
Abstract
A species-independent indirect enzyme-linked immunosorbent assay (iELISA) based on chimeric protein A/G was established for the detection of anti-Brucella antibodies in Arctic wildlife species and compared to previously established brucellosis serological tests for hooded seals (Cystophora cristata), minke whales (Balaenoptera acutorostrata), sei whales (Balaenoptera borealis), fin whales (Balaenoptera physalus), and polar bears (Ursus maritimus), as well as bacteriology results for reindeer and caribou (Rangifer tarandus sp.). The protein A/G iELISA results were consistent with the other serological tests with Cohen kappa values between 0.47 and 0.92, and the protein A/G iELISA can thus offer a technically simple method for these species yielding results consistent with established brucellosis serological tests. Receiver operator characteristics analysis proved that the reindeer and caribou protein A/G iELISA results were consistent with the bacteriological gold standard with an area under the curve of 0.99, and the protein A/G iELISA was thus validated as a sensitive and specific serological method for the detection of anti-Brucella antibodies in reindeer and caribou. The binding of the antibodies from the respective species to protein A and G were also evaluated in the iELISA. The antibodies from hooded seals and polar bears reacted stronger to protein A than to G. The sei whale, fin whale, reindeer, and caribou antibodies reacted stronger to protein G than to A. The minke whale antibodies reacted to both protein A and G. There was a strong correlation (rs = 0.88–0.98) between the optical density results obtained with the iELISA with protein A/G and protein A or G, showing that protein A/G is as well suited as protein A or G for the detection of anti-Brucella antibodies in these species with the iELISA.
Bacterial species of the genus Brucella are present in many domestic species, 9 and brucellosis is an important infectious disease in Arctic wildlife, affecting both terrestrial 35 and marine wildlife species. 29 A few wildlife populations have sustainable infections with brucellae, while spillover from domestic animals results in nonsustainable infections in many wildlife species. 13 With a typically limited knowledge on the infection status of wildlife, and restricted access to organ samples, serological tests are essential tools for evaluating exposure of populations to brucellae.
Brucella spp. have been serologically indicated in, and isolated from, marine mammals in numerous geographic locations, including the Arctic. 29 A high number of serology- and bacteriology-positive animals 38 have been detected in the Northeast Atlantic stock of hooded seal (Cystophora cristata), which was approximately 82,400 animals in 2005, only 10–15% of the population estimate in 1946, and has been stable at this low level since the 1980s (International Council for the Exploration of the Sea: 2011, Report of the working group on harp and hooded seals, August 15–19, 2011, St. Andrews, UK. Available at: http://brage.bibsys.no/imr/handle/URN:NBN:no-bibsys_brage_26447). However, gross pathology associated with isolation of brucellae in marine mammals has been reported exclusively in cetaceans. 29 In the current study, the endangered sei whale (Balaenoptera borealis) and fin whale (Balaenoptera physalus), 31 along with the North Atlantic minke whale (Balaenoptera acutorostrata), the only cetacean species still commercially hunted in Norway, were examined. 4 Brucella ceti have been isolated from a seropositive North Atlantic minke whale showing no Brucella-associated pathology, 6 and anti-Brucella antibodies (Abs) have been detected in seemingly healthy sei, fin, and minke whales from the North Atlantic Ocean. 37 Also, the polar bear (Ursus maritimus) is considered a marine mammal, 33 and anti-Brucella Abs have been detected in polar bears, 36 but there are no reports of isolation, and the origin of brucellae in the polar bear has been debated. 14 In reindeer and caribou (Rangifer tarandus sp.), a sustainable infection with Brucella suis biovar 4 is prevalent in Canada, Alaska, 35 and Russia, 41 making reindeer central in an epidemiological and zoonotic context.
Many brucellosis serological tests have been used for wildlife species. 13 The slow agglutination of Wright (SAW) is a sensitive test prone to detect immunoglobulin (Ig)M. The specificity is increased by an acid environment, chemical reduction or precipitation of IgM, or pretreating the serum with ethylenediamine tetra-acetic acid (SAW-EDTA). 30 The Rose Bengal test (RBT) and the buffered antigen plate agglutination test are agglutination tests using acidified antigen. Both may give false negatives. 30 The complement fixation test (CFT) generally has a high specificity and is used as a confirmatory test in eradication programs. 30 The fluorescence polarization assay is a sensitive and specific test directly measuring Ab binding to the antigen. 25 The competitive enzyme-linked immunosorbent assay (cELISA) is species unspecific and has a high specificity and a lower sensitivity. 30 The indirect ELISA (iELISA) typically performs with high sensitivity and sometimes restricted specificity, making it suited for screening purposes. 30
Some serological methods rely on species-specific Abs, making them unsuited for screening wildlife. 13 This can be overcome by using the species-unspecific protein A (PrA) or protein G (PrG) as conjugates. 36 Protein A is a 42-kDa polypeptide cell wall constituent of Staphylococcus aureus. Protein G is a 30–35-kDa polypeptide cell wall constituent of β-hemolytic group A Streptococcus spp. Protein A and PrG bind nonspecifically to the Fc-region of Abs through interaction with the heavy chain. 15 A chimeric protein A/G (PrA/G) combines the binding features of PrA and PrG. 11
The aim of the current study was to establish a species-independent PrA/G iELISA for the detection of anti-Brucella Abs in Arctic wildlife species and compare its coherence to previously established brucellosis serological tests for hooded seals, minke whales, sei whales, fin whales, polar bears, and to bacteriology results for reindeer and caribou. The binding of PrA and PrG to Abs from these species in the iELISA is also assessed.
