
Abstract
The pathology of Clostridium perfringens type C infection has been described with detail only in foals and piglets. The current report describes the diagnostic workup and detailed pathology of 3 cases of C. perfringens type C infection in calves. A 2-day-old Jersey calf and fresh and fixed tissues from a 4-week-old Angus calf and from a 1-week-old Jersey calf were received at the California Animal Health and Food Safety Laboratory System with a history of digestive disease and death. The gross changes in the gastrointestinal tract of 1 calf consisted of multifocal subserosal hemorrhages of the rumen, diffuse congestion and multifocal hemorrhages of the small intestinal mucosa, and dilated cecum with bloody liquid contents. In a second calf, a large segment of small intestine was hemorrhagic. The small intestine of the third calf was dilated and filled with abundant yellow fluid content. Microscopically, all 3 calves had diffuse coagulation necrosis of the intestinal mucosa. Clostridium perfringens type A was isolated from the intestinal content of 2 calves. In addition, enzyme-linked immunosorbent assay for Bovine rotavirus was positive on colonic content of 1 calf. Small numbers of cryptosporidia were seen in smears of colonic content of 2 calves, and Salmonella sp. group E was detected in the small intestinal content of another calf. Clostridium perfringens beta toxin was detected in the intestinal content of the 3 animals. A diagnosis of C. perfringens type C infection was confirmed based on pathological findings and detection of beta toxin in the intestinal content of the 3 animals.
Keywords
Clostridium perfringens is classified into 5 types (A–E) based on the production of 1 or more of 4 major toxins, namely alpha (CPA), beta (CPB), epsilon (ETX), and iota (ITX). 16 Two other major toxins, enterotoxin (CPE) and beta2 (CPB2), can also be produced by strains of all types of C. perfringens, but are usually not used in the classification of this microorganism. 4 In addition, most C. perfringens isolates can produce several so-called minor toxins. Clostridium perfringens type C produces at least CPA and CPB. 4 CPA is a lecithinase (phospholipase C), which is considered to be the main virulence factor in C. perfringens type A–associated myonecrosis in human beings and animals. 11 However, the contribution of CPA to the virulence of type C isolates is negligible. 12 CPB, on the other hand, is a necrotizing toxin that forms membrane pores in susceptible cells and is considered to be responsible for the intestinal necrosis and systemic alterations (enterotoxemia) seen in type C infections of several animal species. 12
Because CPB is highly susceptible to the action of trypsin, neonatal animals are particularly susceptible to C. perfringens type C infections, due to the low level of intestinal trypsin activity during the first days of life, a consequence of the trypsin inhibitor effect of colostrum. 6 Type C disease also occurs occasionally in adult animals and human beings ingesting significant amounts of food with trypsin inhibitors, such as sweet potato and soy bean, and also in patients with pancreatic disease.4,5,8
Infection by C. perfringens type C occurs worldwide and affects several animal species. 18 The pathology of the disease has been described with some detail in foals and piglets.4,15 Although several textbooks describe the occurrence of C. perfringens type C infection in calves,1,7,13 the present authors could not locate any published report characterizing the pathology of the condition in calves with confirmed detection of CPB in intestinal contents. The present report describes the diagnostic workup and intestinal pathology of 3 cases of C. perfringens type C infection in calves, confirmed by detection of CPB in intestinal content.
A whole carcass of a 2-day-old Jersey calf (calf 1) and fresh and formalin-fixed tissues from a 1-month-old Angus calf (calf 2) and from a 7-day-old Jersey calf (calf 3) from 3 different farms were received at the California Animal Health and Food Safety Laboratory (CAHFS) for necropsy and diagnostic workup. Calf 1 was 1 of 3 calves with a history of scours. Calf 2 was found depressed, reluctant to stand, and anorectic before it died within 24 hr of being found sick. Calf 3 was from a farm that had lost five 3–6-day-old calves 2 days previously, after developing acute diarrhea.
