
Abstract
In a consignment of sheep brains from New Zealand, to be used in Europe as negative control material in scrapie rapid screening test evaluations, brain samples from 1 sheep (no. 1512) gave the following initially confusing results in various screening tests: the brainstem repeatedly produced negative results in 2 very similar screening kits (enzyme-linked immunosorbent assay [ELISA]-1, ELISA-2), a macerate made from brainstem and cerebellum returned a clearly positive result in ELISA-2, and the macerate and a brainstem sample gave negative results in a third screening test (ELISA-3). In subsequent testing, cerebellum tissue alone tested strongly positive in ELISA-1 and produced a banding pattern very similar to atypical scrapie/Nor98 in a confirmatory Western blot (WB). The macerate showed weak staining in the confirmatory WB but presented a staining pattern identical to atypical scrapie/Nor98 in the scrapie-associated fibril WB. The latter test confirmed conclusively the first case of atypical scrapie/Nor98 in a sheep from New Zealand. Other parts of the brain either tested negative or very weak positive in ELISA-2 and in WBs, or tested with negative results by histopathology and immunohistochemistry. It appears that sheep no. 1512 is a case of atypical scrapie/Nor98 in which the abnormal prion protein was detected mainly in the cerebellum. This case emphasizes the need to retain brainstem, and cerebral and cerebellar tissues, as frozen and fixed materials, for conclusive confirmatory testing. Furthermore, consideration should be given to which screening method to use.
Introduction
Ruminants are susceptible to a group of fatal neurologic diseases known as transmissible spongiform encephalopathies (TSE), or prion diseases, which include bovine spongiform encephalopathy in cattle (BSE), scrapie in sheep and goats, and chronic wasting disease in deer and elk. 4,43 An abnormal form (PrPSc) of the normal cellular prion protein (PrPC) accumulates during the disease process in nervous tissue and appears to be the major component of the disease agent of TSEs. 1 New Zealand livestock are free from BSE, scrapie, chronic wasting disease, and other animal TSEs 6,40 (Bruere AN: 2003, Scrapie freedom—the New Zealand story. Surveillance 30:3–7), and until October 2009, had been considered free from atypical scrapie/Nor98 (McIntyre L, Kittelberger R: 2009, A case of atypical scrapie/Nor98 in a sheep from New Zealand. Surveillance 36:6–10).
Scrapie, herein referred to as classical scrapie (to distinguish it from atypical scrapie/Nor98), has been known for hundreds of years. 40 Scrapie is not considered to pose a risk to human health (World Organisation for Animal Health: 2009, Scrapie. Terrestrial Animal Health Code, Chapter 14.9. Available at http://www.oie.int/eng/normes/mcode/en_chapitre_1.14.9.htm. Accessed on July 16, 2010). Multiple strains of the scrapie agent have been identified by differences in distribution in host tissues, variation in incubation periods, lesion profiles, neuropathology, protease resistance, and other features. 35 In 1998, a new prion disease, initially named Nor98, was discovered in sheep in Norway. 4,5 From 2002 onward, scrapie surveillance was intensified in Europe because of concerns that BSE may have entered the small ruminant population. An abnormal scrapie prion protein, different from BSE and different from the classical scrapie prion, was detected during testing with rapid screening tests in a consistent proportion of the samples tested. 4,11,15 The related prion disorder was named “atypical” or “discordant” or “nonclassical” scrapie and was found to be identical to Nor98. 4 In contrast to classical scrapie, in which many distinguishable strains exist, no substrains of atypical scrapie/Nor98 have been identified to date. 4,39
The majority of atypical scrapie/Nor98 cases have been identified in clinically normal sheep sampled at slaughter. 4,12–14,21 Cases have been reported from Norway, Sweden, Finland, the United Kingdom, Germany, France, Portugal, Belgium, The Netherlands, Ireland, Denmark, 15 Spain, 34 the United States, 25 and Canada. 28 Retrospective testing of archived tissues has shown that atypical scrapie/Nor98 has existed in the United Kingdom at least since 1987. 44 The apparent prevalence of the cases in the European Union member states is very low (e.g., in the United Kingdom, prevalence is less than 0.1%) and is distributed evenly across countries. Active surveillance programs have led to the detection of atypical scrapie/Nor98 cases, even in the absence of any case of classical scrapie in sheep in Portugal, Denmark, Sweden, Finland, Poland, 15,33 and in the Falkland Islands, 9 the first region in the Southern Hemisphere to report a case of atypical scrapie/Nor98. A case of atypical scrapie/Nor98 has recently been confirmed in western Australia (report in preparation for Australian Animal Health Surveillance 2 Quarterly, October 2010).
Atypical scrapie/Nor98 appears to be a noncontagious, sporadic degenerative condition of older sheep. A spontaneous etiology, possibly with a genetic determinant, environmental influences, and/or metabolic factors have been suggested in a number of studies based on epidemiologic evidence. 12–15,17,21,26 Contributing to this suggestion is the observation that atypical scrapie/Nor98 cases have been detected by active surveillance programs, wherever such programs have been initiated. 15 In most cases, there has been only a single atypical scrapie/Nor98 case in a flock, and this led to early speculation that the condition is not naturally infectious. 4,12–14,21 Experimental oral transmission to sheep has been unsuccessful to date. Experimental oral transmission to standard mice and to bank voles (Myodes glareolus) also failed. Only experimental intracerebral transmission to transgenic mice expressing the ovine 3,23 or porcine 10 prion gene, and experimental intracerebral challenge in sheep, have so far been successful. 38
The distribution of pathologic changes and deposits of the disease-associated isoform of the prion protein in atypical scrapie/Nor98 are different from classical scrapie. 4 Generally, the intensity of staining by immunohistochemistry (IHC) of PrPSc in the medulla oblongata is much less in atypical scrapie/Nor98 than in classical scrapie cases. The cerebellar and cerebral cortices are the most immunostained areas of the atypical scrapie/Nor98 brains. 4,29,32 Nevertheless, in some cases, immunolabeling can be minimal or totally absent from the cerebellum, whereas, in other cases, the only brain area that shows immunolabeling is the cerebellum. 4,32 In contrast to classical scrapie, PrPSc has not been detected in peripheral tissues of atypical scrapie/Nor98 cases. 4,15
Since 1999, the European Commission has evaluated various rapid postmortem tests for BSE. 7,8,27 For a number of years, these BSE test methods had also been accepted for scrapie testing, with assuming similar performance for scrapie as for BSE. After evaluation of rapid tests for scrapie (European Food Safety Authority [EFSA]: 2005, Scientific report of the European Food Safety Authority on the evaluation of rapid post mortem TSE tests intended for small ruminants. EFSA Sci Rep 31:1–17, and 49:1–16), the EFSA recommended the use of 8 rapid tests for the detection of scrapie on brainstem samples. For the detection of atypical scrapie/Nor98, 7 of these tests could be used but only on cerebellum or cerebrum, rather than brainstem samples. Three of these rapid tests, enzyme-linked immunosorbent assay (ELISA) test kits (ELISA-1, a ELISA-2, b and ELISA-3 c ), were also recommended for the detection of atypical scrapie/Nor98 in the brainstem.
Ever since rapid postmortem test evaluations have been performed by the European Union, the New Zealand Ministry of Agriculture and Forestry, Biosecurity New Zealand (MAFBNZ) and its subcontractor, AsureQuality New Zealand (Whangarei, New Zealand), have supplied ruminant brain tissues as negative control materials for the European Union evaluations mentioned above (McIntyre L: 2007, New Zealand's contribution to explaining the pathogenesis of atypical scrapie. Surveillance 34:9–10), as shown in Table 1. Brain tissues from 4,903 animals, including cattle, sheep, and goats, had been supplied to the European Union by mid-2009.
