
Abstract
Toxoplasma gondii can infect almost all warm-blooded animals, and many serological methods have been developed to detect T. gondii infection in a variety of animal species. In the present study, the seroprevalence of T. gondii infection in farmed mink in northeast China was determined using the modified agglutination test (MAT), a Western blot (WB), and 3 enzyme-linked immunosorbent assays (ELISAs) with protein A/G conjugate, using either of 2 recombinant dense granule antigens, GRA1 and GRA7, or Toxoplasma soluble antigens (TSA). There was no significant difference between the detection results of the GRA1-, GRA7-, and TSA-ELISAs and WB (McNemar chi-square, P > 0.05), but a significant difference was observed between MAT and WB (P < 0.05). A near perfect agreement (97.0%) was found between the GRA7-ELISA and WB (κ = 0.83), and a substantial agreement (92.4–93.1%) was observed in the TSA- and GRA1-ELISAs (κ = 0.68–0.73). The GRA7-ELISA showed the highest sensitivity and specificity, and the lowest false-positive and negative rates, while the MAT gave both a low sensitivity and frequent false positives in comparison to the WB. Receiver operating characteristic analysis revealed the largest area under curve of 0.85 (95% confidence interval: 0.74–0.96), and the highest relative sensitivity (72.7%) and specificity (99.0%) for a cutoff value of 0.19 in the GRA7-ELISA. These results indicate that the GRA7-ELISA is suitable for detection of T. gondii infection in mink and that MAT should be used with caution.
Toxoplasma gondii is an obligate intracellular protozoan parasite that infects almost all warm-blooded animals, including humans. Mink, an important fur-producing animal, can be infected with T. gondii by ingesting undercooked slaughter offal or water contaminated with T. gondii oocysts. 5 Several outbreaks of toxoplasmosis in mink have been reported and have caused significant economic losses. For example, ~26% of 7,800 females suffered abortion, or the neonatal kits died, on a mink farm in the United States. 9 Mortality due to toxoplasmosis in wild mink has been reported to reach ~40% in Canada. 19 Mink can be experimentally infected with T. gondii, resulting in the presence of tissue cysts, and thus representing a potential infection source for humans during the pelting process. 4 Feeding of slaughter offal to mink may result in T. gondii infection. 18 In China, mink breeding has being developed rapidly over the past few decades, but the reproductive performances are sometimes very poor. 8 The prevalence of T. gondii infection in mink farms in China is not known.
There are a range of serological methods to detect anti–T. gondii antibodies in humans and animals, such as the Sabin–Feldman dye test (DT), modified agglutination test (MAT), enzyme-linked immunosorbent assay (ELISA), indirect fluorescent antibody test (IFAT), and Western blot (WB).10,11,15 These tests have 1 or more limitations, including the requirement for live parasites, host species–specific conjugate, or variable performance in different species. The ELISA is a useful and convenient diagnostic tool for detecting anti–T. gondii antibodies in serum samples, with the advantages of high sensitivity and specificity.6,13 However, the lack of species-specific anti–immunoglobulin G (IgG) conjugates limits the application of ELISA in some animal species. As protein A/G can strongly bind to the IgG of many mammalian species, the use of a protein A/G conjugate in ELISA has been demonstrated to be useful for the diagnosis of infectious diseases in multiple species. 1
In the present study, we investigated the level of anti–T. gondii IgG in farmed mink in northeast China using the MAT, WB, and 3 ELISAs with protein A/G conjugate and recombinant dense granule antigen 1 (GRA1), GRA7, and Toxoplasma soluble antigens (TSA). The WB was selected as the reference standard for this evaluation study as it has high specificity and sensitivity. 1
The MAT was conducted as described previously. 7 Briefly, each serum sample was diluted in a 2-fold series from 1:25 to 1:200 in 96-well, U-bottom microtiter plates. Serum samples were considered positive for T. gondii antibodies when the MAT titers were ≥1:25. Each test included positive and negative controls.