Materials and methods
Hooded seal serum samples (n = 172) from the Northeast Atlantic stock (71–78°N, 2–18°W) were obtained on scientific expeditions during 2004–2010. Blood samples (in evacuated 10-ml tubes, a from vena and arteria brachialis during terminal bleeding of the seals) stood overnight at room temperature and serum was collected (1,000 × g, 10 min) and stored at −20°C. Included in the study were also sera from minke (n = 189), sei (n = 32), and fin (n = 76) whales, previously investigated for anti-Brucella Abs by SAW, SAW-EDTA, RBT, CFT, PrG iELISA, and an anti-complement ELISA, 37 as well as plasma from polar bears (n = 230) previously investigated for anti-Brucella Abs by SAW, SAW-EDTA, RBT, and a PrA iELISA. 36 Included in the study were also sera from reindeer (Rangifer tarandus tarandus) from Norway (n = 304) 10 considered as brucellosis negative based on previous investigations, 3 and given that Norway has been free of brucellosis in terrestrial animals for more than 6 decades (Hofshagen M, Lange H, Hauge K: 2011, Norway 2011–trends and source of zoonoses and zoonotic agents in humans foodstuffs, animals and feedingstuffs, including information on food-borne outbreaks, antimicrobial resistance in zoonotic agents and some pathogenic microbiological agents. Annual report according to Council Directive 2003/99/EC. Available at: http://www.vetinst.no/Publikasjoner/Zoonoserapporten), and also sera from bacteriology- and serology-positive barren-ground caribou (Rangifer tarandus groenlandicus) from Canada (n = 34). 2
The iELISA was performed as previously described,22,32 with minor modifications. Tween-20, b Tween-80, b phosphate-citrate buffer, b o-phenylenediamine dihydrochloride, b PrA, c PrG, d and PrA/G e were purchased commercially. The wells of 96-well polystyrene plates f were coated with Brucella abortus lipopolysaccharide (2 μg/ml; 3.5 hr at 37°C, and minimum 24 hr at 4°C). The plates were incubated in the dark for 12 min at room temperature after adding the substrate. A negative control (fetal calf serum), buffer control (sample dilution buffer), and positive control (serum from a bacteriology