The following ancillary tests were performed following CAHFS’s standard operating procedures, unless otherwise specified. Samples of small intestinal contents, liver, lung, and/or spleen of the 3 animals were aseptically collected and inoculated onto 5% sheep blood Columbia agar plates a and incubated in 5–10% CO2 at 37°C for 48 hr. In addition, small intestinal contents of the 3 animals were inoculated onto Brucella, phenylethanol, and egg yolk agar plates a and incubated anaerobically at 37°C for 48 hr. Clostridium perfringens isolates were typed by a multiplex polymerase chain reaction (PCR) technique to amplify segments specific for the genes encoding CPA, CPB, ETX, ITX, CPE, and CPB2, as previously described. 2 A real-time PCR to detect a fragment of the Salmonella-specific invA gene was performed on intestinal content of calf 3 as previously described. 3 Salmonella culture was performed using tetrathionate or selenite enrichment broth and selective plate media on bile and colon pool and intestinal content of calves 1 and 3, respectively. Small intestinal content from calves 1 and 3 was also tested for Escherichia coli K99 antigen by a latex agglutination technique, according to the instructions of the manufacturer. b Samples of small intestinal content of the 3 calves were tested for C. perfringens and the CPA, CPB, and ETX toxins via commercial capture enzyme-linked immunosorbent assay (ELISA) kit, c following the manufacturer’s instructions.
Samples of small and large intestine, lung, liver, kidney, thymus, spleen, heart, abomasum, rumen, cecum, and/or adrenal gland were collected from calf 1 and fixed by immersion in 10% buffered formalin (pH 7.2) for 24–72 hr. Samples of the same tissues from calves 2 and 3 were received in formalin. All tissues were processed by standard histological techniques for the production of 4-μm-thick hematoxylin and eosin–stained sections. Selected sections were also stained with phosphotungstic acid–hematoxylin and Gram stains. Sections of ileum and/or colon from the 3 calves were processed by an indirect immunoperoxidase technique for C. perfringens, using a commercial kit d according to the manufacturer’s instructions, and a rabbit polyclonal anti– C. perfringens antibody e as a primary antibody.
Liver and kidney from calves 2 and 3 were also analyzed for heavy metals (lead, manganese, iron, mercury, arsenic, molybdenum, zinc, copper, and cadmium) by inductively coupled argon plasma emission spectrometry. Selenium concentration in the liver of these 2 animals was determined by inductively coupled plasma spectrometry using hydride generation.
Frozen sections of lung and kidney were processed by fluorescent antibody tests (FATs) for Bovine viral diarrhea virus (BVDV; calves 2 and 3) and Bovine herpesvirus 1 (BHV-1; calf 2). Frozen sections of small intestine and/or colon were processed for FATs for Betacoronavirus 1 (BCV; calves 1 and 3) and Bovine rotavirus (BRV; calf 3). All FATs were performed using commercial conjugates. b Colonic contents from the 3 calves were processed for BRV ELISA according to the instructions of the manufacturer. f Smears of intestinal contents from calves 1 and 3 were stained by modified Kinyoun acid-fast stain to detect Cryptosporidium spp.
Externally, calf 1 had evidence of dehydration, including sunken eyes and loss of skin elasticity. Internally, there were many multifocal subserosal hemorrhages of the rumen. The jejunum and ileum were dilated with dark red fluid in the lumen and showed diffuse congestion and hemorrhage of the mucosa (Fig. 1). The spiral colon and cecum contained a moderate amount of bloody liquid content. No information was available on the external appearance of the carcasses of calves 2 and 3. The fresh segment of small intestine received from calf 2 was transmurally dark red to black and contained dark red to black fluid content. The fresh segment of small intestine of calf 3 received was moderately dilated and filled with yellow fluid content.
