
Abstract
Bovine tuberculosis caused by Mycobacterium bovis has a wide range of hosts including cattle and humans, but its incidence in otters is very rare. Our report describes a case of bovine tuberculosis in an Asian small-clawed otter (Aonyx cinerea). A deceased female otter ~2–3 years of age that was raised in an aquarium was submitted to the Animal and Plant Quarantine Agency (Anyang, Republic of Korea) for autopsy in June 2013. Following gross pathological examination, many white nodules were observed in the lungs and mesentery. The nodules showed central necrosis infiltrated with lymphocytes and macrophages and surrounded by fibrous tissue. Acid-fast bacteria were detected in the necrotic foci, but no fungi were observed. Molecular analysis led to the detection of M. bovis, which is identified in otters in some European countries such as Spain and France.
Bovine tuberculosis is a chronic disease caused by Mycobacterium bovis and has a wide range of mammalian hosts, including humans.3,6 The clinical signs of bovine tuberculosis are weakness, loss of appetite, weight loss, fluctuating fever, intermittent hacking cough, diarrhea, anorexia, and induration of udder. 19 Though the primary M. bovis host is cattle, domestic and wild animals can be infected, which makes disease control very difficult.3,6 In this report, we describe the clinical manifestations and the gross and microscopic postmortem findings in an Asian small-clawed otter with an infection confirmed by molecular methods as due to M. bovis.
On June 13, 2013, a deceased female Asian small-clawed otter (Aonyx cinerea; Fig. 1A) ~2–3 years old was submitted to the Animal and Plant Quarantine Agency (QIA; Anyang, Republic of Korea) for autopsy and postmortem diagnostics. One month prior to submission, the animal was discovered with a distended abdomen, which worsened 1 week before death. An X-ray examination revealed abnormal structures in the lungs that were suspected to be tumors, as well as ascites and gas in the stomach and intestines. Ultrasound examination also revealed ascites and abnormal structures in the abdominal cavity. Abnormal structures were detected when the abdominal of the otter was palpated, and it was apparent that the animal was in pain.
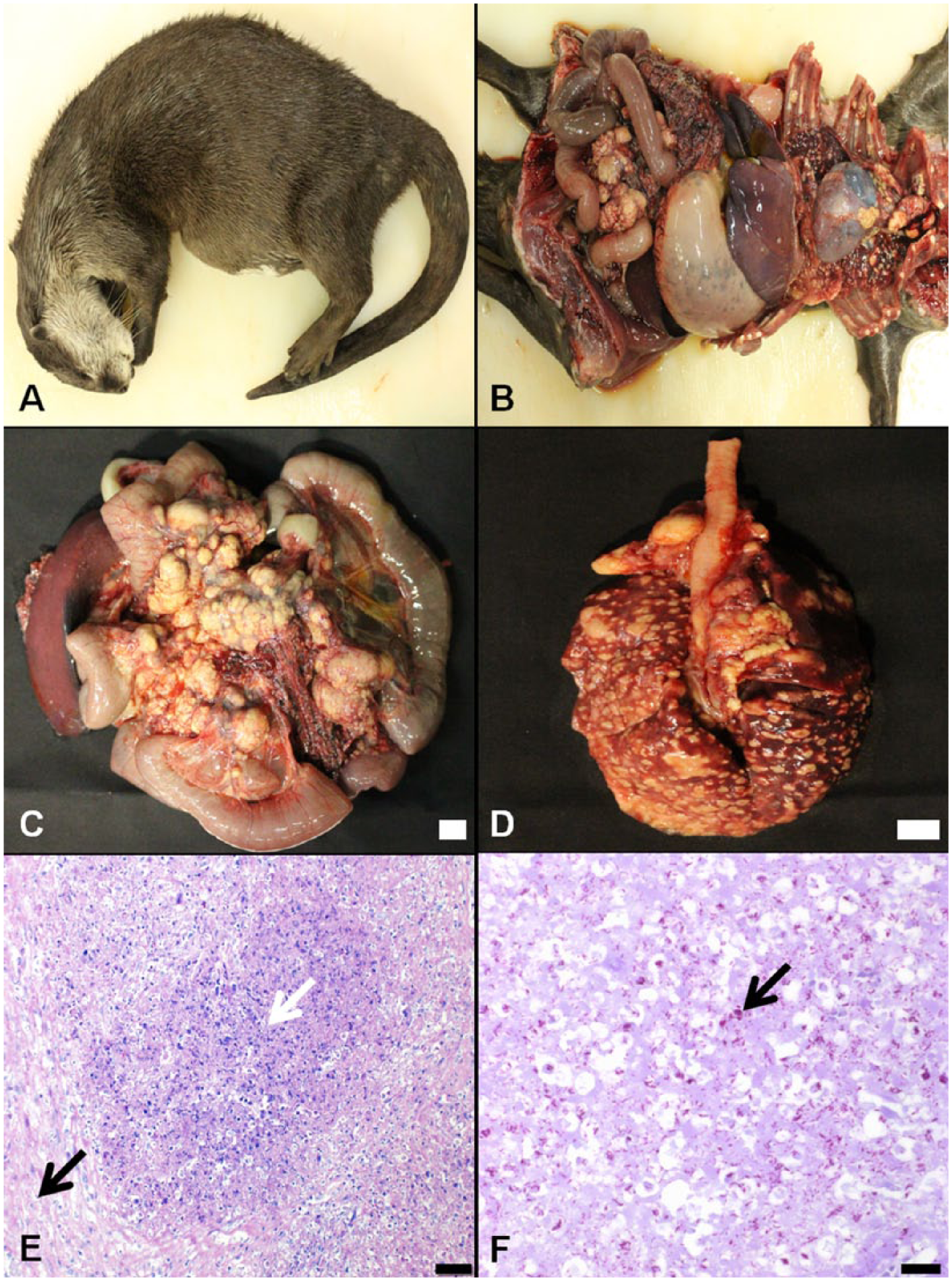
Gross and histological findings.