For the WB, T. gondii lysate proteins were resolved by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane. a The membrane was cut into strips and incubated with the serum sample (diluted 1:50 in Tris buffered saline with Tween 20 [TBS; 10 mM Tris, 150 mM NaCl, pH 8.0, and 0.05% Tween 20]). The membrane strips were further incubated with horse radish peroxidase (HRP)-conjugated protein A/G b diluted at 1:5,000 in TBS after washing. Finally, the strips were incubated with 3,3′-N-diaminobenzidine tetrahydrochloride substrate solution c to visualize the protein bands. A sample was considered positive if there were bands recognized by the test serum. 21
For the ELISA, TSA was prepared by sonicating the T. gondii GT-1 strain tachyzoites. Recombinant proteins GRA1 and GRA7 were expressed in Escherichia coli BL21 (DE3) and purified using a Ni-NTA purification system. d Microtiter plates e were coated with 50 μL of GRA7 (5 μg/mL), GRA1 (5 μg/mL), or TSA (10 μg/mL), respectively. After washing, 100 μL of serum sample diluted at 1:50 in phosphate buffered saline was added to each well, and the plates were incubated for 3 hr at 37°C. Next the plates were washed, and 100 μL of HRP-conjugated protein A/G b diluted at 1:20,000 was added. After incubation for 1 hr at 37°C and washing, color was developed by a substrate solution containing tetramethylbenzidine chromogenic substrate f and stopped by addition of 50 μL of 2 N H2SO4 to each well. The optical densities (ODs) were measured at 450 nm in a microplate reader. g Each serum sample was run in duplicate. The cutoff point of a positive sample was determined as at least 2 times of the negative samples.20,21