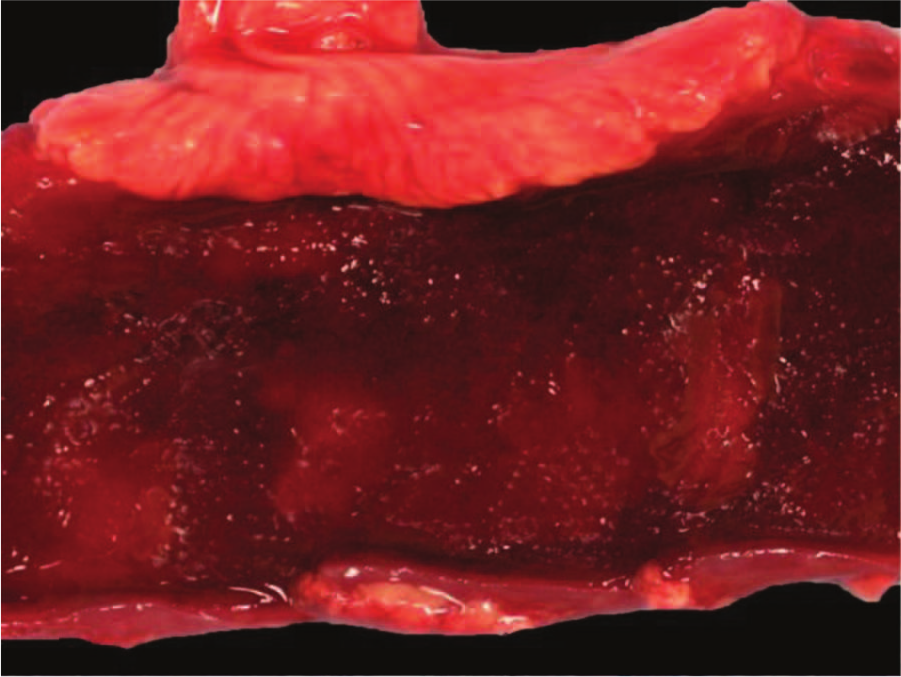
Jejunum, calf 1. Gross appearance of the jejunum mucosa showing diffuse congestion and hemorrhage.
Microscopically, the most striking changes were observed in the small intestine. All 3 calves had changes in the jejunum and ileum consisting of diffuse coagulation necrosis affecting the epithelium and superficial lamina propria. There was also severe villus blunting, and the superficial lamina propria showed diffuse hemorrhage and severe infiltration of viable and dead neutrophils and fewer plasma cells, lymphocytes, and macrophages (Fig. 2). Multifocally, large numbers of neutrophils were seen leaving the lamina propria at the tip of the villi through areas of epithelial necrosis (volcano lesions). Occasionally, there was segmental neutrophilic necrotizing vasculitis and fibrin thrombi of mucosal and submucosal blood vessels (Fig. 3) in calves 2 and 3. Diffuse hemorrhage was present in the deep lamina propria, muscularis mucosa, and submucosa, which, in calf 2, extended to the muscularis and serosa. Overlying the jejunal mucosa of calf 1 was a fibrinonecrotic pseudomembrane composed of fibrin, necrotic cells, cell debris, neutrophils, and myriad basophilic rods. The jejunum of calves 2 and 3 had large numbers of Gram-positive rods lining the remnants of the villi and also within the crypts of Lieberkühn (Fig. 4), which multifocally had degenerate neutrophilic aggregates mixed with necrotic cell debris within the lumen. Calf 3 had a moderate number of Cryptosporidium spp. adhered to the remaining or sloughed epithelial cells in the jejunum.
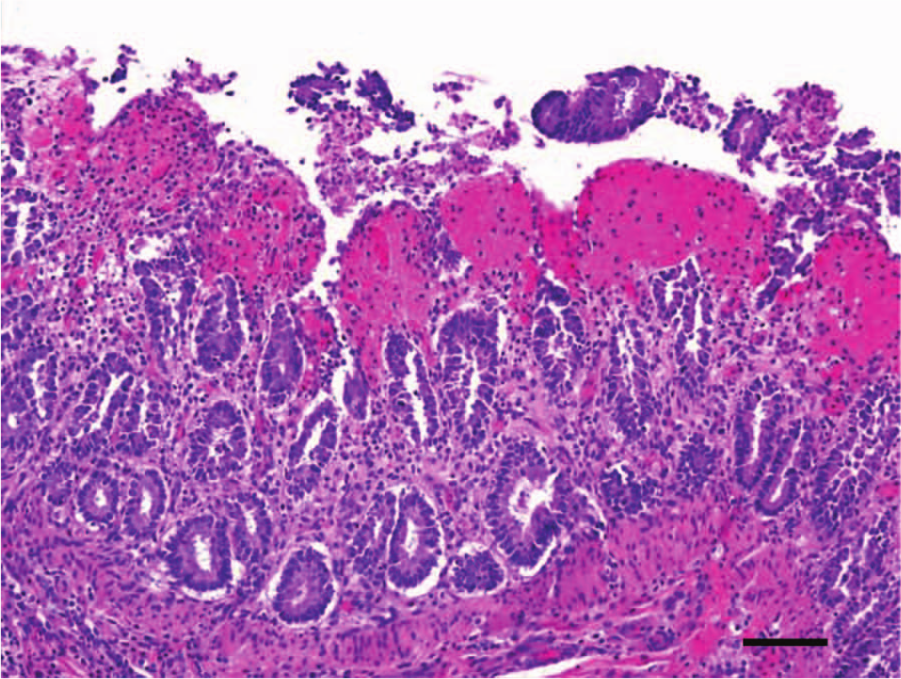
Jejunum, calf 3. Diffuse coagulative necrosis of the epithelium and lamina propria, with pseudomembrane formation. The lamina propria presents severe diffuse hemorrhage and infiltration with neutrophils, plasma cells, lymphocytes, and macrophages. Hematoxylin and eosin. Bar = 50 μm.