During an external examination prior to autopsy, solid masses were palpated in the abdominal cavity. When the thorax and abdominal cavity were opened, turbid dark green pleural fluid and ascites were observed. In addition, numerous white nodules of variable size were scattered in the lungs and in the pleura (Fig. 1B, 1D), and the mesenteric lymph nodes were enlarged and likewise contained many white nodules (Fig. 1C). Further examination revealed that the white nodules consisted of lobules on the cut surface, and had a rubbery or hard consistency. Evidence of hemorrhage was apparent in the large omentum, while the small intestine was filled with watery, chocolate-colored contents, and the mucosa was reddish.
After autopsy, the parenchymal organs were fixed in 10% neutral buffered formalin and embedded in paraffin wax. The embedded tissues were sectioned and subsequently stained with hematoxylin and eosin, acid-fast, and periodic acid–Schiff (PAS). Histological evaluations revealed that the mesenteric lymph nodes contained nodules consisting of a central necrotic focus infiltrated with lymphocytes and macrophages, and surrounded by connective tissues (Fig. 1E). In addition, many foamy macrophages were apparent in the sinus of the normal lymph node parenchyma. The nodules observed in the mesenteric lymph nodes were also found in the lungs, but the nodules in the latter tissues exhibited a lesser degree of connective tissue encapsulation. The alveoli in the normal lung parenchyma also contained foamy macrophages and lymphocytes. Ziehl–Neelsen staining revealed acid-fast bacteria in the necrotic foci of the nodules in the mesenteric lymph nodes, and in the lungs (Fig. 1F), but no PAS-positive microorganisms were observed in the tissue sections. In addition, the spleen exhibited extramedullary hematopoiesis and hemosiderosis. Finally, infiltration of lymphocytes and macrophages was apparent in the serosa of the kidney and intestines, as well as the large omentum.
Bacterial cultures of the lung and mesenteric lymph node tissues were performed to identify potential pathogens. Homogenized tissues were preprocessed with 10% oxalic acid for the removal of other microorganisms, and filtered through a 0.7-µm nylon mesh cell strainer. Each sample was cultured on media selective for mycobacteria, specifically two 7H11 agar slopes a with glycerol or pyruvate and a selective broth tube for mycobacterial growth. b The inoculated media were incubated for 8 weeks at 37°C. The species of the mycobacterial isolates were identified by a commercial polymerase chain reaction (PCR) kit c that amplifies the ORFMTCY227.28c region to differentiate between M. bovis and Mycobacterium tuberculosis. Mycobacterium bovis was identified from the lungs and mesenteric lymph nodes (Fig. 2).