Relative sensitivity was defined with the formula [TP/(TP + FN)] × 100, where TP was the number of true-positive samples by the reference method and FN was the number of false-negative samples. Relative specificity was calculated as [TN/(TN + FP)] × 100, where TN was the number of true-negative samples and FP was the number of false-positive samples. 16
A total of 131 blood samples were collected from farmed minks between November and December 2013 in Jilin Province, northeast China. Sera were separated and assayed for anti–T. gondii antibodies by the MAT, WB, and ELISAs based on GRA1, GRA7, and TSA with protein A/G. As shown in Table 1, there were 19 positive and 112 negative samples by the WB, 20 positive and 111 negative samples by the GRA1-ELISA, and 17 positive and 114 negative samples by both GRA7-ELISA and TSA-ELISA, while there were 10 positive and 121 negative samples by the MAT. A higher prevalence of T. gondii infection (15.3%) was obtained with the GRA1-ELISA, followed by the WB (14.5%), GRA7-ELISA (13%), and TSA-ELISA (13%), whereas the lowest prevalence (7.6%) was detected with the MAT (Table 2).
Detection of Toxoplasma gondii antibodies in mink by different methods using the Western blot as the reference method.*
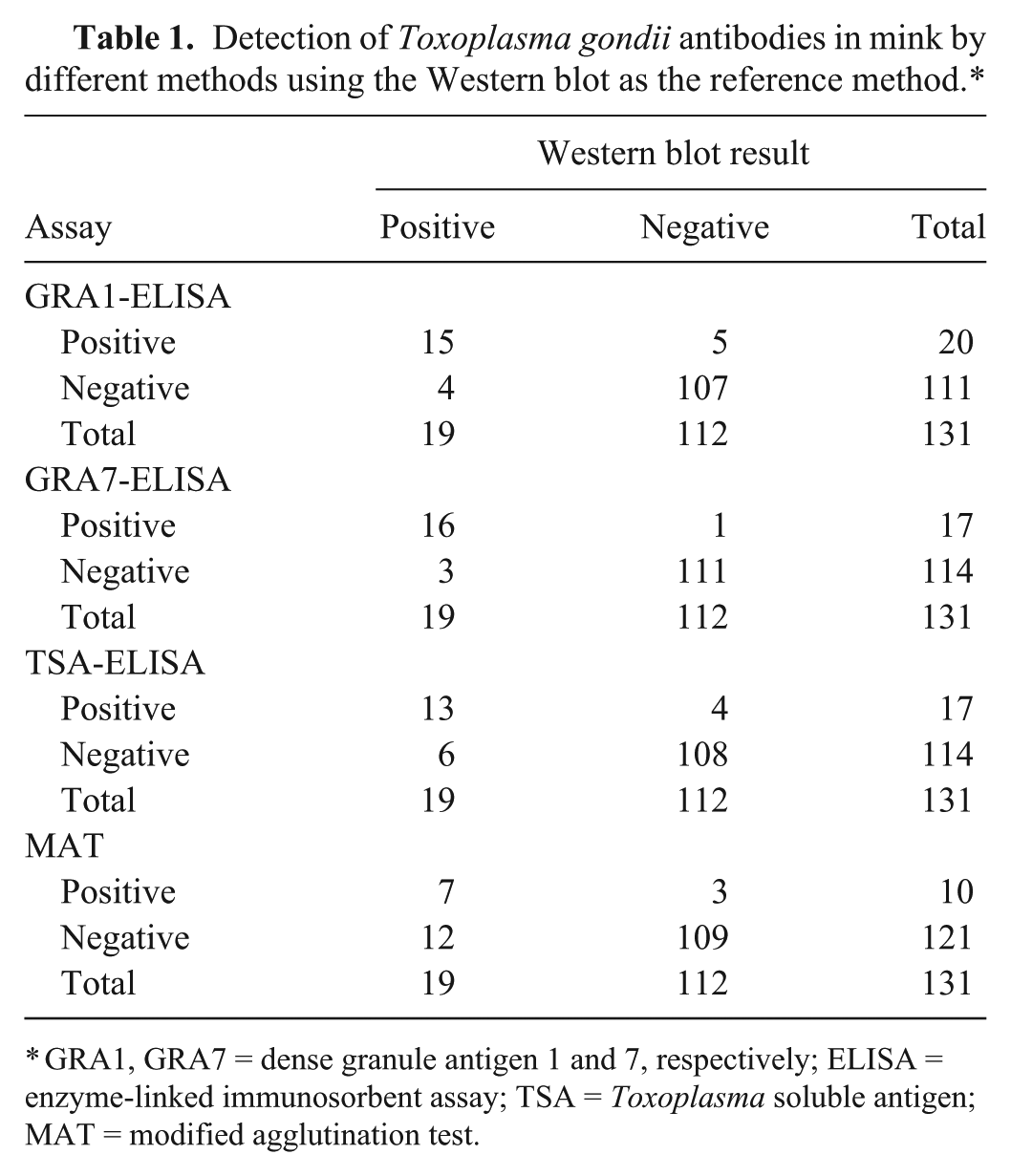
GRA1, GRA7 = dense granule antigen 1 and 7, respectively; ELISA = enzyme-linked immunosorbent assay; TSA = Toxoplasma soluble antigen; MAT = modified agglutination test.
Comparison of detection of Toxoplasma gondii antibodies in mink by modified agglutination test (MAT) and enzyme-linked immunosorbent assays (ELISAs) based on recombinant dense granule antigens (GRA) and Toxoplasma soluble antigen (TSA), with Western blot (WB) as the reference method.
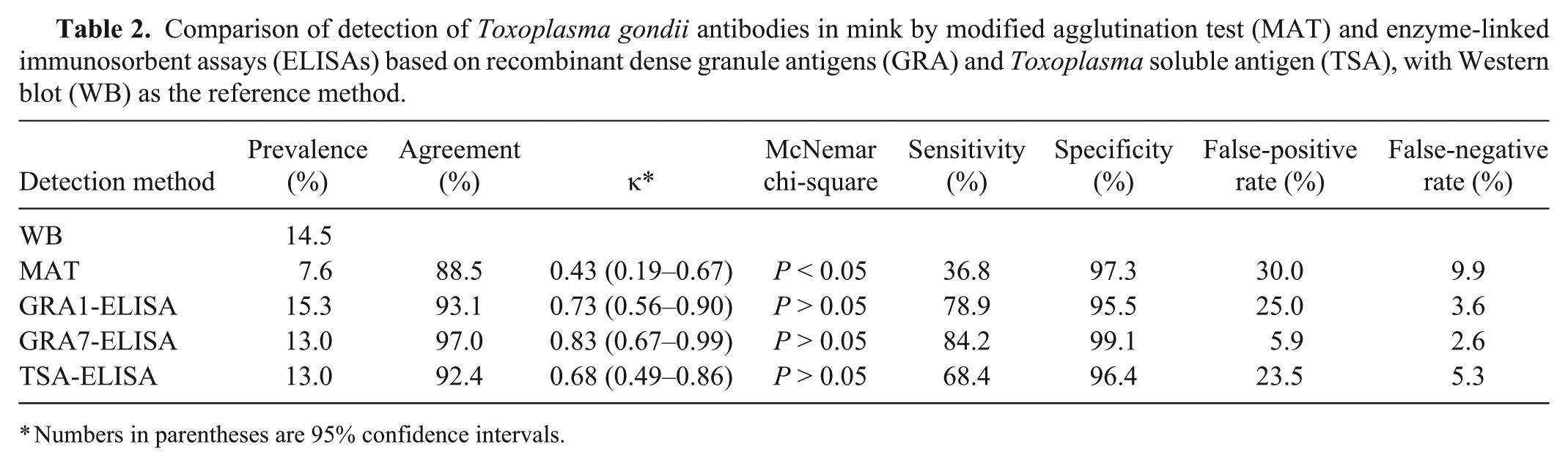
Numbers in parentheses are 95% confidence intervals.