In 2008, a consignment of 800 sheep hindbrains, 150 goat hindbrains, and 200 sheep whole brains was sent to the Institute for Reference Materials and Measurements (IRMM; Geel, Belgium) of the European Commission, the institute that organizes the TSE rapid test evaluations in Europe as set out in the EFSA protocols (EFSA: 2007, Protocol for the evaluation of rapid postmortem tests to detect TSE in small ruminants. Scientific Opinion of the Panel on Biological Hazards. EFSA Journal 509:1–31). In accordance with recommendations for the detection of classical scrapie and atypical scrapie/Nor98 (EFSA: 2005, EFSA Sci Rep 31:1–17), a brainstem sample posterior to the obex from every animal had been tested at the MAFBNZ Investigation and Diagnostic Centre (IDC) Animal Health Laboratory (AHL) by using ELISA-1 before shipment. All samples returned negative results. After arrival at IRMM, brainstem slices had been tested at a contract laboratory by using ELISA-3, with negative results. In July 2009, 1 of 200 macerates, each containing a mixture of brainstem and cerebellum, returned a positive result by using ELISA-2 at the Veterinary Laboratories Agency (VLA UK; Surrey, United Kingdom), a World Organization for Animal Health (OIE) reference laboratory for BSE and scrapie. Macerates had been prepared and tested by VLA UK on behalf of IRMM. Subsequent testing of the macerate by VLA UK by using ELISA-3 gave negative results. These results were reported to the IDC in September 2009, which led to a veterinary laboratory diagnostic investigation.
Summary of shipments of brain tissues supplied by the New Zealand Ministry of Agriculture and Forestry, Biosecurity New Zealand, to the European Union.

Comprehensive testing performed at IDC AHL and at VLA UK revealed the first case of atypical scrapie/Nor98 in a sheep in New Zealand. The present report describes, in detail, the laboratory investigation and confirmation of this atypical scrapie/Nor98 case.
Materials and methods
Brain tissue collection in New Zealand
In November 2007, brain tissues from sheep, including whole brains from 200 sheep, had been collected in New Zealand at slaughter plants from healthy sheep more than 3 years old. Dentition of heads had been checked to ensure that the brains were collected from animals with 6 or more permanent erupted incisors. For whole-brain collection, the skull was cut transversely at the level of the ramus of the mandible, and the whole brain was removed with sterile gloves and placed on a sterile plastic sheet for dissection. The saw cut was sliced clean on both sides of the cerebrum with a sterile scalpel and then discarded.
From the dorsal aspect, the brainstem was cut transversely anterior and posterior to the obex to a width of 2–3 mm. A 3-mm section posterior to the obex and a 5-g slice of cerebellum (only 1.3 g in the case of sheep no. 1512) were frozen. A 3-mm section anterior to the obex was placed in 10% buffered formalin. These tissues were kept at IDC AHL for reference purposes. The fresh tissues were frozen at −25°C; the fixed tissue was kept at ambient temperature.
The remainder of the whole brain was carefully placed in 2 adjacent pieces (the anterior cerebrum abutting the posterior remainder of the cerebrum, cerebellum, and brainstem) in prenumbered plastic bags, heat sealed after expelling the air, and frozen at −25°C. In May 2008, the brains were shipped on dry ice to IRMM.
Tissue processing in Europe
In July 2009, brainstem tissues from 200 New Zealand sheep brains were processed at the VLA UK, under contract from IRMM, into macerates (i.e., homogenates of equal amounts of tissue and water). In the case of sheep no. 1512, the entire cerebellum and most of the brainstem were used for obtaining a total of 20 g of tissue.
The macerate was made by first removing any blood clots or connective tissue, dicing the cleaned material into small cubes (approximately 1 cm3), and adding an equal amount of sterile distilled water (w/v). This material was then homogenized for 3 periods of 30 sec with a domestic handheld blender d at low speed. If there were any visible lumps after this procedure, then a further period of 30 sec of homogenization was required. The homogenate was then placed on a vortex mixer for approximately 1 min before being divided into 1-g aliquots.
In addition, in 2009, slices of brainstem of the New Zealand sheep, including sheep no. 1512, were sent by IRMM to the National Veterinary Institute (Copenhagen, Denmark) to be tested by ELISA-3. In July 2009, IRMM requested a second test by using ELISA-3 to reconfirm the first negative result on the slice of brainstem from the sheep no. 1512 sample.
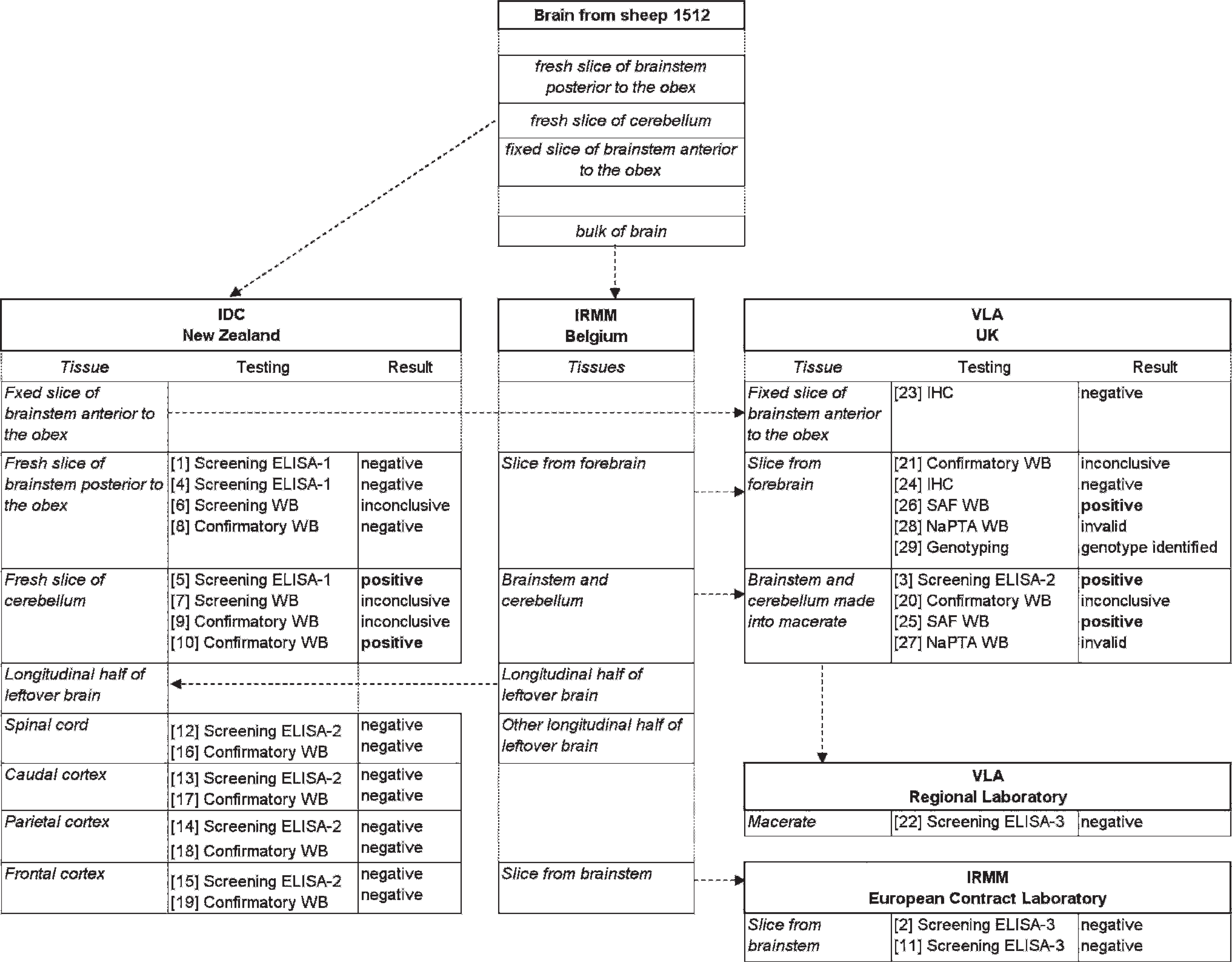
Flow diagram, showing use, movement, testing, and test results of brain tissues from sheep no. 1512 at various institutions. Note: Tissue movements and testing are not in chronologic order. Numbers in square brackets give the chronologic order of testing.