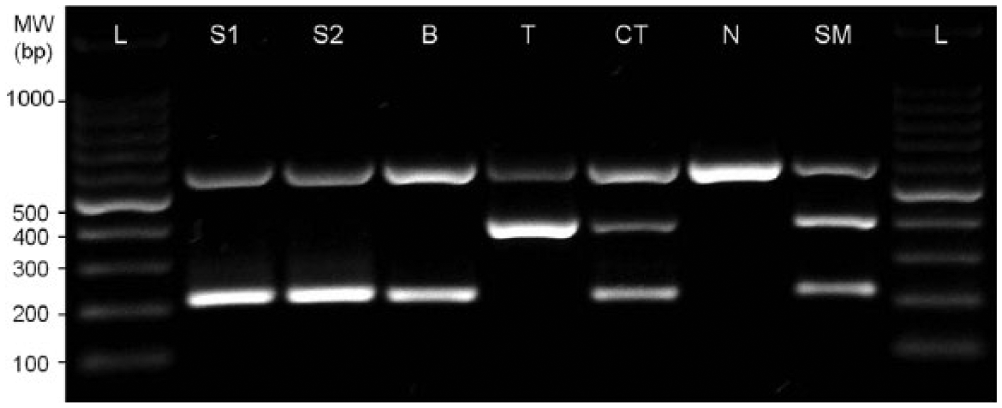
Polymerase chain reaction results for identification of Mycobacterium bovis isolated from lung and mesenteric lymph nodes of Asian small-clawed otter (Aonyx cinerea). The ORFMTCY227.28c region was amplified to differentiate between M. bovis (223 bp, ORFMTCY227.28c flanking region) and Mycobacterium tuberculosis (406 bp, ORFMTCY227.28c region). Lane L: ladder; lane S1: lung isolate; lane S2: mesenteric lymph node isolate; lane B: M. bovis AN5; lane T: M. tuberculosis H37Rv; lane CT: control template; lane N: negative control; lane SM: standard marker.
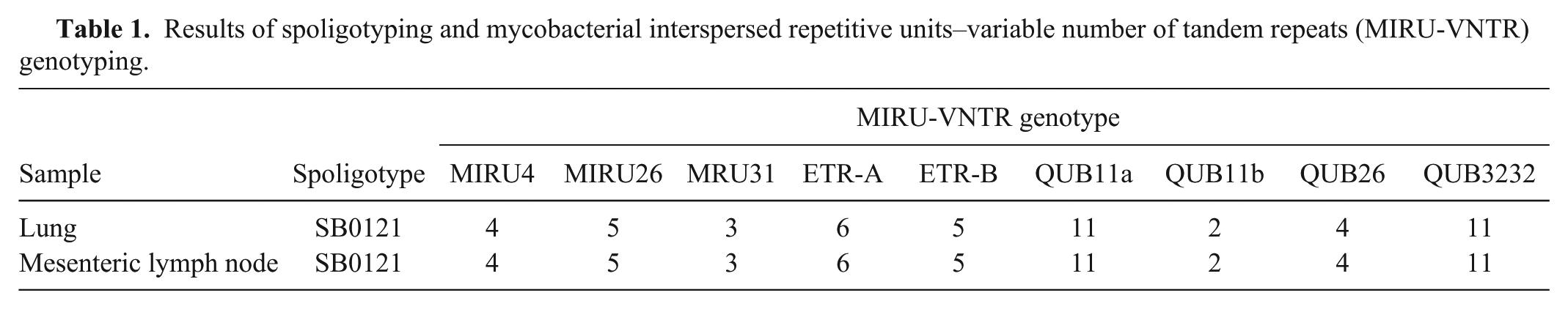
Spoligotyping as well as mycobacterial interspersed repetitive units–variable number of tandem repeats (MIRU-VNTR) typing were used to determine the genotypes of the M. bovis isolates. Spoligotyping was performed using a commercially available kit d as previously described, and the resulting spoligotype pattern was compared to the M. bovis spoligotype database (http://www.mbovis.org/). 8 The MIRU-VNTR typing was performed as previously described using the 9 selected MIRU-VNTR primers (MIRU4, MIRU26, MIRU31, ETR-A, ETR-B, QUB11a, QUB11b, QUB26, and QUB3232).7,11 The results indicated that the 2 isolates from the lungs and mesenteric lymph nodes were spoligotype SB0121, and the MIRU-VNTR genotypes of both were 4-5-3-6-5-11-2-4-11 (Table 1). The SB0121 spoligotype has been mainly identified in Spain, Portugal, France, Belgium, and Italy. 14 Therefore, the MIRU-VNTR genotype of the isolates was compared to the genotypes of other European SB0121 isolates and Korean isolates from livestock using cluster analysis software e as previously described.7,9 Cluster analysis was performed with 7 selective MIRU-VNTR primers (MIRU26, ETR-A, ETR-B, QUB11a, QUB11b, QUB26, and QUB3232) used in previous studies in Spain, France, and Portugal.5,13,17 The results indicated that the genotypes closely clustered with isolates from Spain (Fig. 3).
Results of spoligotyping and mycobacterial interspersed repetitive units–variable number of tandem repeats (MIRU-VNTR) genotyping.
