinary Laboratory, Woerden, The Netherlands.
g.
LSAB Dako 0690, Dako Denmark A/S, Glostrup, Denmark.
h.
AEC-kit Vector SK-4200, Vector Laboratories Inc., Burlin-game, CA.