
Abstract
Herpesviruses are important pathogens of chelonians, and include Chelonid herpesvirus 5, which is associated with fibropapillomatosis in sea turtles. Spirorchid trematodes are blood flukes that reside within the cardiovascular system of marine turtles and may be associated with severe disease. An eastern box turtle (Terrapene carolina) at the South Florida Wildlife Care Center (Fort Lauderdale, Florida) was presented to the facility with papillomatous growths behind both rear legs. Surgical removal resulted in remission for 8 months; however, lesions recurred, prompting a second surgery and acyclovir therapy. Surgical biopsies revealed subacute superficial inflammation associated with the supporting stroma of the cutaneous papillomas and granulomas within the superficial dermis containing fragmented and collapsed brown trematode eggs surrounded by multinucleated giant cells and epithelioid macrophages. Pan-herpesviral and pan-trematode consensus polymerase chain reaction and sequencing were run on tissue samples. Comparative sequence analysis revealed a novel alphaherpesvirus and a novel trematode in the genus Spirorchis. The animal became anorexic and was euthanized due to poor quality of life. While we do not yet have a complete understanding of the effects of herpesvirus and trematode infections in eastern box turtles, the findings thus presented provide initial insights into the disease relationships among these chelonians.
Eastern box turtles (Terrapene carolina) are North American turtles in the family Emydidae. These turtles are a nearly threatened species according to the International Union for Conservation of Nature, with the population decreasing as a result of nest disturbance, habitat loss, and moving vehicles. 2 With low fecundity, low juvenile survival rates, and long adult life spans, eastern box turtles have a life history that predisposes them to impaired population recovery following the loss of adult animals (such as loss by disease). 8 Known diseases affecting eastern box turtles include ranaviral disease and upper respiratory tract disease caused by a Mycoplasma sp.6,10 Adenoviruses and a herpesvirus have been identified in box turtles in the past 2 years.5,18 The prevalence and manifestations of infectious agents, many of which likely remain undocumented, are not fully understood in eastern box turtles.
Herpesviruses are of significant concern in a variety of chelonian species involved in rehabilitation, conservation, and breeding efforts. 21 Herpesviruses have typically coevolved with their host organisms, and aberrant host species are more strongly affected by disease than endemic host species.3,24 Herpesviruses were initially classified based on the host species, morphology, and disease signs. However, herpesviruses are now categorized by genetic sequence and phylogenetic analyses. 4 The family Herpesviridae is categorized into the subfamilies Alphaherpesvirinae, Betaherpesvirinae, and Gammaherpesvirinae, which are further divided into distinct genera and species.4,21 The subfamily Alphaherpesvirinae contains all herpesviruses infecting non-avian reptiles. Individuals of the Emydidae family have been reported to develop disease in association with herpesvirus-like particles. In various tortoise species, herpesviruses are known to cause tracheitis, stomatitis, pneumonia, and conjunctivitis. 25 There has been previous histologic evidence suggesting herpesviral infections in emydid turtles 25 ; and a novel alphaherpesvirus was reported in 2014 in another emydid, an eastern river cooter (Pseudemys concinna concinna). 12 Although the freshwater turtle in that report showed no antemortem clinical signs, microscopic examination revealed hepatocytes containing intranuclear inclusion bodies and herpesviral particles. 12
The superfamily Schistosomatoidea contains blood flukes in the families Sanguinicolidae, Schistosomatidae, and Spirorchiidae. Spirorchid flukes in turtles are similar to schistosomes in birds and mammals. Trematodes within the family Spirorchiidae include 16 recognized genera, and reside within the cardiovascular system of turtles. The circulatory system transports eggs (deposited by adults) that cause blockages within blood vessels and organs, which in turn cause inflammation and microgranulomas.11,19 Whereas freshwater pulmonate snails are known to be intermediate trematode hosts among freshwater turtles, the life cycle for marine spirorchids is not fully understood. The marine spirorchid, Learedius learedi, was detected within a potential intermediate host, the limpet Fissurella nodosa. 20 To our knowledge, spirorchid fluke eggs have not previously been reported in box turtles, and with secondary infections potentially leading to death, this holds clinical significance for box turtle health. 11 Thus far, information on spirorchids in turtles is limited, as most species have yet to be examined for these blood flukes. 19
Our project aimed to characterize a novel herpesvirus and a novel Spirorchis sp. found in an eastern box turtle. The results provide a better understanding of the relationships of these agents and enable predictions using close relatives as models.