The WB was used as a reference test to validate the sensitivity and specificity of the ELISAs and MAT in the detection of T. gondii–specific antibodies in mink. There was no significant difference in the proportion of positive test results when comparing the GRA1-, GRA7-, and TSA-ELISAs with WB (McNemar chi-square, P > 0.05), but significant difference was observed when comparing the MAT with WB (P < 0.05; Table 2). A near perfect agreement (97.0%) was found between the GRA7-ELISA and WB (κ = 0.83; 95% confidence interval [CI]: 0.67–0.99), and a substantial agreement (92.4–93.1%) was observed in the TSA- and GRA1-ELISA (Table 2).
When compared with the WB, the least false-positive and false-negative samples were found in the GRA7-ELISA (Table 1), demonstrating that the GRA7-ELISA had the highest sensitivity and specificity, and the lowest false-positive and negative rates (Table 2). Receiver operating characteristic (ROC) analysis revealed the largest area under curve (AUC) of 0.85 (95% CI: 0.74–0.96) for the GRA7-ELISA (Fig. 1) and the estimated sensitivities and specificities for different OD ratio cutoff values, showing that the cutoff value at 0.19 for the GRA7-ELISA has the highest relative sensitivity of 72.7% and relative specificity of 99.1% among the 3 ELISAs (Fig. 2).
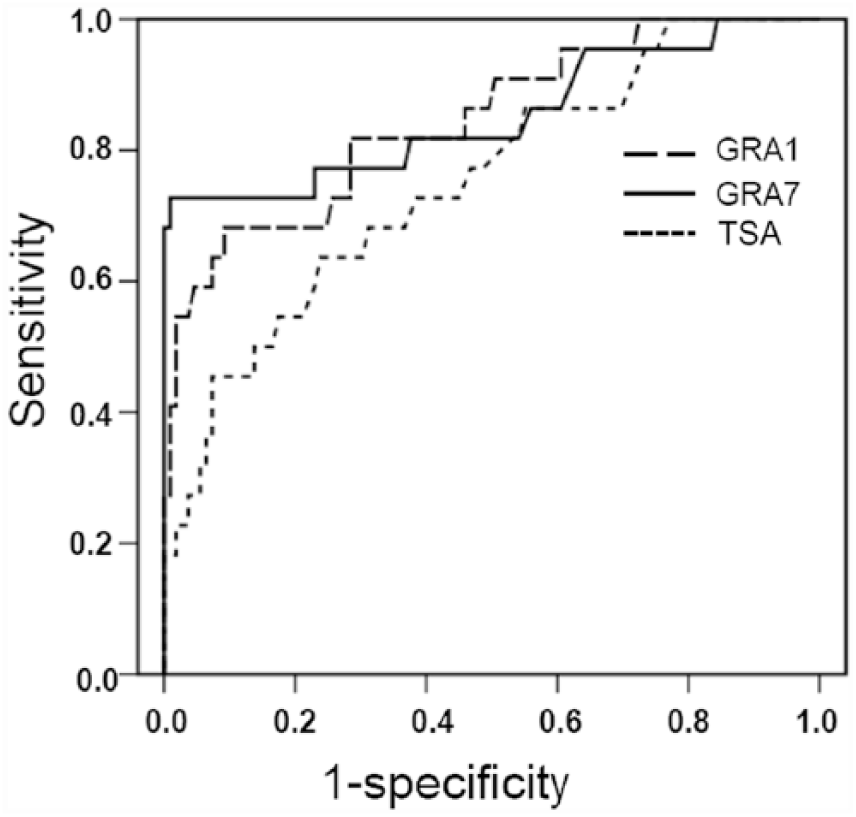
Receiver operating characteristic (ROC) analysis of the enzyme-linked immunosorbent assays (ELISAs). The ROC analysis showed an area under the curve of 0.84 (95% confidence interval (CI): 0.75–0.94) for dense granular antigen 1 (GRA1)-ELISA, 0.85 (95% CI: 0.74–0.96) for the GRA7-ELISA, and 0.75 (95% CI: 0.64–0.87) for the Toxoplasma soluble antigen (TSA)-ELISA.
