
Abstract
Antibody levels to outer surface proteins C and F (OspC and OspF, respectively) in sera collected from laboratory Beagle dogs at 1, 2, and 4 months after challenge with infected black-legged ticks (Ixodes scapularis) were determined. Each dog was confirmed by culture to harbor Borrelia burgdorferi in the skin (n = 10) or the skin and joints (n = 14). Significant levels of immunoglobulin M (Ig)M or IgG anti-OspC antibodies were detected in single serum samples from only 3 (13%) dogs. Similarly, IgM anti-OspF antibodies were detected in only 1 (4%) serum sample collected from a dog with B. burgdorferi in the skin and joints. In contrast, 4 (29%) dogs with skin and joint infections produced IgG anti-OspF antibodies after 2 months, and the response expanded to include 2 (20%) dogs with skin infection and 4 additional dogs with skin and joint infections (overall sensitivity = 62%) after 4 months. The findings failed to support the utility of OspC-based antibody tests for diagnosing canine Lyme disease, but demonstrated that dogs with B. burgdorferi colonizing joint tissue most often produced significant levels of IgG anti-OspF antibodies. Therefore, additional studies to more thoroughly evaluate the clinical utility of OspF-based antibody tests are warranted.
Keywords
Lyme disease is a tick-associated zoonosis caused primarily by Borrelia burgdorferi in the United States. The bacterium induces a variety of clinical manifestations in humans, including a characteristic skin lesion termed erythema migrans, pericarditis, encephalomyelitis, and arthritis. 14 Dogs also become infected with B. burgdorferi, but the illness differs significantly from human infection. 9 Most notably, humans with Lyme disease reliably develop illness shortly after infection, 14 whereas most infected dogs remain asymptomatic for extended periods of time even after the spirochetes have disseminated to the joints.6,15 Furthermore, the detection of anti-C6 peptide antibodies is currently the only well-established test for confirming canine Lyme disease.5,8 However, conflicting reports7,12 of the ability of the C6 antibody test to discriminate active infection from past exposure have generated considerable confusion, especially when seropositive dogs fail to exhibit clinical abnormalities. 14 Therefore, additional tests based on antibody responses associated exclusively with canine Lyme disease would be helpful, especially if the specificity or intensity of the responses correlated more reliably with the duration and severity of the B. burgdorferi infection.
Researchers have shown that each of 4 dogs challenged with black-legged ticks (Ixodes scapularis) produced immunoglobulin (Ig)G antibodies to outer surface protein (Osp)C within 21 days post-challenge or IgG antibodies to OspF within 49 days post-challenge, 17 which suggested that anti-OspC and anti-OspF antibodies were reliable markers of infection with B. burgdorferi. In addition, the timing of the responses suggested that anti-OspC antibodies were produced primarily during early infection and that anti-OspF antibodies were associated more closely with later stages of the illness. Therefore, detection of the antibody responses could be used to confirm canine Lyme disease and also provide accurate prediction of the duration of the infection. In this study, we examined these possibilities more closely by evaluating the IgM and IgG antibody responses detected by a recombinant (r)OspC or rOspF enzyme-linked immunosorbent assay (ELISA) using sera collected at multiple time points from dogs confirmed by culture to harbor viable Lyme disease spirochetes.
We first recovered rOspC as described previously.
18
Briefly, Escherichia coli JM109
a
expressing OspC from B. burgdorferi sensu stricto isolate 50772
18
was cultured in tryptone–yeast extract (2xTY) broth that contained 100 μg/mL of ampicillin,
b
diluted 10-fold with additional 2xTY broth, and then incubated for an additional hour. Isopropyl-β-
In addition, rOspF was recovered using a modification of a previously described procedure. 17 Briefly, ospF bases 1–684 (GenBank HM756745.1) were synthesized commercially d with SphI (5′) and KpnI (3′) compatible ends, and the fragment was digested with SphI and KpnI, ligated into a pQE-30 Xa expression vector, e and electroporated into E. coli JM109. a The cells were then cultured and pelleted at 8,000 × g for 30 min, washed with PBS as described above, resuspended in ~20 mL of buffer B (100 mM NaH2PO4, 10 mM Tris–Cl, and 8 M urea) adjusted to pH 8, and then lysed by sonication. c The 6× His-tagged rOspF was then recovered by binding to nickel–nitrilotriacetic acid resin e and eluting the protein with buffer B adjusted to pH 6.3. The sequence of the rOspF construct was also confirmed by DNA sequencing. f
We next confirmed the integrity and purity of each recombinant protein by sodium dodecyl sulfate–polyacrylamide gel electrophoresis using standard techniques. We then added 100 μL of carbonate coating buffer (0.02 M Na2CO3, 0.03 M NaHCO3, pH 9.6) that contained either 100 ng of rOspC or rOspF to the individual wells of microtiter plates g and incubated the plates overnight at 4°C. Following incubation, the plates were washed with PBS containing 0.05% Tween 20 (PBST) and blocked with PBST containing 1% bovine serum albumin. One hundred microliters of serum diluted 200-fold with PBST followed by 100 μL of horseradish peroxidase–conjugated anti-dog IgM or IgG h were added to individual wells. The plates were then re-incubated, washed, and o-phenylenediamine substrate b was used to detect bound antibodies. The reactions were stopped by adding 1 N H2SO4, and the absorbances at 490 nm i were determined immediately.