An adult eastern box turtle was presented to the South Florida Wildlife Care Center (Fort Lauderdale, Florida) in October 2012 with papillomatous skin lesions on the cranial aspect of the proximal hind limbs near the junction with the plastron, and on the medial aspect of the lateral hind limb near the junction with the carapace (Fig. 1). The box turtle was sedated with ketamine, dexmedetomidine, and butorphanol for tissue sampling. Excisional biopsy samples were sectioned and a portion was frozen at -20°C; the remainder was placed in 10% neutral buffered formalin. The latter was shipped to Zoo/Exotic Pathology Service (West Sacramento, California) for histologic examination. Paraffin-embedded tissues were sectioned at ~5 µm, mounted on glass slides, and stained with hematoxylin and eosin. Frozen tissue sections were placed in a sterile cryotube and shipped with cold packs to the University of Florida (Gainesville, Florida) for molecular testing. Surgical excision resulted in remission for 8 months; however, lesions recurred, prompting a second surgery and acyclovir therapy

Gross appearance of box turtle fibropapillomas.
The animal became anorexic and was euthanized due to poor quality of life. No postmortem examination was performed due to financial constraints. DNA was extracted from the biopsy samples using a commercial kit. a Nested polymerase chain reaction (PCR) amplification of a partial sequence of the viral DNA–dependent DNA polymerase gene was performed on the DNA extract using previously described methods. 23 The product was resolved on 1% agarose gel and purified using a commercial extraction kit. b Sequences for primers used in our study are listed in Table 1. To obtain an additional sequence, 2 reverse primers, TerHV2R1 and TerHV2R2, were designed using the sequence obtained from the initial protocol. Forward primers DIEC, 14 DFA, 23 and SIIQ 14 were used in combination with the reverse primers TerHV2R1 and TerHV2R2 for PCR under identical conditions to those used in the first amplification. 23
Primers used in terrapene herpesvirus and spirorchid trematode sequencing.