ELISA-1 and ELISA-2 testing
ELISA testing for both ELISA-1 and ELISA-2 (which are from the same manufacturer and are very similar in performance and regarding evaluation data) was either performed by using the ruminant ELISA-2 kit (at VLA UK) or the sheep and goats ELISA-1 kit (at IDC) by following the manufacturer's instructions. Both kits had been shown to perform equally well for scrapie prion protein detection on brain tissues (EFSA: 2005, EFSA Sci Rep 31:1–17, and 49:1–16). Although only 3 atypical scrapie/Nor98 samples were available for the evaluation, both kits had been recommended for the detection of atypical scrapie/Nor98 in cerebrum, cerebellum, and brainstem tissues.
Homogenates of fresh brain samples or macerates were prepared from 0.350 g tissue in a ryboliser e by using the grinding tubes provided in the test kit. From the grinding tubes, 250 μl was removed by using a syringe, mixed with proteinase K, and digested at 37°C for 10 min. After addition of 250 μl of precipitation buffer, the sample preparation was centrifuged f at 15,000 × g at 20°C for 7 min. The supernatant was removed, and the pellet was redissolved in 25 μl of sample buffer by heating at 100°C for 5 min and vortexing afterward.
For the ELISA testing, 100 μl of sample buffer was added to every sample and mixed, and 100 μl were loaded onto a microtiter plate that was coated with a prion protein—specific monoclonal antibody (mAb). The plate was incubated at 37°C for 30 min, washed 3 times with washing buffer, and then incubated with 100 μl of conjugate at 2°C–8°C for 30 min. After 5 washes, 100 μl of revelation solution (substrate) was added, and the plate was incubated in the dark at room temperature (18–30°C) for 30 min. After adding 100 μl of stop solution, the absorbance or optical density (OD) was measured at 450 nm by using a spectrophotometer.
All samples with an OD value below the OD mean of 4 negative control samples plus a factor of 0.210 (ELISA-2) or a factor of 0.140 (ELISA-1) were considered negative. Samples equal to or greater than this cutoff value were considered positive.
ELISA-3 testing
ELISA-3 was performed according to the manufacturer's instructions. Briefly, each sample was diluted with a working plate diluent and then loaded into the abnormal prion protein—specific antigen-capture plate, along with positive and negative controls, followed by 2 hr of incubation at room temperature. After washing the plate and incubating with the conditioning buffer for 10 min at room temperature, the plates were washed and incubated with a horseradish peroxidase—conjugated anti—prion protein antibody for 1 hr at room temperature. After washing and incubation with substrate for 15 min, the reaction was stopped, and the absorbance was measured at 450 nm by using a spectrophotometer. All samples with an OD value less than the mean of 2 negative control samples plus a factor of 0.180 were considered negative. Samples equal to or more than the mean of 2 negative controls plus 0.180 were considered positive.
Confirmatory WB testing
The confirmatory WB g assay was performed by following the manufacturer's instructions. Briefly, homogenates were prepared either by using tissues or macerates as for the ELISA-1 or ELISA-2, but pellets were redissolved in Laemmli sample buffer that contained 5% (v/v) mercaptoethanol and 2% (w/v) sodium dodecyl sulfate. After centrifugation, the supernatants were heated at 100°C for 5 min and centrifuged before electrophoresis of the supernatant. Protein separation was performed by gel electrophoresis in 18-well 12% Criterion XT Bis—Tris gels under reducing conditions. h The proteins were wet transferred onto polyvinylidene fluoride (PVDF) membranes at 115 V for 60 min and then blocked for 30 min at room temperature. The kit's primary antibody (mAb SHA31, diluted to 1:10 of the supplied kit concentration) was applied for 30 min at room temperature. Membranes were washed, incubated for 20 min in the kit's secondary antibody at room temperature, washed again, and incubated with an enhanced chemiluminescent substrate. Digital images of the blots were produced by using an image analysis system. i At VLA UK, in addition to the kit mAb SHA31, a PVDF membrane with the same samples as for the kit mAb was probed with the mAb P4 j at a dilution of 1:5,000, an antibody known to react with the N-terminal end of the prion protein.
OIE scrapie—associated fibril WB
The OIE scrapie—associated fibril (SAF) WB method was performed according to standard procedures at the VLA UK, as described in detail elsewhere. 42 Starting material was 2 g of forebrain tissue cut into small pieces and 4 g of macerate from sheep no. 1512. To each sample, brain lysis buffer (10% N-lauroylsarcosine sodium salt in 20 mM sodium phosphate buffer pH 7.4) was added and thoroughly homogenized in a 50-ml conical plastic tube by using a homogenizer. k After transfer to a centrifuge tube, the homogenate was centrifuged at 20,000 × g for 30 min at 10°C. The supernatant was carefully moved to clean centrifuge tubes, filled to two-thirds of the tube with brain lysis buffer, and centrifuged at 177,000 × g for 2 hr 15 min at 10°C. After discarding the supernatant, suspending the pellet in 16.7 mM Tris—HCl buffer (pH 7.4), and incubation in a water bath at 37°C for 15 min under constant stirring, a 15% buffered potassium iodide solution was added, and the mixture was incubated for a further 30 min. The macerate homogenate was divided into 2 aliquots; 1 aliquot was digested with proteinase K, and the other remained undigested. The entire forebrain homogenate was treated with proteinase K. After centrifugation at 189,000 × g for 1 hr at 10°C, the supernatants were removed, and the pellets were redissolved in 1 M sodium chloride solution. This solution was centrifuged at 11,600 × g for 15 min at room temperature, the supernatant was removed, and 50 μl of formic acid was added to the pellet. The pellet was dried in a concentrator l for 1 hr at 45°C. Samples and controls then were dissolved in 40 μl of electrophoresis sample buffer by heating at 100°C for 5 min.