positive hooded seal or caribou, 2 or from a seropositive sei whale, 37 or plasma from a seropositive polar bear 36 ) were included on each plate. The mean optical density (OD) g of duplicate wells was expressed as a percentage of the reactivity of the positive control for the respective species: ([OD sample/OD positive control] × 100) = percent positivity (%P). All samples were analyzed with PrA/G. A subsample was analyzed with all three proteins (Table 1). All hooded seal sera were tested with CFT, SAW-EDTA, and RBT. 39 All reindeer from Norway and the caribou sera were tested with RBT. 39
Mean optical density (OD) from a set of positive (positive with all tests) and negative (negative with all tests) samples from each species tested with protein A, protein G, and protein A/G in the indirect enzyme-linked immunosorbent assay.*
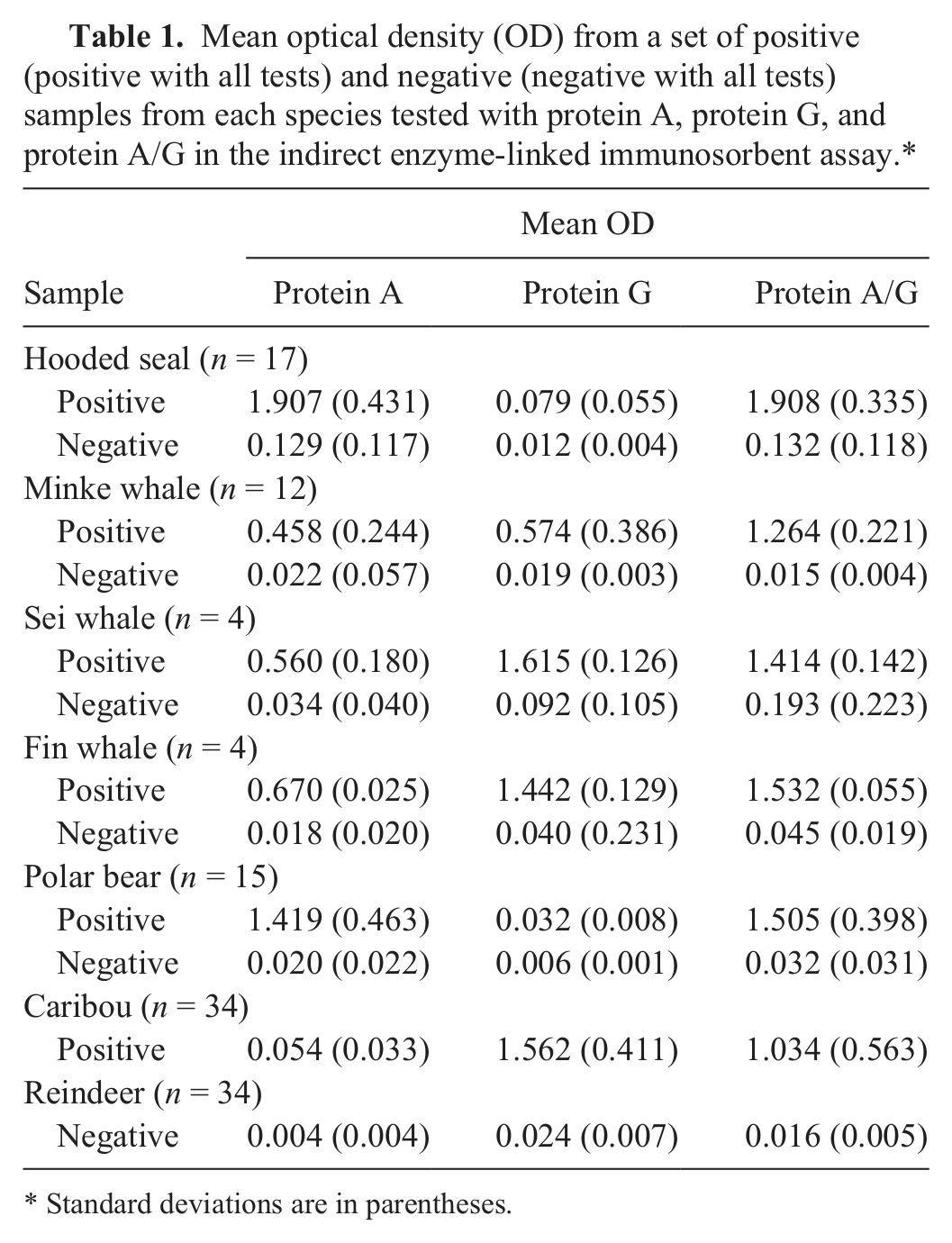
Standard deviations are in parentheses.
Statistical analysis was conducted using StataSE 11 h and R (http://www.R-project.org/). The proportion of samples positive in each test was calculated with 95% confidence intervals (CI) using general linear models with a binomial distribution and logit link. To assess pairwise coherence among the different serological tests, Cohen kappa (κ) was calculated. 7 Receiver operator characteristic (ROC) analysis 12 was performed on the PrA/G iELISA results from the reindeer and caribou using the bacteriology-positive caribou as true positives. 2 The sensitivity (the proportion of positives in the PrA/G iELISA that were from the bacteriology-positive caribou) and specificity (the proportion of negatives in the PrA/G iELISA that were from the negative reindeer from Norway) of the PrA/G iELISA for reindeer and caribou was calculated. The Spearman rank correlation coefficient (rs) 40 quantifies the compliance between the results obtained with the different proteins. The cutoff for the PrA/G iELISA for each species was calculated by the mean value of the specified negative samples plus 2.58 standard deviations (SD). 16
Results
Hooded seal
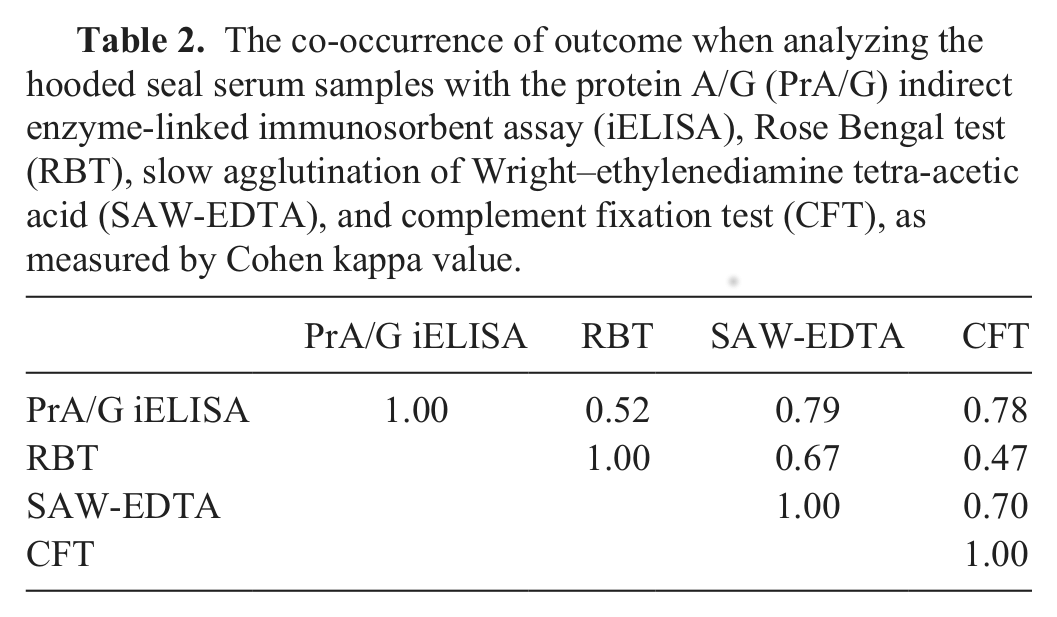