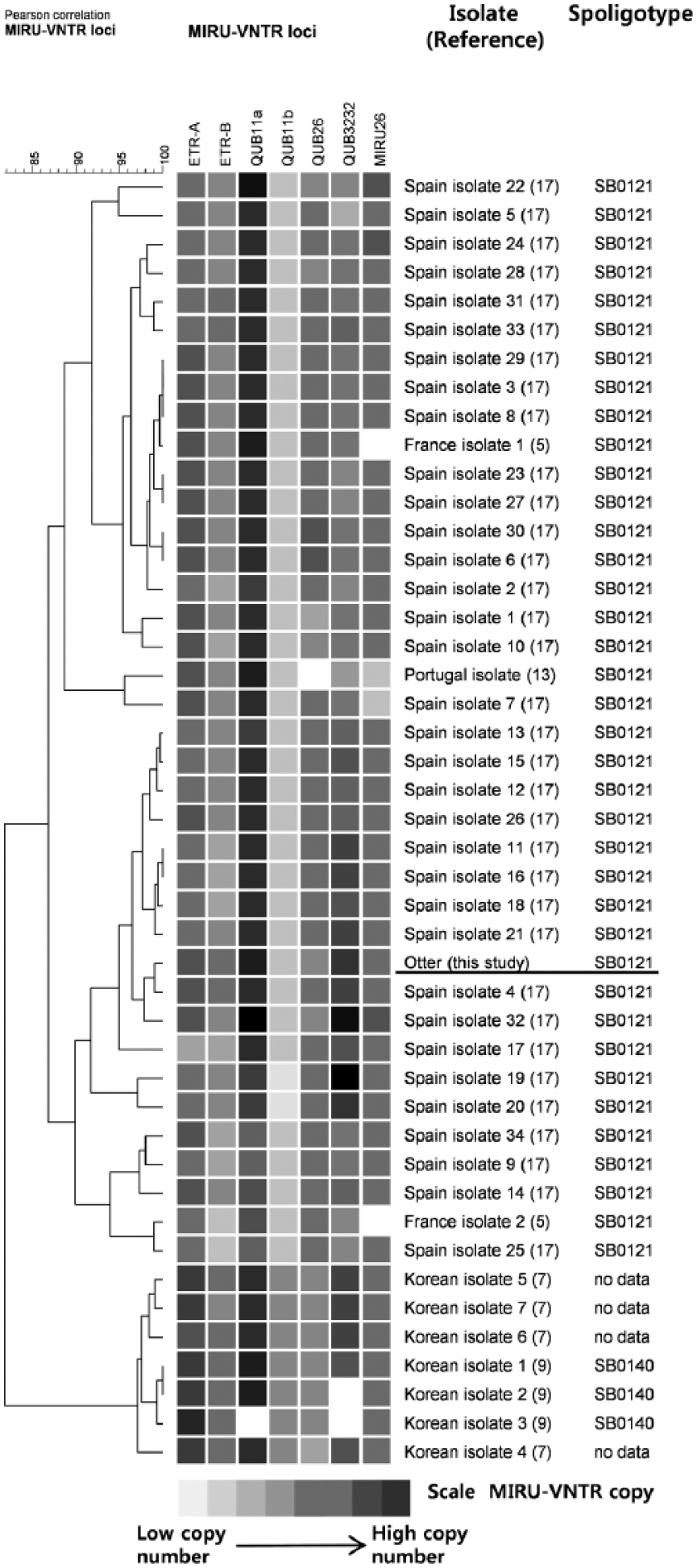
Cluster analysis of Mycobacterium bovis isolates. The color of the scale indicates the range of copy numbers in each locus. The range of copy numbers for 5 primers (MIRU26, ETR-A, ETR-B QUB11b, and QUB26) is 0–10, while the range for 2 primers (QUB11a and QUB3232) is 0–14. The M. bovis isolate in this study is underlined.
The range of hosts susceptible to tuberculosis caused by M. bovis is broad, and includes cattle, deer, elk, bison, buffalo, goats, camels, llamas, swine, elephants, rhinoceros, dogs, foxes, cats, mink, badgers, nonhuman primates, and humans. However, the natural disease is most common in cattle, cervids, humans, and swine. 1
Tuberculosis in the otter is very rare. A review of postmortem examination records of 341 otters from southern and southwest England during the period of 1996–2003 indicated that all of the lung lesions suggestive of tuberculosis examined by histology and culture proved to be negative for mycobacteria. 18 However, a case of bovine tuberculosis was reported in a Eurasian otter (Lutra lutra) in 2009. The carcass of the otter was found in a reservoir in Northern Ireland, and M. bovis was subsequently isolated from the lungs and spleen, and confirmed by acid-fast staining. The spoligotype and VNTR type of the M. bovis isolates from the Eurasian otter were SB0140 and 01, respectively, which suggested that the infection might have been acquired from cattle or a wild animal in Northern Ireland. 10
Seven otters, including the otter submitted to QIA, were imported to Korea from The Netherlands in June 2012. The otters lived together in the aquarium since their importation to Korea, but 3 of them died of similar symptoms in autumn 2012, January 2013, and June 2013, respectively. An autopsy of the first otter that died was not conducted. However, gross lesions apparent in the second dead otter were consistent with the lesions detected in the third otter we examined, but bacterial typing was not performed following autopsy of the second otter. The other 4 otters have been isolated and kept under constant surveillance.