Gel electrophoresis of samples and controls was done in 12% gels m at 200 V for 45 min in a gel electrophoresis system. n Electrophoretic transfer to PVDF membranes was performed at 150 V for 60 min (the same procedure as for the confirmatory WB g ). Afterward, the membranes were blocked at room temperature for 30 min, incubated with primary antibody (mAbs SHA31, 6H4, o or P4) at 5°C overnight, washed, incubated with secondary antibody conjugate at room temperature for 30 min, washed again, and incubated with chemiluminescence substrate at room temperature for 5 min. Chemiluminescence was visualized by using a photoimager system. p
Sodium phosphotungstic acid WB
The sodium phosphotungstic acid (NaPTA) WB was performed as previously described. 42 Briefly, tissue (0.2–0.3 g) was homogenized with spleen lysis buffer (10 mM MgCl2, 0.5% N-lauroylsarcosine, 100 mM sodium chloride, and 2% Tergitol in 50 mM Tris—HCl buffer pH 7.4) to generate a 10% homogenate. One and a half milliliters of the homogenate was transferred to a 1.5-ml conical centrifuge tube and centrifuged at 100 × g for 1 min. Thereafter, 500 μl of the supernatant was transferred to a bijoux vial, 500 μl of 4% N-lauroylsarcosine in Dulbecco phosphate buffered saline solution (PBS) was added, and the solution was incubated for 10 min at 37°C. Then, 100 μl of 11.5 mM MgCl2 in Dulbecco PBS and 50 μl of 50 units/ml of benzonase in Dulbecco PBS was added and incubated for 30 min at 50°C. Fifty microliters of proteinase K was added and incubated for a further 60 min at 50°C. Then, 100 μl of NaPTA was added and incubated for 30 min at 37°C. The entire solution was transferred to a 1.5-ml conical centrifuge tube and centrifuged at 15,800 × g for 30 min at 10°C. The supernatant was discarded, and the pellet was resuspended in 200 μl of 0.1% N-lauroylsarcosine in Dulbecco PBS and 200 μl of 250 mM ethylenediamine tetra-acetic acid in Dulbecco PBS and centrifuged at 15,800 × g for 30 min at 10°C. The supernatant was discarded, and the pellet was resuspended with 20 μl of 0.1% N-lauroylsarcosine in Dulbecco PBS, after which 20 μl of screening WB p sample buffer was added. Samples were then stored overnight at −20°C. The samples were thawed and boiled at 100°C before electrophoresis; transfer; immunodetection with mAbs SHA31, P4, and 6H4 as described under screening WB; and visualized on a photoimager system. p
Screening WB
The screening WB q was performed according to the procedure given by the manufacturer. Briefly, samples were homogenized, proteinase K digested, heat denatured in sample buffer, run in 12% gels m n at 200 V for 36 min, and transferred to PVDF membranes at 100 V for 30 min. r Protein molecular weight markers and a normal brain homogenate control sample were run on every gel. Membranes were blocked, incubated overnight at 4–8°C with the first antibody, washed, and incubated with the second antibody conjugate at room temperature for 60 min, washed, incubated with luminescence buffer, and incubated with substrate s at room temperature. The membranes then were sandwiched between transparent plastic sheets and images were taken with an imager. i
Histopathology and/or IHC
For histopathology and IHC, samples of fixed brain tissue were trimmed, embedded in paraffin wax, and sectioned at 4 μm. The brainstem sample fixed at the time of selection was processed and embedded at IDC by using standard histologic methods. Further fixed material was generated from a frozen slice of basal ganglion by immersion of the tissue in ice-cold 10% formol saline solution and then allowed to warm to room temperature and fix for a further 3 days before processing and embedding. Sections for histopathology were stained with hematoxylin and eosin. Detection of abnormal prion protein by IHC was performed by using the mouse mAb 2G11 t raised against the ovine PrP peptide sequence 146-R154R171-182, as previously described. 37,39 In brief, sections were immersed for 30 min in undiluted formic acid, then washed in running tap water for 15 min, followed by autoclaving at 121°C in 10 mM citrate buffer pH 6.1. Endogenous peroxidase was blocked by using 3% hydrogen peroxide in methanol, and the washing buffer used throughout the procedure was 50 mM Tris-buffered saline solution, supplemented with 0.2% Tween 20. The primary antibody was applied at a dilution of 1:400 for 1 hr at room temperature, with immunodetection performed by using biotinylated goat anti-mouse and avidin—biotin—peroxidase complex technique u by using diaminobenzidine chromogen prepared in McIlvane citrate buffer. Sections were counterstained by using Mayer hematoxylin, then routinely dehydrated, cleared, and mounted in dibutyl phthalate in xylene before examination by light microscopy.
Genotyping
At VLA UK, scrapie genotyping for sheep no. 1512 was performed by sequencing the full open reading frame (ORF) of the ovine PrP gene. 36 A 1.1-kbp region of the ovine PrP gene, including the ORF, was amplified by polymerase chain reaction (PCR; by using the PCR primers indicated below) from DNA extracted from ovine brain tissue (1 mm2) by using a tissue kit. v The 1.1-kbp PCR product generated then was visualized on an agarose gel. The PCR products were purified by using size exclusion technology and quantified by absorbance. The purified PCR products were used in the cycle sequencing reactions w to generate sequencing products by using 6 sequencing primers (F1X, F2, F3, R1, R2, and R3; see below). After ethanol purification, the samples were run on a genetic analyzer. x The sequence data generated from the 6 sequencing primers for each sample were assembled and analyzed by using specific software y that aligns the data to the wild-type sequence. Polymorphisms were automatically detected by the software. The following PCR primers were used: P3 F8871: 5′-ATTTATGACCTAGAATGTTTA TAGCTGATGCCACT-3′, P3 F8872: 5′-CCAGTTTTGT TTTTTTGAATGAATATTATGTGGC-3′. The following sequencing primers were used: F1x primer F 8856: 5′-TGACCTAGAATGTTTATAGCTGATGCCACTG-3′, F2 primer F 8865: 5′-CATGGTGGTGGAGGCTG GGGTC-3′, F3 primer F 8869: 5′-GGGAGAACTT CACCGAAACTGACATCA-3′, R1 primer R 8857: 5′-TTGAATGAATATTATGTGGCCTCCTTCCAGA-3′, R2 primer R 8866: 5′-GCTTCATGTTGGTTTTTGGCT TACTGG-3′, R3 primer R 8867: 5′-GGATTCTCTCT GGTACTGGGTGATGCA-3′.
Results
Overview of results
This investigation started on September 2, 2009, when IDC had received all the information from IRMM on the positive ELISA-2 results of a macerate that had been made from brainstem and cerebellum, and the negative results in ELISA-3 of a slice of brainstem only, both from sheep no. 1512 from New Zealand. With the brainstem tissues available from this sheep, classical scrapie and BSE could be ruled out early in the investigation. The laboratory investigation to confirm atypical scrapie/Nor98 was hindered by the limited availability of appropriate brain tissues, such as intact fresh and formalin-fixed cerebellum. The geographic locations and movements of tissues between various institutions and the numerous tests that had been performed over time are shown in Figure 1. Numbers in square brackets give the chronologic order of testing.
Testing of brain tissues at IDC before shipping to IRMM
A small slice of brainstem posterior to the obex and a slice from the cerebellum had been retained as fresh tissue reference material at IDC. A small slice of the brainstem anterior to the obex had been fixed in formalin and also retained at IDC.
In April 2008, before shipping of the brains, the slice of posterior brainstem had been tested by ELISA-1 at IDC, with negative results. The OD of the sample was 0.017 (positive control OD = 1.956; negative control OD = 0.008; cutoff OD = 0.148). The average OD of the 200 brainstem samples was 0.017 ± 0.015 (mean ± standard deviation). This ruled out the presence of classical scrapie and BSE in this brain. According to evaluation data for ELISA-1 and ELISA-2 (EFSA: 2005, EFSA Sci Rep 31:1–17), abnormal atypical scrapie/Nor98 prion protein should have been detected in the brainstem if it had been present. Hence, there was no suspicion of abnormal prion at this time in samples from sheep no. 1512.