The mean value of the %P for samples negative in CFT, SAW-EDTA, and RBT (125/172) was 18.1 %P with a SD of 21.5 %P, providing a cutoff for the PrA/G iELISA for hooded seals of 73.6 %P. The PrA/G iELISA yielded a seroprevalence of 23.8% (41/172, 95% CI: 17.4–30.3), while the seroprevalence found with CFT was 26.2% (45/172, 95% CI: 19.5–32.8), followed by SAW-EDTA (31/172, 18.0%, 95% CI: 12.2–23.8) and RBT (17/172, 9.9%, 95% CI: 5.4–14.4). There was a good pairwise fit between the tests (Cohen kappa; Table 2). The proportion of samples positive in 1 or 2 of the CFT, SAW-EDTA, and RBT was 17.4% (30/172, 95% CI: 11.7–23.2). The proportion of samples positive in CFT, SAW-EDTA, and RBT simultaneously was 9.9% (17/172, 95% CI: 5.4–14.4). Boxplots show the distribution of the %P in the PrA/G iELISA for the groups “negative in CFT, SAW-EDTA, and RBT,” “positive in 1 or 2 out of CFT, SAW-EDTA, and RBT,” and “positive in CFT, SAW-EDTA, and RBT” (Fig. 1). The hooded seal sera reacted stronger with PrA and PrA/G, compared to PrG (Table 1). The %P obtained with PrA and PrA/G in the iELISA were well correlated (rs = 0.93, P < 0.01, Spearman rank correlation).
The co-occurrence of outcome when analyzing the hooded seal serum samples with the protein A/G (PrA/G) indirect enzyme-linked immunosorbent assay (iELISA), Rose Bengal test (RBT), slow agglutination of Wright–ethylenediamine tetra-acetic acid (SAW-EDTA), and complement fixation test (CFT), as measured by Cohen kappa value.
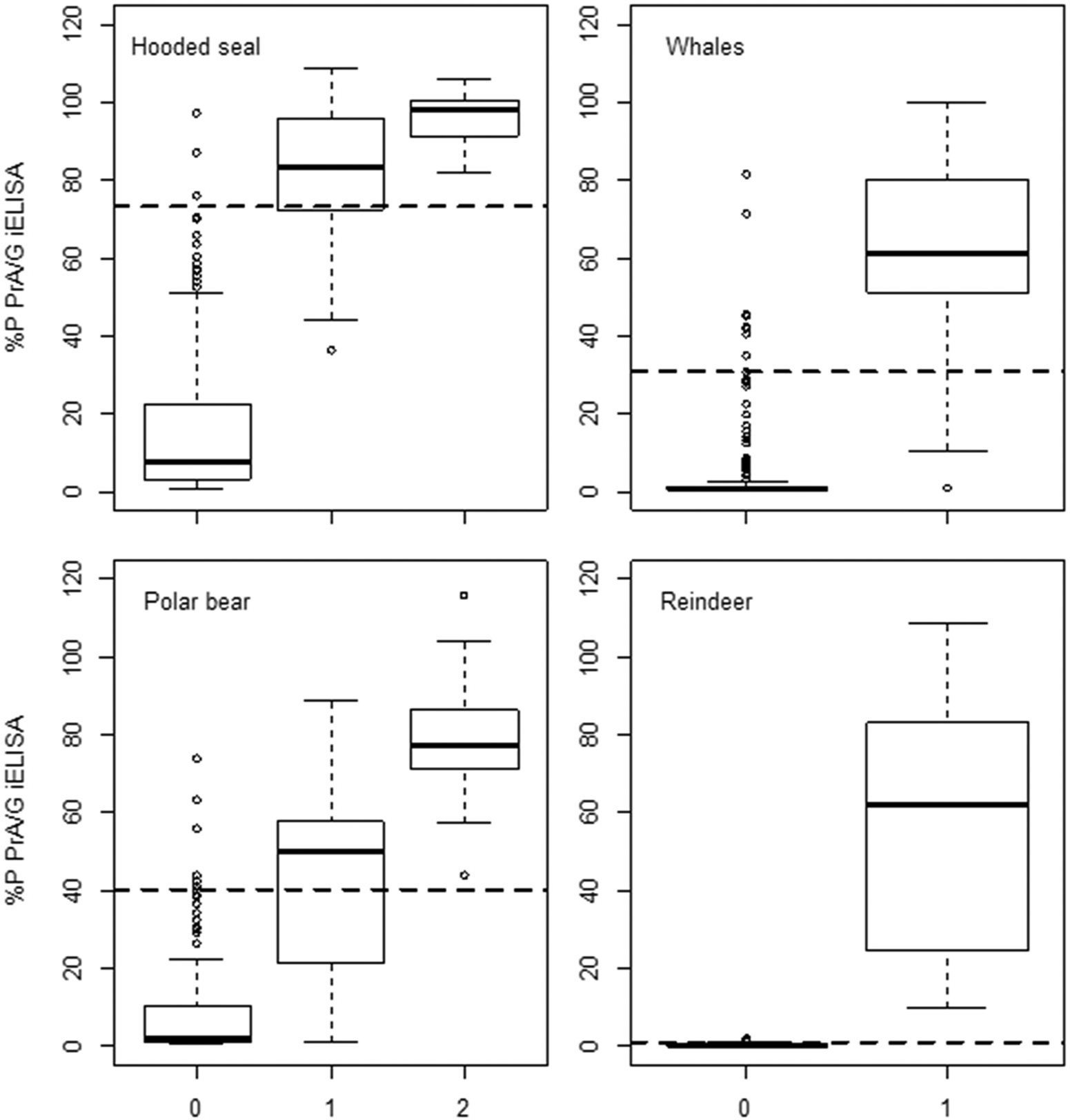
Boxplots showing the median, the 25% and 75% quartiles, whiskers (1.5 times the interquartile range), and the outliers for the protein A/G (PrA/G) indirect enzyme-linked immunosorbent assay (iELISA) percent positivity ([optical density sample/optical density positive control] × 100 = percent positivity [%P]) for each species. Categories for hooded seal: 0 = negative in complement fixation test (CFT), slow agglutination of Wright–ethylenediamine tetra-acetic acid (SAW-EDTA), and Rose Bengal test (RBT); 1 = positive in 1 or 2 out of CFT, SAW-EDTA, and RBT; and 2 = positive in CFT, SAW-EDTA, and RBT. Categories for whales: 0 = negative in the previous serological study 37 ; and 1 = positive in the previous serological study. 