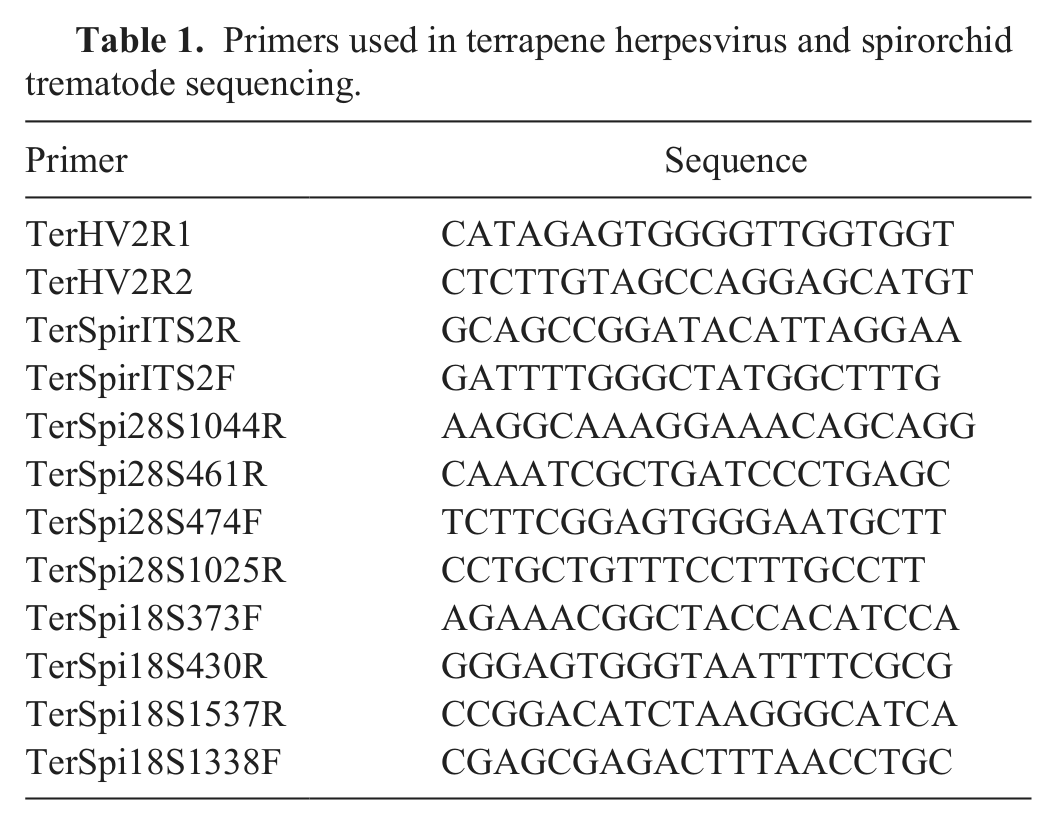
To obtain a spirorchid DNA sequence, amplification of the spirorchid internal transcribed spacer (ITS)2 region was performed using forward primer SPIR1 20 with reverse primer SPIR2. 20 To obtain the spirorchid 18S ribosomal (r)RNA gene sequence, the previously reported forward primer 18SE 19 was used with reverse primers TerSpirITS2R designed from the ITS2 sequence obtained and the previously reported WORMB. 19 Forward primers 18SE 19 and TerSpi18S1338F were used with reverse primer TerSpirITS2R. To obtain the 28S rRNA gene sequence, novel forward primers TerSpirITS2F and U178 19 were used with reverse primer L1642. 19 Forward primer TerSpirITS2F was used with reverse primers L1642 19 and TerSpi28S461R under identical conditions to those used in the first amplification. Direct sequencing was performed using a commercial kit c and analyzed on automated DNA sequencers. d To obtain a larger region of the spirorchid genome, additional sequencing was performed using primers 14–21 in Table 1. Primer sequences were trimmed off prior to further analysis.
Amino acid sequences from the herpesviral DNA-dependent DNA polymerase gene were aligned using MAFFT. 13 Partial homologous amino acid sequences for which full-length sequences were not available were included, with ambiguities added for unknown amino acids for gerrhosaurid herpesvirus 1 (59 aa), gerrhosaurid herpesvirus 2 (59 aa), gerrhosaurid herpesvirus 3 (60 aa), varanid herpesvirus 1 (58 aa), lacertid herpesvirus 1 (59 aa), tortoise herpesvirus 1 (60 aa), tortoise herpesvirus 2 (60 aa), tortoise herpesvirus 4 (141 aa), indotestudo herpesvirus (60 aa), cooter herpesvirus (60 aa), and red-eared slider herpesvirus (60 aa). Cooter and red-eared slider herpesviruses are not in GenBank due to the recent requirement of a minimum of 200 nucleotides for submission. Iguanid herpesvirus 2 (GenBank accession no. AY236869) was designated as the outgroup because of its early divergence from other herpesviruses.16,24 Bayesian analyses of amino acid alignments were performed using MrBayes 3.1.2 on the CIPRES server, with gamma distributed rate variation and a proportion of invariant sites (Miller MA, et al. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In: Proceedings of the Gateway Computing Environments Workshop (GCE), 14 Nov. 2010, New Orleans, LA, 2010;1–8). 17 Amino acid substitution models were selected using ProtTest. 1 The first 25% of 1,000,000 iterations were discarded from the beginning of the chain, hereafter referred to as a “burn in.” Maximum likelihood (ML) bootstrap analyses of each alignment were performed using RAxML on the CIPRES server with gamma distributed rate variation and a proportion of invariant sites (Miller MA, et al., 2010, Creating the CIPRES Science Gateway). 22 Bootstrap analysis was used to test the strength of the tree topology, with 1,000 subsets. 7
Nucleotide sequences from the trematode 18S gene were aligned using MAFFT. 13 Sasala nolani (GenBank accession no. AY157184) was designated as the outgroup. Bayesian analyses of nucleotide alignments were performed using MrBayes 3.1.2, and ML bootstrap analyses of each alignment were performed using RAxML on the CIPRES server, with a general time reversible model, gamma distributed rate variation, and a proportion of invariant sites (Miller MA, et al., 2010, Creating the CIPRES Science Gateway).17,22 The first 25% of 1,000,000 iterations in the Bayesian analysis were discarded as a burn in. Bootstrap analysis was used to test the strength of the tree topology, with 1,000 subsets. 7
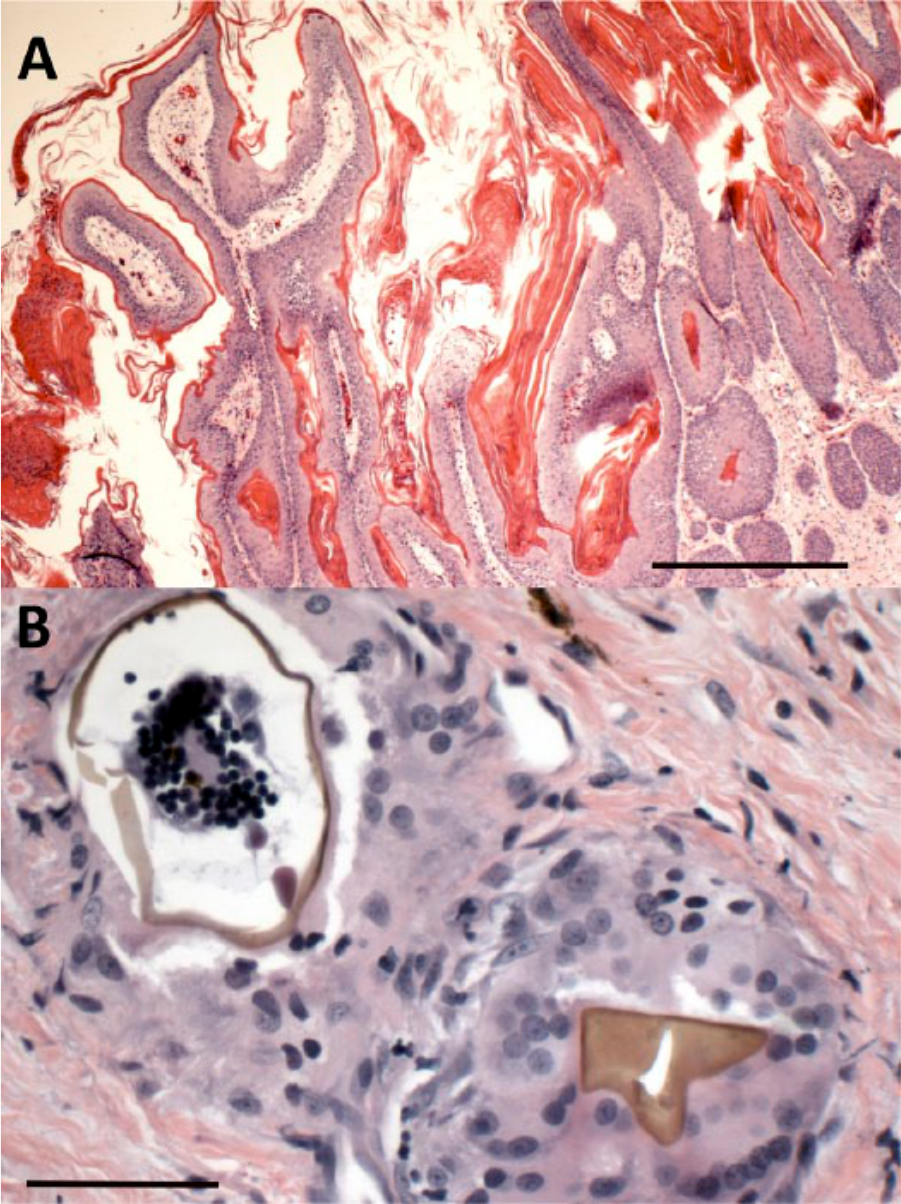
Histologic examination of the eastern box turtle sample revealed variable papillary hyperplastic stratified epithelium supported on cores of fibrous connective tissue infiltrated by lymphocytes, plasma cells, and heterophils. Dense compressed keratin and crusts of keratin and cell debris covered the epithelium (Fig. 2A). The superficial dermis contained occasional granulomas consisting of fragmented and collapsed brown trematode eggs surrounded by multinucleated giant cells and epithelioid macrophages. The shells were smooth, and rare aggregates of globular basophilic and eosinophilic contents were within the egg (Fig. 2B).