Brain processing and initial testing in Europe
Between July and September 2009, IRMM organized the processing and testing of brain tissue samples into macerates, carried out by VLA UK, and, in parallel, as a routine procedure, the testing of brainstem slices by the National Veterinary Institute by using ELISA-3. Results on the brainstem slice of sheep no. 1512 gave negative results with an OD of 0.043 (positive control OD = 3.368; negative control OD = 0.036; cutoff OD = 0.016). A later retest of the remainder of this sample after rehomogenization also gave a negative result, with an OD of 0.051 (positive control OD = 3.261; negative control OD = 0.030; cutoff OD = 0.049) when using ELISA-3.
All cerebellum material and most of the brainstem held by IRMM (totaling 20 g) from sheep no. 1512 were used to make a macerate at VLA. This macerate repeatedly tested positive in independent test runs in the ELISA-2, with an OD of 0.937 and 0.948 (positive control ODs = 2.599 and 2.011; negative control ODs = 0.009 and 0.006; cutoff ODs = 0.219 and 0.216).
Testing at IDC during investigation after September 2, 2009
Initial results at IDC were difficult to interpret. ELISA-1 presented a very strong positive reaction with the cerebellar tissue (OD = 4.148), but a negative reaction with the brainstem (OD = 0.041; positive control OD = 1.226; negative control OD = 0.007; cutoff OD = 0.147). Subsequent testing in the screening WB assay showed only weak staining patterns for both tissues, which appeared to be nonspecific. The patterns resembled incompletely digested normal brain tissue (data not shown).
Further testing in the confirmatory WB showed a clear staining pattern characteristic of atypical scrapie/Nor98 in the cerebellum, and negative results for the brainstem (Fig. 2, blot A). Results from this blot were initially treated as inconclusive, because the atypical scrapie/Nor98 control did not produce a signal. In a repeated blot run on September 11, 2009, the staining pattern of the cerebellum showed high similarity to the atypical scrapie/Nor98 control (data not shown). The confirmatory WB results corroborated the previous ELISA-1 and ELISA-2 results.
At the request of IDC, half of the brain material still available at IRMM was returned to New Zealand at the end of September 2009. From this, the parts of the brain that could be expected to harbor abnormal atypical scrapie/Nor98 prion protein, such as frontal cerebral cortex (FCC), parietal cerebral cortex (PCC), and caudal cerebral cortex (CCC), as well as spinal cord as control material, were tested at IDC by using ELISA-2 with negative results: OD FCC = 0.016, OD PCC = 0.017, OD CCC = 0.019, and OD spinal cord = 0.025 (positive control OD = 1.723; negative control OD = 0.009; cutoff OD = 0.219). Confirmatory WB testing also gave negative results (Fig. 2, blot B). The atypical scrapie/Nor98 controls in this blot showed patterns identical to the staining pattern of the cerebellum of sheep no. 1512 in the initial confirmatory WB at IDC (Fig. 2, blot A).
Results at IDC from ELISA-1, ELISA-2, and confirmatory WB showed abnormal prion protein indicative of atypical scrapie/Nor98 only within the cerebellum and not in other relevant parts of the brain. In view of this, confirmatory IHC testing on fixed tissue was requested at VLA UK to confirm the presence of abnormal prion proteins indicative of atypical scrapie/Nor98 in sheep no. 1512. The IDC also requested that the confirmatory WB be conducted on the remaining macerate of combined brainstem and cerebellum and that the ELISA-3 be conducted on the macerate as well. Unfortunately, there was no fixed or fresh whole cerebellum available to be used at VLA UK.
Testing at VLA
The formalin-fixed brainstem from anterior to the obex that had been retained in New Zealand was sent to VLA UK. A slice of the frozen tissue that represented the frontal cortex and basal ganglia, held at IRMM, was also sent to VLA UK, and the basal ganglion from one side was then fixed in formalin. The remainder of the slice was used for further WB testing. All fixed tissues were subjected to histopathology and/or IHC, and each returned negative results. Classical scrapie and BSE were ruled out. ELISA-3 performed on the macerate also gave a negative result, with an average sample OD from 4 aliquots of 0.067 (positive control OD = 2.818; negative control OD = 0.062; cutoff OD = 0.242). The confirmatory WB on brainstem and cerebellum macerate, and frontal cortex and basal ganglion, showed very weak staining patterns suggestive of atypical scrapie/Nor98 with 2 antibodies (SHA31 and P4; Fig. 3). With mAb SHA 31, there was a faint diffuse molecular mass banding clearly visible after the image was enhanced. This diffuse band was in a similar molecular mass position to the top band of the atypical scrapie/Nor98 control. With mAb P4, low molecular mass proteinase K—resistant bands were clearly visible on enhancement of the image. These appeared to bear some resemblance to the position of the lower molecular mass bands of the atypical scrapie/Nor98 control. These bands were seen in the 3 macerate samples but also in the forebrain sample, albeit with differing intensity.
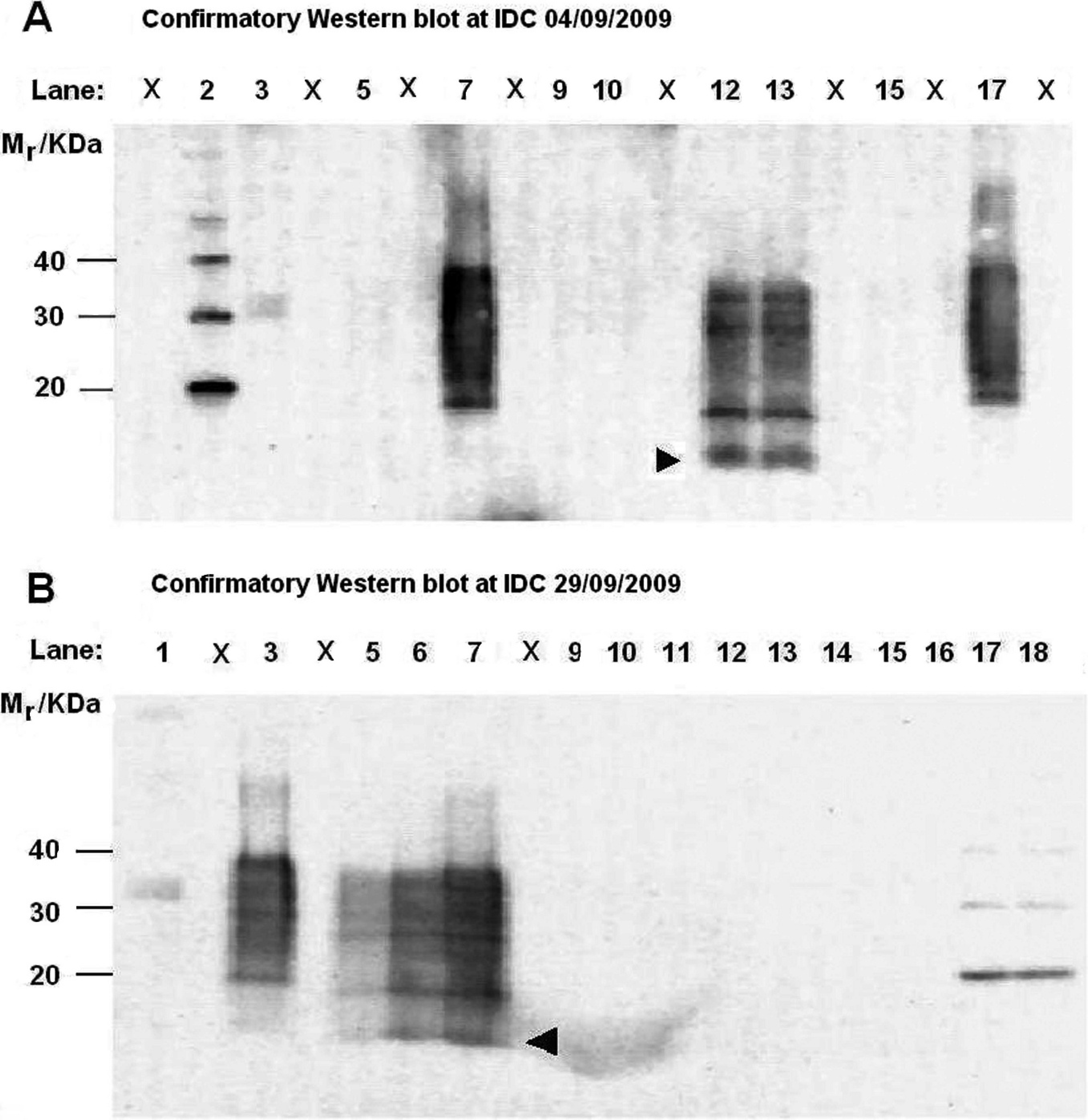
Confirmatory Western blot (WB) tests performed at the Investigation and Diagnostic Centre on various tissues from sheep no. 1512 on September 4 and 29, 2009. Relative molecular mass (Mr) for MagicMark markers are shown on the left. Arrowhead indicates low Mr in atypical scrapie/Nor98 cases.