37 Categories for polar bear: 0 = negative in SAW, SAW-EDTA, and RBT; 1= positive in 1 or 2 out of SAW, SAW-EDTA, and RBT; and 2 = positive in SAW, SAW-EDTA, and RBT. Categories for reindeer: 0 = bacteriology-negative reindeer from Norway; and 1 = Brucella suis biovar 4 bacteriology-positive caribou. The cutoff for the PrA/G iELISA for each species is indicated with a dotted line.
Whales
Data showing which samples were positive in all tests simultaneously was obtained from a previous study 37 on whales (SAW, SAW-EDTA, RBT, CFT, PrG iELISA, and an anticomplement ELISA; 28/297, 9.4%, 95% CI: 6.1–12.8). The mean value of the %P of whales previously found negative (269/297) was 4.0 %P with a SD of 10.4 %P, providing a cutoff for the PrA/G iELISA for whales of 30.8 %P. The PrA/G iELISA yielded a seroprevalence of 12.2% (23/189, 95% CI: 7.5–16.9) for minke whales, 5.3% (4/76, 95% CI: 0.1–10.4) for fin whales, and 12.5% (4/32, 95% CI: 0.39–24.6) for sei whales. When comparing the PrA/G iELISA results with the previous results, 37 there was a good pairwise fit between the results (Cohen kappa); κ = 0.69 for minke whales (negative in both studies: 164/189; diverging results in the two studies: 11/189; positive in both studies: 14/189), κ = 0.64 for fin whales (negative in both studies: 68/76; diverging results in the 2 studies: 4/76; positive in both studies: 4/76), κ = 1.00 for sei whales (negative in both studies: 28/32; positive in both studies: 4/32), and κ = 0.72 for the whales as a group (negative in both studies: 260/297; diverging results in the two studies: 15/297; positive in both studies: 22/297). Figure 1 illustrates the distribution of the %P in the PrA/G iELISA for the groups “negative in the previous study” and “positive in the previous study” by boxplots. The sei and fin whale Abs reacted stronger with PrG than with PrA, while the minke whale Abs reacted equally with PrA and PrG. All whale species reacted with PrA/G (Table 1). The %P obtained with PrG and PrA/G in the iELISA were well correlated (rs = 0.88, P < 0.01, Spearman rank correlation).