PCR amplification of the partial DNA-dependent DNA polymerase gene yielded a 460 base pair (bp)-product after editing out primers. PCR amplification of the spirorchid 28S gene and ITS region resulted in a 2,046-bp product after editing out primers. PCR amplification of the spirorchid 18S gene resulted in a 1,905-bp product after editing out primers. Sequences were deposited in GenBank under accession numbers KM507472 and KM507473.
ProtTest analysis of the herpesviral alignment found that the LG model with gamma distribution and a portion of invariant sites was the best model, followed by the WAG model. LG is not implemented in MrBayes, so the WAG model was used for the Bayesian analysis. The Bayesian phylogenetic tree of the herpesviruses is displayed in Figure 3. The LG model was used for the ML analysis in RAxML. Bootstrap values as percentages from the ML analysis are shown in the Bayesian tree. The phylogenetic analysis shows that this virus, hereafter called terrapeneHV2 and indicated by the arrow in Figure 3, is most closely related to indotestudoHV, with a Bayesian posterior probability of 98% and a ML bootstrap value of 90%. These 2 viruses cluster closely with loggerheadOCV, with a Bayesian posterior probability of 91% and a maximum likelihood bootstrap value of 73%. TerrapeneHV2 has close genetic relationships with a clade of chelonian herpesviruses, including terrapeneHV1, tortoiseHV1, tortoiseHV2, tortoiseHV3, tortoiseHV4, cooterHV, and red-eared sliderHV, with a Bayesian posterior probability of 100% and a ML