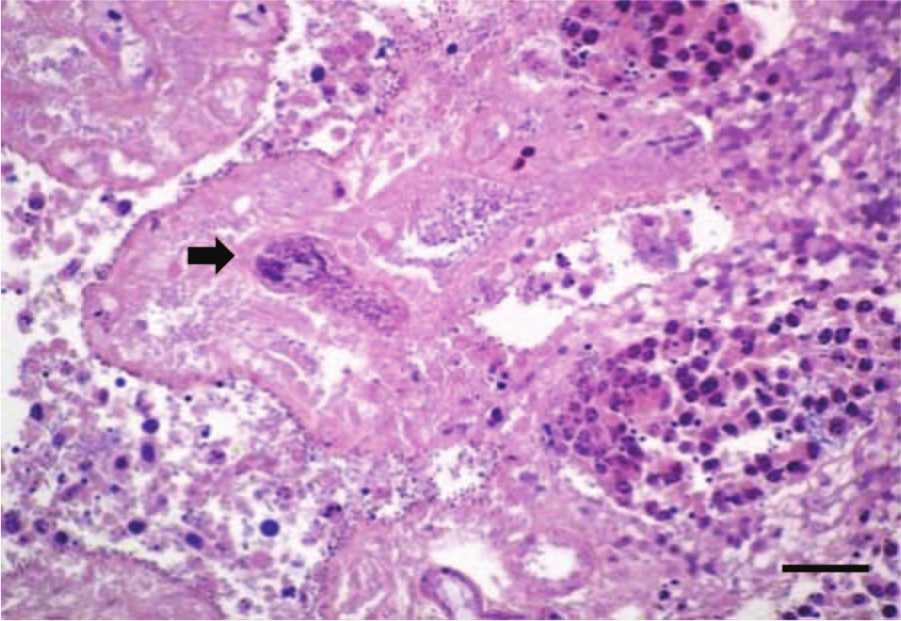
Jejunum, calf 2. Fibrin thrombi within small caliber artery of the lamina propria (arrow). Phosphotungstic acid–hematoxylin stain. Bar = 35 μm.
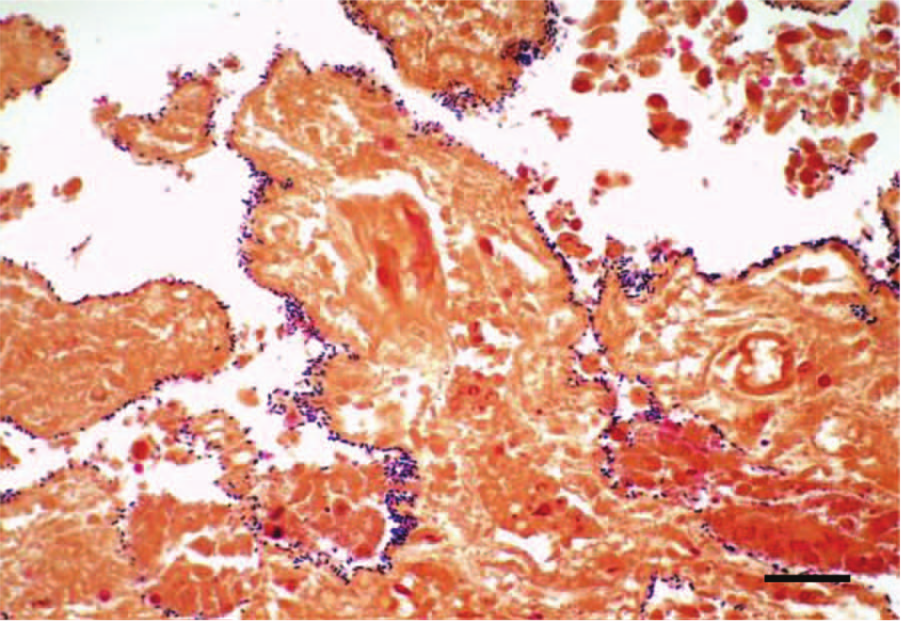
Jejunum, calf 2. Gram stain showing Gram-positive bacilli lining the denudated intestinal villi. Bar = 35 μm.