Polar bear
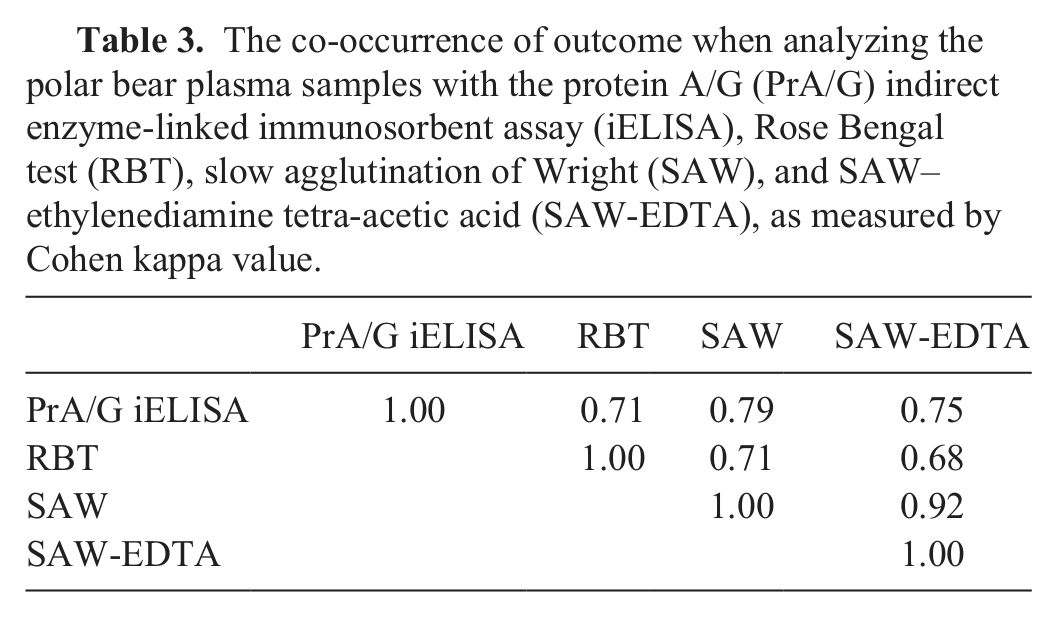
The mean value of the %P of samples negative in SAW, SAW-EDTA, and RBT 36 (198/230) was 8.2 %P with a SD of 12.5 %P, providing a cutoff for the PrA/G iELISA for polar bears of 40.5 %P. The PrA/G iELISA yielded a seroprevalence of 13.5% (31/230, 95% CI: 9.0–17.9), while the seroprevalence found with SAW-EDTA was 13.0% (30/230, 95% CI: 8.7–17.4), followed by SAW (28/230, 12.2%, 95% CI: 7.9–16.4) and RBT (18/230, 7.8%, 95% CI: 4.3–11.3). There was a good pairwise fit between the tests (Cohen kappa; Table 3). The proportion of samples positive in 1 or 2 of the tests SAW, SAW-EDTA, and RBT was 6.5% (15/230, 95% CI: 3.3–9.7). The prevalence found when considering samples only as positive if they were positive in SAW, SAW-EDTA, and RBT simultaneously was 7.4% (17/230, 95% CI: 4.0–10.8). The distribution of the %P in the PrA/G iELISA for the groups “negative in SAW, SAW-EDTA, and RBT,” “positive in 1 or 2 out of SAW, SAW-EDTA, and RBT,” and “positive in SAW, SAW-EDTA, and RBT” are illustrated in Figure 1. The polar bear Abs reacted less with PrG as compared to PrA and PrA/G (Table 1). The %P obtained with PrA and PrA/G in the iELISA were well correlated (rs = 0.98, P < 0.01, Spearman rank correlation).
The co-occurrence of outcome when analyzing the polar bear plasma samples with the protein A/G (PrA/G) indirect enzyme-linked immunosorbent assay (iELISA), Rose Bengal test (RBT), slow agglutination of Wright (SAW), and SAW–ethylenediamine tetra-acetic acid (SAW-EDTA), as measured by Cohen kappa value.