bootstrap value of 75%. The phylogenetic analysis supports classification of terrapeneHV2 within the family Herpesviridae, subfamily Alphaherpesvirinae. Branch lengths show genetic distances in support of the classification of terrapeneHV2 as a novel herpesviral species.
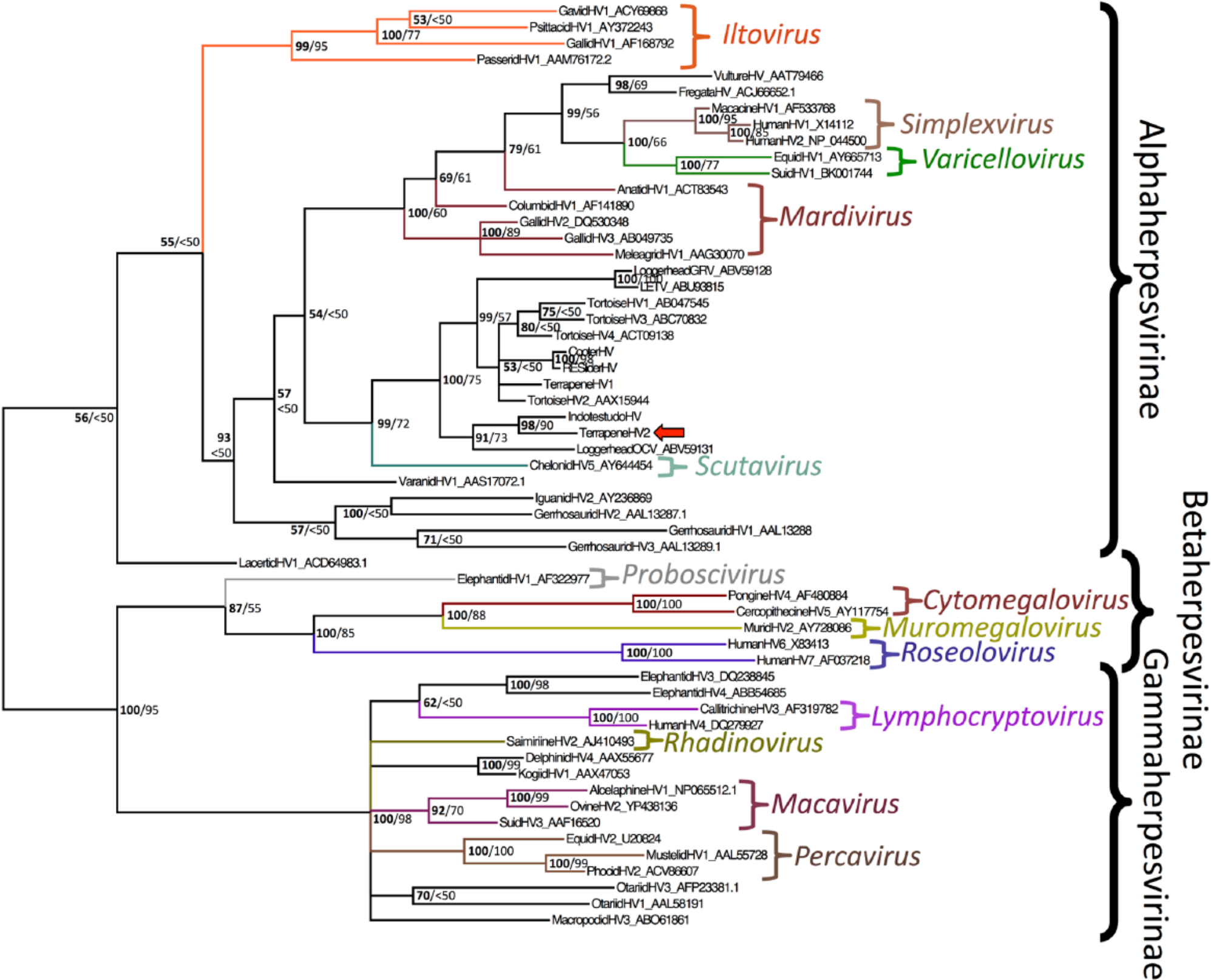
Bayesian phylogenetic tree of predicted amino acid sequences of herpesviral DNA–dependent DNA polymerase sequences based on MAFFT sequence alignment. Bayesian posterior probabilities of clusters as percentages are on the left, and maximum likelihood bootstrap values for clusters based on 1,000 resamplings are given to the right. Iguanid herpesvirus 2 was used as an outgroup. Terrapene herpesvirus 2 is indicated by the arrow. Brackets indicate genera. Accession numbers of sequences retrieved from GenBank are given after the name.
The Bayesian phylogenetic tree of the spirorchid trematodes is displayed in Figure 4. Bootstrap values as percentages from the ML analysis are shown in the Bayesian tree. The phylogenetic analysis shows that the terrapene spirorchid, indicated by the arrow in Figure 4, is most closely related to Spirorchis artericola, S. scripta, and S. haematobius, with a Bayesian posterior probability of 100% and a ML bootstrap value of 100%. Additionally, the terrapene spirorchid is shown to be the most basal of the described species in the genus, and the family Spirorchiidae appears to be paraphyletic. The phylogenetic analysis supports classification of the terrapene spirorchid within the family Spirorchiidae.