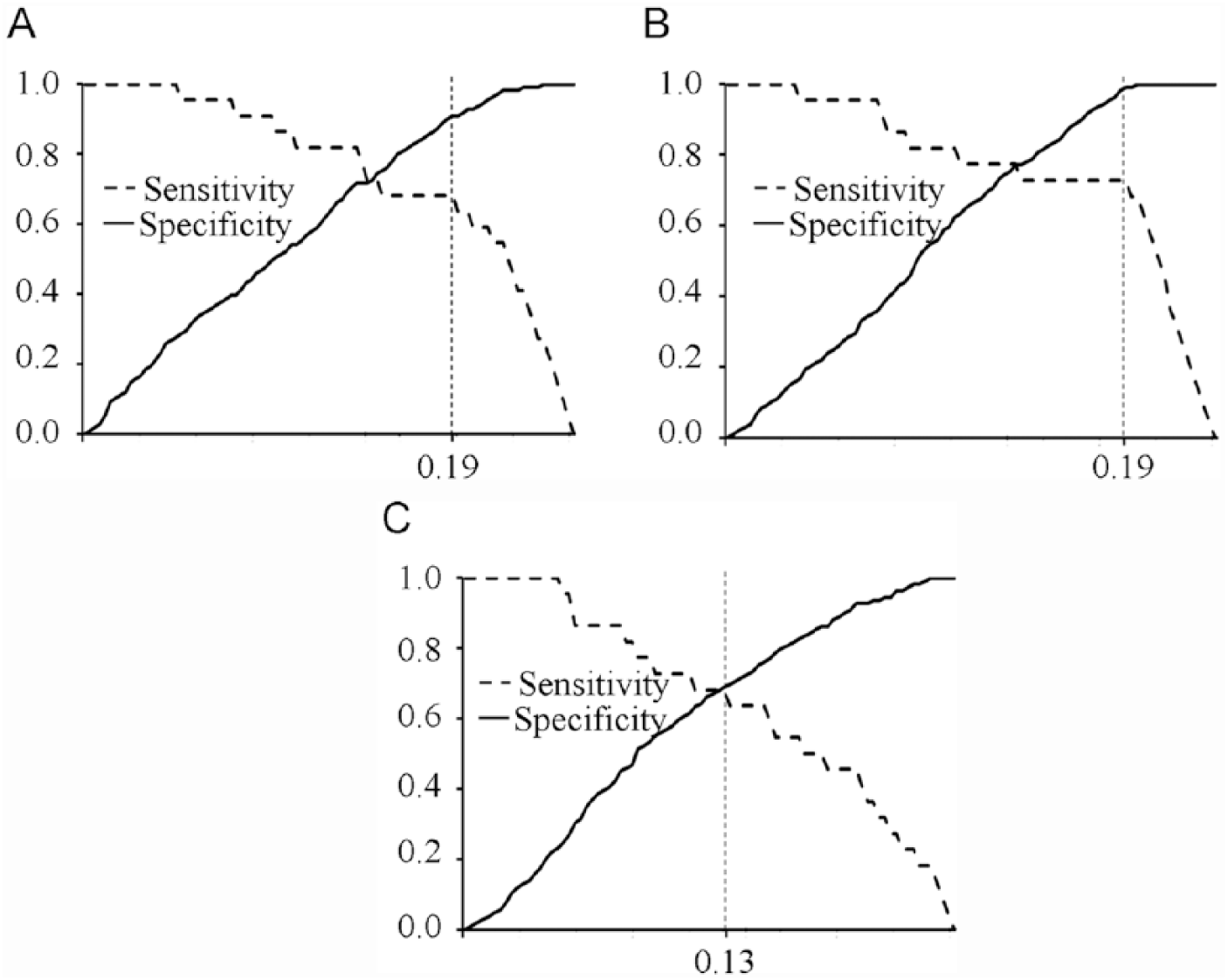
Relative sensitivity and specificity of enzyme-linked immunosorbent assays (ELISAs) based on dense granular antigen 1 (GRA1), GRA7, and Toxoplasma soluble antigen (TSA) at different cutoffs.