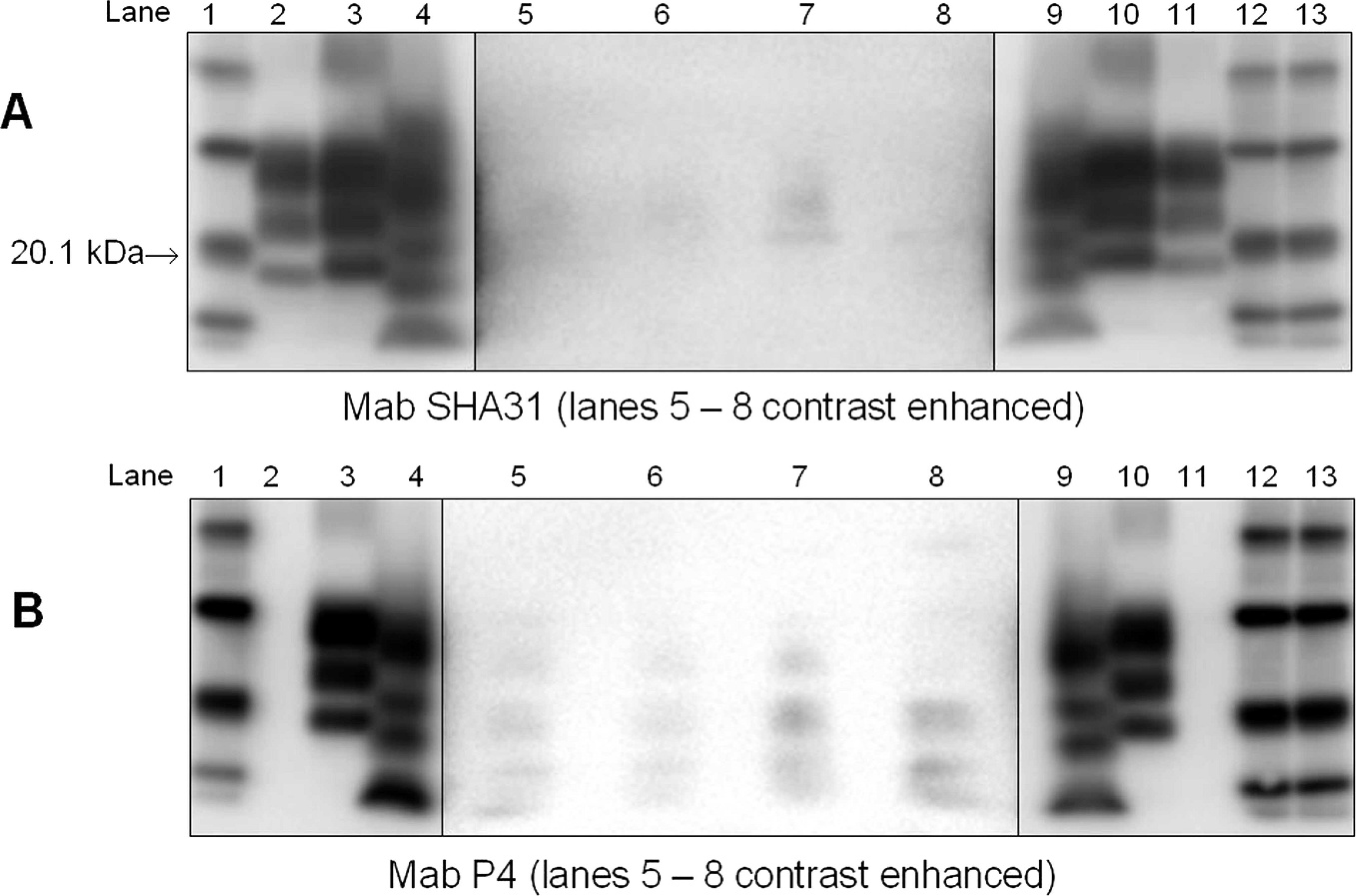
Confirmatory Western blot tests performed at the Veterinary Laboratories Agency on various tissues from sheep no. 1512. Lanes 1, 12, 13: molecular weight marker; lanes 2, 11: bovine spongiform encephalopathy positive control; lanes 3, 10: classical scrapie positive control; lanes 4, 9: atypical scrapie/Nor98 positive control; lanes 5–7: various macerate samples from sheep no. 1512; lane 8: forebrain sample from sheep no. 1512. Note: To improve visualization of the weak signal detected in the samples, lanes 5–8 have been contrast enhanced in these images compared with the controls.
Although neither mAb detected a full and clear profile that corresponded to either an atypical or classical scrapie profile, there was definitely some form of proteinase K—resistant protein present in the 4 samples. Therefore, at this stage, the WB results were considered inconclusive, and further testing by the confirmatory methods, OIE-SAF WB and NaPTA WB, was performed.
Results of the OIE-SAF WB are shown in Figure 4. The macerate sample showed clear detection of PrPSc with mAbs SHA31 and P4, whereas mAb 6H4 showed a weak detection that was more visible on the enhanced contrast image.
Monoclonal antibody SHA31 showed the strongest signal of the 3 antibodies with the fresh tissue sample from the forebrain, but this sample was extremely weak in comparison with the macerate. Again, the enhanced contrast image was clearer. The band profile for the samples from sheep no. 1512 matched closely to the known positive atypical scrapie/Nor98 control. Overall, sheep no. 1512 was considered positive for atypical scrapie/Nor98 by the OIE-SAF WB assay. The NaPTA WB was considered invalid because the control ovine negative samples exhibited residual PrP c from underdigestion with proteinase K. Because the OIE-SAF WB had already given conclusive results, it was decided not to repeat this method.
Genotyping
Comparison of a sample from sheep no. 1512 with a wild-type ovine PrP ORF (GenBank no. AY350267) showed that sheep no. 1512 is of the genotype AA136 LL141 RR154 RR171. Compared with the wild type, in nucleotide position 512, adenine was replaced by guanine, leading to a difference in the amino acid position 171 of arginine (R) instead of glutamine (Q) in the wild type. These are commonly observed mutations. No other differences in sequence were observed.