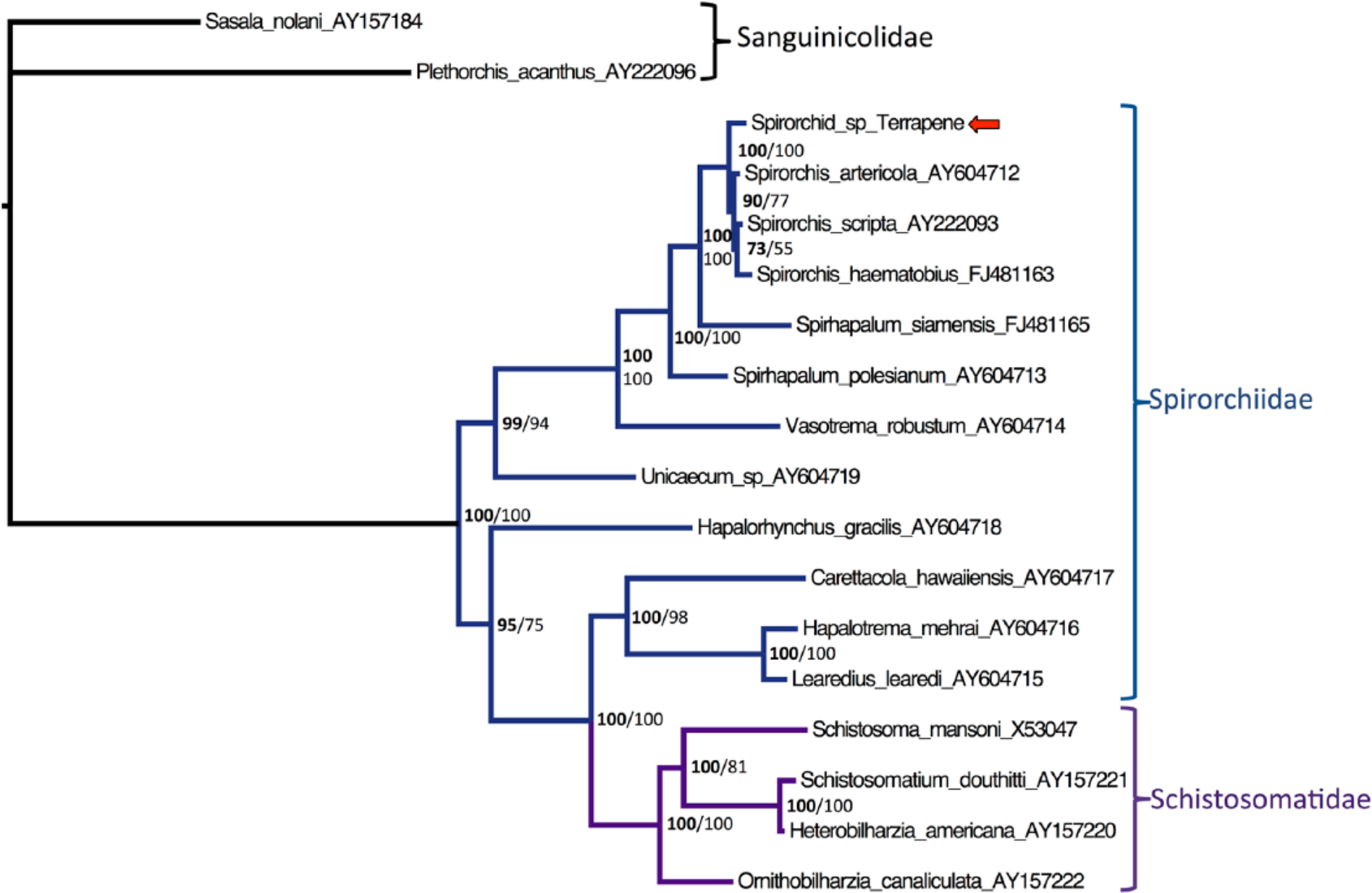
Bayesian phylogenetic tree of predicted nucleotide sequences of spirorchid trematode sequences based on MAFFT sequence alignment. Bayesian posterior probabilities of clusters as percentages are on the left, and maximum likelihood bootstrap values for clusters based on 1,000 re-samplings are given to the right. Sasala nolani was used as an outgroup. The terrapene spirorchid is indicated by the arrow. Brackets indicate genera. Accession numbers of sequences retrieved from GenBank are given after the name.