The SB0121 spoligotype of the bacterium isolated in our study was consistent with the pattern identified in some European countries, but this spoligotype has not been previously isolated from an animal in Korea.2,4,16 Furthermore, the results of the MIRU-VNTR genotype cluster analysis indicated differences between the European SB0121 isolates and the Korean isolates (Fig. 3), which strongly suggests that the otter was infected prior to importation. The Netherlands has been officially tuberculosis free since 1993; however, 10 cases of M. bovis infection have been identified in livestock since 1993, all involving imported livestock. 12 Some studies have noted that livestock trading has contributed to the sharing of the SB0121 spoligotype among countries.5,14 It is possible that the otters involved in our study had already been infected with M. bovis prior to export to Korea through contact with imported livestock carriers in the Netherlands.
The route of infection is typically determined by the distribution of the tuberculosis lesions. In most cases, gross lesions are confined to the respiratory tract and are associated with lymphoid tissues. 1 Consequently, the presence of tubercles or craterous ulcers in the intestines or mesenteric lymph nodes suggests an oral route of infection, or ingestion of infected sputum that has been coughed up from the lungs. Furthermore, generalized disease is less common but has been well-established. 1 In the current case, the tubercles were observed in both the lungs and mesenteric lymph nodes. Although the exact mode of transmission was not determined, the autopsy results suggested that both the alimentary and respiratory tracts were potential routes of infection.
Typical histology of tuberculosis lesions found in cattle includes the detection of a central necrotic focus with a variable degree of mineralization, surrounded by a moderate mantle of epithelioid macrophages, lymphocytes, plasma cells, and moderate numbers of Langhans-type giant cells.1,15 Few to numerous acid-fast bacteria were detected within macrophages and giant cells of the mantle zone, as well as extracellularly in the caseous core.1,15 In deer, fibrosis and giant cells are less evident than in cattle, mineralization is more variable, and suppurative inflammation may be prominent.1,15 The histological features of the tubercles found in the otter in this case were different from those previously detected in cattle and deer. Necrosis was prominent, while mineralization, epithelioid macrophages, plasma cells, and Langhans-type cells were not observed. In addition, fibrosis was not prominent, and numerous acid-fast bacteria were present within macrophages and the extracellular space in tubercles and normal parenchyma of the lungs and mesenteric lymph nodes. The lung lesions were less encapsulated accumulations of macrophages and lymphocytes with interspersed areas of fibrinous necrosis that were similar to the lesions observed in the case of bovine tuberculosis in a Eurasian otter. 10
The spoligotype of the bacterium was consistent with the type found in some European countries, but was different from genotypes previously detected in Korea, which suggested that the otter was infected with M. bovis prior to importation to Korea. Consequently, this case indicated the importance of imposing quarantine on imported animals. As well, education and management regarding tuberculosis in aquatic and zoo animals are necessary, because many people, including keepers and visitors, could come into contact with infected animals.
Footnotes
a.
BD Biosciences, Franklin Lakes, NJ.
b.
BD Biosciences, Franklin Lakes, NJ.
c.
SolGent, Daejeon, Republic of Korea.
d.
Optipharm, Osong, Republic of Korea.
e.
Applied Maths NV, Sint-Martens-Latem, Belgium.
Authors’ contributions
H Lee contributed to conception and design of the study. B Lee contributed to acquisition of data. K Baek, H-Y Kim, and S Ryoo contributed to analysis of data. Y Jang contributed to analysis and interpretation of the data. K Lee contributed to interpretation of data. H Lee and Y Jang drafted the manuscript. J-M Kim, M-H Lee, Y-C Bae, E-J Choi, and B So critically revised the manuscript. All authors gave final approval and agree to be accountable for all aspects of the work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the grants (N-AD21-2010-19-05) from National Veterinary Research and Development Foundation, Ministry of Agriculture, Food and Rural Affairs, Republic of Korea.