We subsequently established the significant reactivity cutoff values for each ELISA by testing non–Lyme disease (normal) sera (n = 72) from client-owned dogs in Wisconsin (n = 15), New Hampshire (n = 44), and New York (n = 13) with no prior clinical or serologic (tested using a commercial assay j ) evidence of infection with B. burgdorferi or suspicion of tick exposure (e.g., city and/or house dogs). The blood was collected by venipuncture in clot-activator tubes, and the serum was then stored frozen at −80°C until tested. The mean of the resultant optical density (OD) values was then calculated, and OD values ≥2 standard deviations (SDs) above the mean were considered significant. The appropriate positive controls were then identified by testing serial 2-fold dilutions of a dog serum that contained a high level of anti-OspC antibodies induced by vaccination against Lyme disease 4 and a human Lyme disease serum that contained a high level of anti-OspF antibodies. The minimum amount (last dilution) of serum necessary to yield significant (OD ≥ 2 SD above mean of normal dog sera) reactivity was then determined. The appropriate positive control serum was then evaluated on each subsequent ELISA plate, and significant reactivity to the same endpoint dilution was required to ensure consistent plate-to-plate reactivity. We also included a normal canine serum control and tested the serum samples from individual dogs on the same ELISA plate.
Archived serum samples from 24 healthy laboratory purpose-bred Beagles that had received only placebo prior to challenge with B. burgdorferi–infected ticks in previous studies3,4 were then evaluated. The dogs were challenged by attaching 10 female and 10 male B. burgdorferi–infected I. scapularis ticks and allowing the female ticks to feed to repletion. Serum samples were collected immediately prior to the tick challenge, and 1, 2, and 4 months postchallenge. In addition, the sera were stored at −80°C without previous thawing until used for this study. Prior to testing, the sera was grouped into 2 separate groups defined as localized skin infection (n = 10), where spirochetes had only been recovered from skin biopsies, or disseminated joint infection (n = 14), where spirochetes were recovered from both skin biopsies and joint tissues (n = 11). In addition, we included 3 dogs in the latter group that had spirochetes recovered from only skin biopsies, but also had arthropathies (n = 2) or episodic lameness (n = 1) characteristic of disseminated B. burgdorferi infection.4,15 As an additional control for intra- or interassay variability, the sera from the B. burgdorferi–infected dogs was also re-evaluated 2 additional times using freshly prepared reagents and ELISA plates, and the subsequent runs each produced identical findings.
Significant levels of IgM anti-OspC antibodies were not detected in the dogs within the first 2 months of infection, and only 3 (13%) dogs produced the antibodies after 4 months (Table 1). In addition, IgG anti-OspC antibodies were detected in only 3 dogs (13%) after 1 month, and the frequency of detection decreased thereafter. Similarly, significant levels of IgM anti-OspF antibodies were detected in only 1 (4%) dog after 2 months, and the response was not detected thereafter. In contrast, however, 4 (29%) dogs with B. burgdorferi in the skin and joints produced IgG antibodies within 2 months, and the response persisted in the dogs after 4 months. In addition, the antibody response was detected in 4 additional dogs with joint infections (overall sensitivity = 62%) after 4 months, and was also detected in 2 (20%) additional dogs with B. burgdorferi recovered from only the skin.