The phylogenetic tree in Figure 3 from the eastern box turtle in our study shows that terrapeneHV2 clusters with other herpesviruses in family Herpesviridae and subfamily Alphaherpesvirinae. Based on the similar structure of this phylogenetic tree to those from larger data sets and the fact that mammals are the only known hosts of subfamilies Betaherpesvirinae and Gammaherpesvirinae, terrapeneHV2 has support for classification within the subfamily Alphaherpesvirinae.16,21 The Bayesian posterior probability in our analysis of the known chelonian herpesviruses clustering together is 99%, with a ML bootstrap value (a more conservative method of estimation) of 72%. Based on branching patterns between herpesviruses and their host species, previous phylogenetic analyses are consistent with host–virus coevolution.15,21 Future research, including additional data sets and longer sequences for comparison, will help delineate the phylogenetic relationships between this novel herpesviral species and others. 21
The phylogenetic tree in Figure 4 shows that terrapene spirorchid clusters within the family Spirorchiidae. Spirorchiidae and Schistosomatidae families vary with respect to reproduction, morphology, and hosts. 19 Spirorchids reside basally on the phylogenetic tree, with schistomatids as a more recently derived species. Thus, the family Spirorchiidae is paraphyletic. Our study finds strong support from both Bayesian posterior probabilities and bootstrapping for the basal position of the Spirorchiidae clade (Bayesian posterior probability of 100% and a ML bootstrap value of 100%).
The box turtle that was presented to the South Florida Wildlife Care Center in October of 2012 had inflamed cutaneous papillomas characteristic of fibropapilloma-associated herpesviral infection. Several granulomas were found in the superficial dermis, typically associated with collapsed trematode eggs and more specifically, spirorchid fluke eggs. Reptilian viruses may be diagnosed through histology and further molecular testing. 25 Histologic evaluation and DNA sequencing confirmed the presence of papillomatous lesions containing herpesviral DNA and spirorchid fluke DNA.
Chelonians have the best studied herpesvirus flora of any non-avian reptile clade, although this primarily speaks to the paucity of information on non-mammalian herpesviruses. 21 Herpesviruses are known to cause neoplastic diseases and papillomas among vertebrates. 9 Common sites of fibropapilloma growth on turtles include the skin over the flippers, tail, jaw, and neck, as well as the eyes and inside the oral cavity or on the skin adjacent to the mouth. 9 The box turtle in the present case report displayed papillomatous growths, which had spread on the rear legs for several months prior to sampling. In addition to fibropapillomas, chelonians with herpesviruses may be afflicted with dermatitis, conjunctivitis, necrotizing hepatitis, stomatitis, and respiratory tract infections.9,21
For successful population management, it is important to identify the diseases present and the pathogenicity for a given host species. Our study findings represent the second herpesvirus and, to our knowledge, the first Spirorchis sp. documented in eastern box turtles. Coinfection of the novel herpesvirus and Spirorchis sp. may reveal valuable insights into disease characteristics and reasons for minimizing the mixing of certain species in captivity. Intermediate spirorchid hosts are not fully identified and thus, further studies of gastropods and other potential intermediate hosts are warranted. Because fibropapilloma-associated herpesvirus and spirorchid trematode infection routes and relationships are not fully understood in eastern box turtles, preventative measures such as isolation of infected individuals and quarantine of new turtles should be taken by animal handlers to eliminate potential disease transmission to healthy animals.
Footnotes
Authors’ contributions
SB Yonkers and JFX Wellehan Jr contributed to the conception and design of the study and drafted the manuscript. All authors contributed to acquisition, analysis, or interpretation of data; critically revised the manuscript; gave final approval; and agree to be accountable for all aspects of the work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
a.
DNeasy kit, Qiagen Inc., Valencia, CA.
b.
QIAquick gel extraction kit, Qiagen Inc., Valencia, CA.
c.
Big-Dye terminator kit, Applied Biosystems, Foster City, CA.
d.
ABI 3130, Applied Biosystems, Foster City, CA.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.