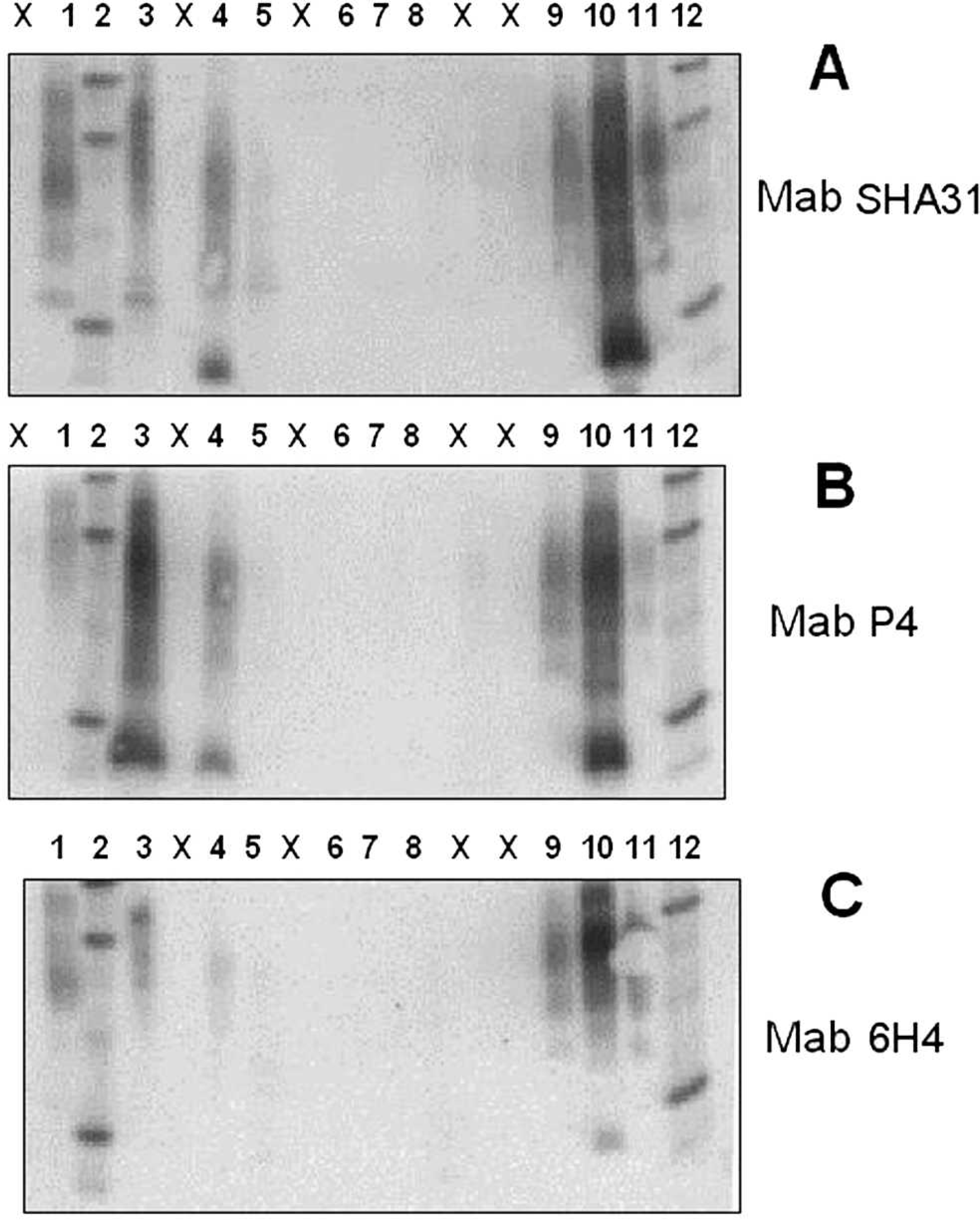
World Organization for Animal Health scrapie-associated fibril Western blot tests performed at the Veterinary Laboratories Agency on various tissues from sheep no. 1512. Lane 1: kit control p (undigested normal brain tissue); lanes 2, 12: molecular weight marker; lane 3: macerate from sheep no. 1512, undigested; lane 4: macerate from sheep no. 1512, proteinase K digested; lane 5: forebrain homogenate from sheep no. 1512; lane 6: negative New Zealand sheep control sample; lane 7: negative U.K. sheep control sample; lane 8: negative U.K. bovine BSE (bovine spongiform encephalopathy) sample; lane 9: classical scrapie control sample; lane 10 = atypical scrapie/Nor98 control sample; lane 11: bovine BSE control sample. X = blank lanes. Panels A, B, and C were processed identically except for using different monoclonal antibodies.
Discussion
There are distinct differences in the staining patterns in WBs for the abnormal prion proteins of classical scrapie and atypical scrapie/Nor98. After proteinase K treatment, all classical scrapie strains and BSE prions show a typical triplet pattern that comprises the di-, mono-, and unglycosylated bands migrating between 18 and 30 kDa. 19,20 Such patterns are readily detectable in the brainstem. Neither classical scrapie nor BSE was present in sheep no. 1512.
The WB patterns for atypical scrapie/Nor98 include multiple protein bands of different sizes, with a characteristic unglycosylated low molecular-weight band below 15 kDa. 2,4,5,16,18,22 A pattern characteristic of atypical scrapie/Nor98 was clearly seen in the cerebellum from sheep brain no. 1512 in the confirmatory WB. The WB pattern for the macerate in the OIE-SAF WB conclusively confirmed this first case of atypical scrapie/Nor98 in a New Zealand sheep.
In the majority of atypical scrapie/Nor98 cases reported to date, abnormal prion protein has been found in the cerebellar and cerebral cortex, and in the brainstem at lower concentration. 4,29,32 This may reflect true anatomical diversity or, alternatively, may be the result of a biased sample, in which only sheep that were not negative when the brainstem was tested with a rapid test then were subjected to further detailed testing. The only way to clarify the prevalence of anatomical distribution would be to compare matched cerebellum and brainstem samples. Sheep no. 1512 appears to be a case in which abnormal prion protein was present almost exclusively in the cerebellum and was not initially detected by rapid testing of brainstem material from anterior to the obex.
ELISA-3, together with the ELISA-2 and ELISA-1, is 1 of 3 rapid screening tests recommended by EFSA for atypical scrapie/Nor98 testing on brainstem and cerebellum, even though this evaluation was based on only 3 samples (EFSA: 2005, EFSA Sci Rep 31:1–17). In December 2009, EFSA reported an assessment of the analytical sensitivity for all the currently approved TSE rapid tests for BSE, classical scrapie, and atypical scrapie/Nor98 (Scientific opinion on analytical sensitivity of approved TSE rapid tests. EFSA Panel on Biological Hazards. European Food Safety Authority, Parma, Italy, 2009. Available at http://www.efsa.europa.eu/en/scdocs/doc/s1436.pdf). Contrary to previous evaluations, each test was evaluated against the same sample sets, which allowed a direct comparison of their analytical sensitivity. For classical scrapie, all 9 screening test methods examined, including ELISA-1, ELISA-2, ELISA-3, and the screening WB, performed equally well. With regard to atypical scrapie/Nor98, ELISA-1, ELISA-2, and ELISA-3 performed well within the set limits of maximal 2-log10 inferiority range compared with the most sensitive test system.