Detection of anti–outer surface protein (Osp)C or anti-OspF antibodies after infection with Borrelia burgdorferi.*
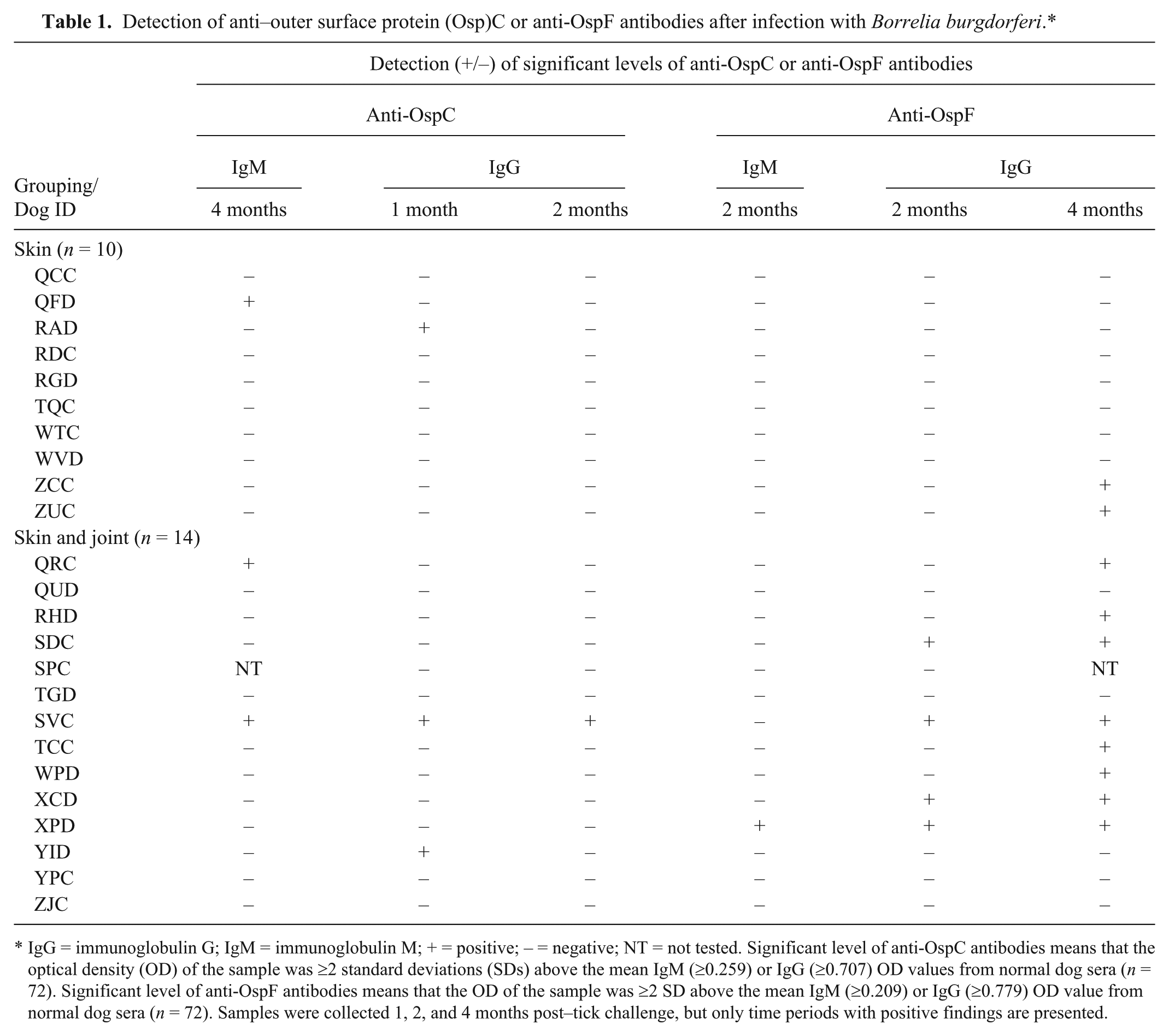
IgG = immunoglobulin G; IgM = immunoglobulin M; + = positive; – = negative; NT = not tested. Significant level of anti-OspC antibodies means that the optical density (OD) of the sample was ≥2 standard deviations (SDs) above the mean IgM (≥0.259) or IgG (≥0.707) OD values from normal dog sera (n = 72). Significant level of anti-OspF antibodies means that the OD of the sample was ≥2 SD above the mean IgM (≥0.209) or IgG (≥0.779) OD value from normal dog sera (n = 72). Samples were collected 1, 2, and 4 months post–tick challenge, but only time periods with positive findings are presented.
The findings therefore failed to provide support for the ability of an OspC-based antibody test to confirm canine Lyme disease, and instead confirmed a previous report 11 that, in contrast to the immunodominant anti-OspC IgM antibody response detected during early human Lyme disease,2,10,18 dogs rarely produce anti-OspC antibodies. In addition, the results contrasted sharply with a previous report 17 that IgG anti-OspC antibodies were produced reliably during the early stages of canine B. burgdorferi infection. Therefore, additional studies to explain the discrepant results remain necessary.