Serological techniques, such as IHA, MAT, IFAT, and ELISA, are widely used to detect T. gondii infection in animals and humans.6,17 Among these tests, IFAT and ELISA require host species–specific conjugate. Though IHA and MAT are not species specific, their sensitivity and specificity may vary in different species. For example, IHA is an insensitive test in chickens, 3 and MAT may produce frequent false-negative results in dogs. 22 No commercial species-specific conjugate is available for some host species, making it difficult to evaluate the performance of MAT and IHA in these animals. Previous studies have shown that protein A/G conjugate can be used for detection of anti–T. gondii IgG in some mammalian hosts, 1 a situation confirmed in the current study for mink.
The most commonly used method for the detection of T. gondii infection worldwide is MAT as it is not species specific.2,6,20 However, the MAT shows frequent false negatives in dogs and in mink. 14 Thus, MAT should be used with caution in these 2 species.
In the present study, we used the ELISA and WB with HRP-conjugated protein A/G to detect anti–T. gondii IgG in mink. In the WB, there were specific bands recognized by the positive serum samples, and no band was observed in negative samples (data not shown). With the WB as the reference test, the GRA7-ELISA showed the highest sensitivity and specificity, and the lowest false-positive and false-negative rates among the evaluated tests. The agreement between the GRA7-ELISA and WB was excellent, with the κ value of 0.83 (Table 2). Among the 3 ELISAs, the largest AUC and the highest relative sensitivity and specificity from ROC analysis was observed with the GRA7-ELISA, suggesting that the GRA7-ELISA is a suitable method for serological detection of T. gondii infection in mink.
Our study showed different prevalences of T. gondii infection in mink by the MAT, WB, and ELISAs, due to the differences in the sensitivity and specificity of these tests. The detection results of the GRA7-ELISA (13%) and WB (14.5%) might reflect the real infection status of T. gondii in farmed mink in northeast China. In Poland, a similar overall seroprevalence, as detected by the latex agglutination test (LAT), of T. gondii infection (13.9%) was found in farmed mink, although a higher seropositivity (26.3%) was present in large farms, and a lower seropositivity (2.9%) in small farms. 18 In contrast, in Denmark, a low prevalence (3%) was found in farmed mink by LAT. 12 These differences between the studies may be related to differences in the serological tests, the mink populations, the local climatic factors or to some combination of these conditions. Overall, our study confirms that the prevention and control of toxoplasmosis in farmed mink in northeast China needs to improve.
Footnotes
Authors’ contributions
Q Liu contributed to conception and design of the study. Y Gu, Z Wang, Y Cai, X Li, F Wei, L Shang, and J Li contributed to acquisition, analysis, and interpretation of data. Y Gu drafted the manuscript. Q Liu critically revised the manuscript and gave final approval. All authors agree to be accountable for all aspects of the work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Chinese National Nature Science Foundation and the Special Fund for Agro-scientific Research in the Public Interest in China.
a.
Polyvinylidene fluoride (PVDF) membrane, Millipore Corp., Billerica, MA.
b.
Horseradish peroxidase (HRP) conjugated protein-A/G, Thermo Fisher Scientific Inc., Waltham, MA.
c.
3,3′-N-diaminobenzidine tetrahydrochloride substrate (BAD) kit, CWbio, Beijing, China.
d.
Ni-NTA purification system, Qiagen GmbH, Dusseldorf, Germany.
e.
Microplate, Jet Biofil, Guangzhou, Guangdong Province, China.
f.
Tetramethylbenzidine chromogenic substrate TMB, Thermo Fisher Scientific Inc., Waltham, MA.
g.
Microplate reader ELX-800, BioTek Instruments Inc., Winooski, VT.