Reindeer
The mean value for the samples from the negative reindeer from Norway (n = 304) was 0.36 %P with a SD of 0.30 %P, providing a cutoff for the PrA/G iELISA for reindeer and caribou of 1.13 %P. All caribou samples (n = 34)2 were positive in the PrA/G iELISA and 2 Norwegian samples were weak positives (<2.0 %P), but were negative when tested with RBT, CFT, and SAW-EDTA. 39 All reindeer from Norway were negative, and all caribou were positive in the RBT. 39 Boxplots illustrate the distribution of the %P in the PrA/G iELISA for the groups “brucellosis-negative reindeer from Norway” and “B. suis biovar 4 bacteriology-positive caribou” (Fig. 1). ROC analysis of the PrA/G iELISA results, using as true positives the bacteriology-positive caribou, 2 showed an area under the curve (AUC) of 0.99 (95% CI: 0.99–1.00), a sensitivity of 100.0%, and a specificity of 99.3%, yielding 99.4% of the animals being correctly classified. The reindeer and caribou Abs reacted less with PrA, as compared to PrG and PrA/G (Table 1). The %P obtained with PrG and PrA/G in the iELISA were well correlated (rs = 0.96, P < 0.01, Spearman rank correlation).
Discussion
The PrA/G iELISA offers a serological method for hooded seals, minke whales, sei whales, fin whales, and polar bears, yielding results consistent with established brucellosis serological tests. For reindeer and caribou, the PrA/G iELISA was validated as a sensitive and specific serological method in coherence with a bacteriological gold standard.
When conducting brucellosis serology in wildlife, an important issue is to set, and confirm, a cutoff value. The cutoff value for the reindeer and caribou in the current study was established using the mean value of the %P of reindeer from Norway, utilizing samples that were very likely to be true negatives. ROC analysis of the reindeer and caribou PrA/G iELISA results, using as true positives the bacteriology-positive caribou 2 showed an AUC of 0.99, verifying the cutoff. In the absence of a gold standard, the cutoff for the hooded seal, polar bear, and whales was based on agreement with other serological tests. The present study found that the PrA/G iELISA results for these species, using the suggested cutoffs, were consistent with the results of the other serological tests, as shown by Cohen kappa values ranging from 0.47 to 0.92.
Brucellosis serological tests utilizing species-specific Abs designed for the detection of Abs in a given wildlife species have been developed. 17 For whales, it has been shown that Abs against IgG of toothed whales had a reduced ability to recognize IgG of baleen whales, 28 demonstrating the rationale of producing species-particular Abs against each host species of interest. This often has the drawback of necessitating the production of a finite amount of antispecies Abs in experimental animals that has to be renewed, which is not in line with the principles of reduction, refinement, and replacement of animal testing. 24
Protein A or PrG are good alternatives to species-specific Abs. The Fc-region, and thus the affinity of PrA and PrG to the Fc-region, differs between Abs from different species. 15 Carnivore Abs are shown to react mostly with PrA, 22 and PrA has been used to detect anti-Brucella Abs in polar bear 36 and seals. 38 Protein G has also been used to detect anti-Brucella Abs in seals, 37 even though seal Abs have been shown to react less to PrG, as compared to PrA. 34 Abs from bowhead whale (Balaena mysticetus) react less with PrA, as compared to the Atlantic bottlenosed dolphin (Tursiops truncatus), 21 and both PrA 18 and PrG 37 have been used to detect anti-Brucella Abs in various dolphin and whale species. The Abs of most ungulates react predominantly with PrG. 34 The current study confirms the species variation in the ability of the Abs to bind PrA and PrG; the Abs from hooded seal and polar bear reacted predominantly with PrA, whereas the reindeer, sei whale, and fin whale Abs reacted strongest with PrG, and the minke whale Abs reacted with both PrA and PrG.
Protein A/G combines the binding features of PrA and PrG, 11 reacting with Abs from a number of species, 26 and has been used to detect anti-Brucella Abs in Hawaiian monk seal (Monachus schauinslandi) 27 and Australian fur seal (Arctocephalus pusillus doriferus). 23 In a multispecies assay, PrA/G reduces the risk of using the wrong conjugate, which can be particularly important when working with wild animals where the affinity of the Abs is not known and the serum might be sparse and valuable. Some species might also show surprising Ab binding capability, as demonstrated for the minke whale in the present study, while certain species might also have considerable intraspecies variability in their Ab binding. 19 Antibodies from all species included in the current study bound readily to PrA/G, and there was a strong correlation (rs = 0.88–0.98) between the OD results obtained with the iELISA with PrA/G and PrA or PrG.