One possible explanation for the decreased detection of IgG anti-OspC antibodies between the current study and the prior report could be differences in the antigen loads of the challenge, as we evaluated the antibody responses after challenging the dogs with 10 ticks and dogs were challenged with up to 50 ticks in the previous study. 17 However, dogs would seem likely to also contract infection after attack by less than 50 I. scapularis ticks, especially in areas where Lyme disease is endemic and owners have been well versed in preventing the ticks from attaching. If this possibility occurs, then our findings provide strong evidence that confirming the infection by detecting anti-OspC antibodies would be difficult.
Another possible explanation is that the heterogeneity of OspC, which is well known to have confounded efforts to use detection of anti-OspC antibodies for serodiagnostic confirmation of human Lyme disease,16,18 may have contributed to the decreased sensitivity of the ELISA by failing to detect antibodies formed against B. burgdorferi isolates that express other ospC genotypes. In support, we used an OspC that reliably binds anti-OspC antibodies produced by dogs after vaccination with a Lyme disease bacterin3,4 and those formed during human infection. 18 However, dogs may also be infected with ospC genotypes that differ significantly from those associated commonly with human infection. 13 As further support, a 2014 report 1 demonstrated significant genotype-specific diversity in the anti-OspC antibody responses after B. burgdorferi infection of laboratory Beagles by tick bite. Moreover, we used an OspC that was derived from an ospC with only 87% sequence identity compared with the ospC used previously. 17 However, if heterogeneity was responsible for the discrepant results, our findings still cast doubt on the widespread utility of a test that depends on antibody binding to an OspC derived from only a single B. burgdorferi isolate. As an additional confounding factor, previous vaccination would further complicate diagnosis by an OspC-based antibody test because a widely used commercially available vaccine k prevents B. burgdorferi infection specifically by inducing high levels of anti-OspC antibodies against a widely conserved epitope.3,4
In contrast, however, the results provided support for the previous observation 17 that dogs with Lyme disease more reliably produced anti-OspF antibodies, and that the response was associated most closely with later stages of the infection. In addition, we demonstrated that IgG antibodies predominated, and that the response was produced most often by dogs harboring spirochetes in the joints. However, we failed to detect the antibodies in 5 (38%) dogs with culture-confirmed joint infections, and the response was also detected in 2 (20%) dogs with spirochetes in only the skin tissue. Therefore, a negative finding did not rule out infection, and the response was not associated exclusively with dissemination of spirochetes to the joints. Moreover, we did not address the specificity of the IgG antibody response by testing sera from dogs with other illnesses.
In summary, at multiple times post-infection, we tested sera from dogs with viable B. burgdorferi in the skin or the skin and joints for anti-OspC and anti-OspF antibodies. The findings failed to provide support for the clinical utility of OspC-based antibody tests. However, the OspF-based IgG ELISA provided some valuable serodiagnostic evidence of canine Lyme disease, especially after the spirochetes had disseminated to the joints, so additional studies to more completely characterize the clinical usefulness of the assay appear warranted.
Footnotes
Authors’ contributions
SM Callister and RL LaFleur contributed to conception and design of the study. DA Jobe, SD Lovrich, and TL Wasmoen contributed to design of the study. SM Callister and RL LaFleur contributed to analysis and interpretation of data. DA Jobe and SD Lovrich contributed to acquisition and analysis of data. TL Wasmoen contributed to interpretation of data. SM Callister drafted the manuscript. All authors critically revised the manuscript, gave final approval, and agree to be accountable for all aspects of the work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
a.
Promega Corp., Madison, WI.
b.
Sigma-Aldrich, St. Louis, MO.
c.
Model W350, Branson Sonic Power, Danbury, CT.
d.
GenScript USA Inc., Piscataway, NJ.
e.
Qiagen Inc., Valencia, CA.
f.
Laragen Inc., Culver City, CA.
g.
Immunolon 2 HB, Thermo Labsystems, Franklin, MA.
h.
Kirkegaard & Perry Laboratories, Gaithersburg, IL.
i.
VersaMax, Molecular Devices Corp., Sunnyvale, CA.
j.
SNAP-4Dx, IDEXX Laboratories Inc., Westbrook, ME.
k.
Merck Animal Health, Elkhorn, NE.
Declaration of conflicting interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Drs. LaFleur and Wasmoen are employees of Merck Animal Health Inc., and the company manufactures canine Nobivac Lyme bacterin. Dr. Callister is a compensated Merck Animal Health Inc. consultant. The other authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Financial support was provided by Gundersen Health System Medical Foundation.