A slight sensitivity difference between ELISA-1 or ELISA-2, and ELISA-3 at high dilutions was observed, which, particularly with a sample with the characteristics described herein from sheep no. 1512, may lead to the discrepant results among the immunoassays, and it emphasizes the importance of selecting the most appropriate type of tissue for testing. Furthermore, from previous evaluations, it has been observed that homogenization of tissue can have a great influence on the apparent analytical sensitivity, because of artifacts that may have been introduced by the homogenization procedure (Schimmel H, Catalani P, Le Guern L, et al.: 2002, Report on the evaluation of 5 rapid tests for the diagnosis of transmissible spongiform encephalopathy in bovines (2nd study). Reference Materials Unit, Institute for Reference Materials and Measurements, Joint Research Centre, Geel, Belgium. Available at http://ec.europa.eu/food/food/biosafety/bse/bse42_en.pdf). Whatever the reasons may be, in a recent publication on the first 6 atypical scrapie/Nor98 cases in the United States, at least 2 cases in which brainstem and cerebellum were positive by ELISA-2 but negative by the ELISA-3 have been reported. 25
The genotype of the PrP gene from sheep no. 1512 was determined as ALRR/ALRR. The ARR genotype is the second most common genotype in New Zealand sheep 24 and also the second most common in Romney sheep, the breed of sheep no. 1512. The national flock in New Zealand consists of approximately 48% ARQ, 36% ARR, 10% AHQ, 5% VRQ, and 2% ARH genotypes. 24
Three cases of atypical scrapie/Nor98 had been detected in 2006 and 2007 in Cheviot sheep of New Zealand lineage or origin in a research sheep flock in the United Kingdom. 37 The sheep were of the genotypes AFRQ/AFRQ, AFRQ/ALRQ, and AFRQ/AFRQ, thus, belonging to the most common PrP genotype in New Zealand sheep. The most likely explanation for these 3 cases of atypical scrapie/Nor98 was that they arose spontaneously. Consistent with most atypical scrapie/Nor 98 cases, the 3 sheep from the U.K. sheep unit were of advanced age: 6, 8, and 9 years old. Sheep no. 1512 was more than 3 years old but its exact age is not known.
Sheep with atypical scrapie/Nor98 are often found to carry phenylalanine (F) rather than leucine (L) in position 141 of the PrP gene, which seems to confer increased susceptibility to atypical scrapie/Nor98. 30,31,36 The 3 cases in the U.K. sheep flock belonged to this more susceptible genotype, whereas case no. 1512 was of the less susceptible genotype regarding position 141.
The U.K. National Scrapie Plan groups PrP genotypes into 5 groups according to their resistance to classical scrapie 36 and promotes breeding for resistance. Most resistant are group 1 genotypes (ARR/ARR), group 2 genotypes (ARR/AHQ, ARR/ARH, ARR/ARQ), and group 3 genotypes (AHQ/AHQ, AHQ/ARQ, ARQ/ARQ). The majority of New Zealand sheep fall into these 3 classical scrapie—resistant groups. As an irony of nature, atypical scrapie/Nor98 is most commonly found in these 3 genotype groups. What impact these genotype differences in resistance for classical scrapie and atypical scrapie/Nor98 on eradication policies in Europe and elsewhere will have remains to be seen. 41
The suggestion of a spontaneous etiology for atypical scrapie/Nor98 with possible genetic determinants 12–15,17,21 would mean that, in New Zealand, with a sheep flock of approximately 30 million and with PrP genotypes favorable for this condition, it is likely that more atypical scrapie/Nor98 cases could be discovered during routine scrapie surveillance. Whatever the causes, atypical scrapie/Nor98 was clearly able to induce pathology in the New Zealand nonintensive, pasture-based system and hence may not be associated with more intensive farming methods. In the 2,484 New Zealand sheep and goat brains sent to the European Union for rapid test evaluations (Table 1), only 1 brain (no. 1512) was detected with atypical scrapie/Nor98, which confirmed that this condition is indeed a rare event. Implications for other countries that are scrapie-free are that a case of atypical scrapie/Nor98 may eventually be detected there. A first case of atypical scrapie/Nor98 has been reported from Australia in March 2010 (report in preparation for Australian Animal Health Surveillance 2 Quarterly, October 2010).
In summary, this first case of a sheep with atypical scrapie/Nor98 in New Zealand was conclusively confirmed by applying a number of confirmatory WB assays, despite the limitation of available tissues. In this interesting case, the abnormal prion protein was mainly detected in the cerebellum, with low concentration in the forebrain and none in the brainstem. The genotype of this sheep is consistent with the majority of genotypes of sheep from New Zealand, which are of the types resistant to classical scrapie but more likely to develop atypical scrapie/Nor98. In future shipments of brains to the European Union, care needs to be taken to retain not only brainstem tissues but also other relevant tissues from small ruminant brains, such as fixed cerebellum and forebrain in sufficient amounts.
Acknowledgements
The authors thank staff from all laboratories involved in the testing of the brain samples. Furthermore, the authors would like to acknowledge the contributions made by various teams from MAFBNZ, the NZ Food Safety Authority, and AsureQuality, and from stakeholders who have been involved in this investigation. The authors also thank Dr. Peter Lind from the National Veterinary Institute, Technical University of Denmark (Copenhagen, Denmark), for ELISA-3 testing, and Dr. Richard Ellis, VLA UK, for performing the genotyping.
Footnotes
a.
TeSeE Sheep and Goat ELISA detection kit, Bio-Rad Laboratories, Hercules, CA.
b.
TeSeE Ruminant ELISA detection kit, Bio-Rad Laboratories, Hercules, CA.
c.
HerdChek BSE-scrapie ELISA, IDEXX Laboratories Inc., Westbrook, ME.
d.
Hand-held blender, Cookworks, Vancouver, Canada.
e.
Precess 24 ryboliser, Bio-Rad, Marnes-la-Coquette, France.
f.
Eppendorf Centrifuge 5804R, Eppendorf, Hamburg, Germany.
g.
TeSeE Western blot confirmatory assay, Bio-Rad Laboratories, Marnes-la-Coquette, France.
h.
Criterion Electrophoresis and Western Blot System, Bio-Rad Laboratories, Hercules, CA.
i.
Chemi-Doc System 2000, Bio-Rad Laboratories, Hercules, CA.
j.
R-Biopharm AG, Southmarshall, MI.
k.
Omni general laboratory Homogenizer, Omni International, Kennesaw, GA.
l.
SpeedVac system, Thermo Fisher Scientific Inc., Waltham, MA.
m.
Bis-Tris NuPage gels, Invitrogen Ltd, Paisley, United Kingdom.
n.
X cell Sure lock Mini-Cell, Invitrogen Ltd, Paisley, United Kingdom.
o.
Prionics AG, Schlieren, Switzerland.
p.
Fluor S Multimager, Bio-Rad Laboratories, Hercules, CA.
q.
Prionics Check Western small ruminant kit, Prionics AG, Schlieren, Switzerland.
r.
Criterion blotter, Bio-Rad Laboratories, Hercules, CA.
s.
CDP-Star, Roche Diagnostics GmbH, Mannheim, Germany.
t.
Institute Pourquier, Montpellier, France.
u.
Vectastain Elite ABC kit, Vector Laboratories Ltd, Orton Southgate, United Kingdom.
v.
Qiagen DNeasy™, Qiagen Ltd., Crawley, United Kingdom.
w.
BigDye v3.1, Applied Biosystems, Carlsbad, CA.
x.
ABI Prism 3730 Genetic Analyzer, Applied Biosystems, Carlsbad, CA.
y.
Seqscape, Applied Biosystems, Carlsbad, CA.