Two (of the 304) samples from the reindeer from Norway were classified as weak positives (<2.0 %P) in the PrA/G iELISA, but were negative with RBT, CFT, and SAW-EDTA. 39 The samples were regarded as false positives, in agreement with the finding of no seropositive individuals during assaying for brucellosis of more than 5,500 reindeer from Finnmark County, Norway, in 1990–1994, 3 and given that Norway has been free of brucellosis in terrestrial animals for more than 6 decades (Hofshagen M, Lange H, Hauge K: 2011, Norway 2011).
Serological cross-reactions and false positives are major problems when detecting anti-Brucella Abs by serological methods. 8 In cattle, most of the problems arise from an immune response of the animal to another microorganism that shares epitopes with brucellae O-polysaccharides, 8 with Yersinia enterocolitica O:9 being the most important. 5 In a large study of marine mammals from the North Atlantic Ocean, all animals seropositive for brucellae were seronegative for Y. enterocolitica 0:9. 37 In polar bears, titers against Y. enterocolitica 0:9 have been detected in a few samples regardless of their anti-Brucella titers. It was thus concluded that the positivity in the brucellosis serological tests was not due to infection with Y. enterocolitica 0:9. 36 Investigation of reindeer in Norway yielded solely animals negative for Y. enterocolitica O:9. 20 Some other known possible cross-reacting agents are Francisella tularensis, Salmonella group N (O:30), Escherichia coli O:157 and O:116, Stenotrophomonas maltophilia, and Vibrio cholera, 1 but in wildlife research, it is difficult to assess whether agents are cross-reacting when little is known regarding the agents present and their ability to cross-react. When interpreting results, diagnosticians should be aware of their possible presence, and always interpret serological results with caution. Thus, the gold standard in brucellosis diagnostics still remains bacterial isolation.
The aim of the current study was to establish a species-independent PrA/G iELISA for the detection of anti-Brucella Abs in Arctic wildlife species and to compare its coherence to previously established brucellosis serological tests for hooded seals, minke whales, sei whales, fin whales, and polar bears and to bacteriology results for reindeer and caribou. However, to use the true gold standard, the PrA/G iELISA results should be compared to isolation of the causative organism under controlled conditions for all species included in the study. This was not possible given that organ samples were not readily available from all included species; however, Cohen kappa values confirmed that the PrA/G iELISA results were consistent with the findings obtained when using previously established brucellosis serological tests for hooded seal, minke whale, fin whale, sei whale, and polar bear. The PrA/G iELISA might hence offer a technically simple method for these species yielding results consistent with established brucellosis serological tests, and might also be an improvement compared to some of the serological tests that are sensitive to the quality of the sample. ROC analysis confirmed that the results obtained with the PrA/G iELISA for reindeer and caribou were consistent with the bacteriological gold standard; thus, the PrA/G iELISA was validated as a highly sensitive and specific serological method for reindeer and caribou.
Footnotes
Acknowledgements
The authors would like to thank E. M. Breines and E. Hareide at the Norwegian School of Veterinary Science, Section for Arctic Veterinary Medicine, Tromsø, Norway for excellent laboratory work, and C. de Smedt and D. Fretin at the Veterinary and Agrochemical Research Centre, Brussels, Belgium, for helping with design and validation of the iELISA. The authors also acknowledge T. Haug, K. T. Nilssen, A. K. Frie, N-E Skavberg, andM. T. Poltermann at the Institute of Marine Research, Tromsø, Norway, for making the sampling of hooded seals possible; Ø. Wiig, A. Derocher, J. Aars, and M. Andersen for samples from polar bears; and M. Kjeld, A. Alfredson, and A. Arnason, for samples from fin and sei whales from Iceland. The authors greatly appreciate the help from A. Evans in correcting the language.
a.
Venoject tubes for biochemistry (serum; no additives, 10 ml), Terumo Medical Corp., Somerset, NJ.
b.
Sigma Aldrich, St. Louis, MO.
c.
Protein A-Peroxidase from Staphylococcus aureus, horseradish (1 mg/ml, 1:2,000 dilution, HRP:protein 1:2), Sigma Aldrich, St. Louis, MO.
d.
Purified Recomb Protein G, peroxidase conjugated (1 mg/ml, 1:2,000 dilution, HRP:protein 2:1), Thermo Fisher Scientific Inc., Waltham, MA.
e.
Purified Recomb Protein A/G, peroxidase conjugated (1 mg/ml, 1:16,000 dilution, HRP:protein 1:1), Thermo Fisher Scientific Inc., Waltham, MA.
f.
Nunc PolySorp 475094, Thermo Fisher Scientific Inc., Waltham, MA.
g.
Epoch Microplate Spectrophotometer, BioTek Instruments Inc., Winooski, VT.
h.
Stata: Release 11, StataCorp LP, College Station, TX.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work was supported by Astri and Birger Torsteds grant, Norway.