Microscopic changes outside the small intestine were observed in all calves. The cecum and spiral colon of calf 1 had multifocal extensive mucosal and less frequently submucosal hemorrhages with occasional extension to the serosa. The reticulum and rumen of this calf had multifocal submucosal and subserosal hemorrhages, thrombosis, and occasional necrotizing vasculitis. The abomasum had diffuse mucosal hemorrhage and rare foci of necrosis of the superficial epithelium, with fibrin and neutrophilic infiltration. The mesenteric lymph nodes of calf 2 showed many neutrophils and fewer macrophages expanding the medullary and subcapsular sinuses. The rumen and omasum mucosa of calf 3 had multifocal intraepithelial pustules, and the lamina propria of these organs was edematous and contained mild diffuse infiltration of neutrophils and macrophages. No other significant histological lesions were observed in any of the other tissues from the 3 calves examined.
Clostridium perfringens (typed as type A CPE and CPB2 negative by PCR) was isolated from the small intestinal content of calves 1 and 2. Bacterial aerobic and anaerobic cultures of small intestinal content from all calves were negative or yielded mixed flora. Salmonella sp. group E (not typed) was detected by PCR and culture in the small intestinal content of calf 3. Latex agglutination for E. coli K99 was negative in calves 1 and 3.
Clostridium perfringens CPB was detected in the small intestinal content of the 3 calves by ELISA, while CPA was detected in small intestinal content of calves 2 and 3. Immunohistochemical staining for C. perfringens in the small intestine of all 3 calves was positive, staining most of the rods seen on hematoxylin and eosin–stained sections.
The FATs for BCV, BHV-1, and BVDV were negative in all the samples tested, while ELISA for BRV was positive on colonic content of calf 3. Small numbers of cryptosporidia were seen in smears of colonic content of calves 1 and 3. The heavy metal and selenium concentrations in the liver samples of calves 2 and 3 were within the normal ranges.
In the present 3 cases, a presumptive diagnosis of C. perfringens type C enterotoxemia was made based on gross and microscopic lesions of necrotizing enteritis, and it was confirmed by detection of CPB in the intestinal content of all 3 calves. Detection of CPB in intestinal contents is considered the gold standard for a diagnosis of C. perfringens type C enterotoxemia in horses, 4 pigs, 15 sheep, and goats. 17 While no specific studies have been performed on the diagnostic significance of detection of CPB in the intestine of calves, based on the information available for other animal species,4,15,17 it is likely that the presence of CPB in the intestine of calves is also diagnostic for type C enterotoxemia. This speculation is also supported by the fact that this toxin is not usually detected in the intestinal content from healthy calves (F. Uzal, unpublished observation). Clostridium perfringens type C must produce, at minimum, CPA and CPB. 4 CPA, however, was only detected in the intestinal content of 2 calves. This is not unusual, and it was probably due to the variation in the amount of CPA produced by different strains of C. perfringens. It is likely that the amount of CPA produced in the intestine of calf 1 was below the limit of detection of the ELISA. It is not possible to rule out, however, that CPA was not produced at all by the strain(s) of C. perfringens present in the intestine of calf 1. Nevertheless, detection of CPA in the intestine of these animals was not considered to be of diagnostic significance, as this toxin is detected in the intestine of most healthy animals of several different species, including cattle. 17 Furthermore, the contribution of this toxin to intestinal disease is considered negligible. 12
Failure to isolate C. perfringens type C in the calves in the current study does not preclude a diagnosis of type C enterotoxemia because detection of CPB was positive in all cases. Clostridium perfringens type C infection can have a patchy, localized intestinal distribution4,14 with some segments of intestine being devoid of this microorganism. It is possible that, in these cases, the samples collected were from areas of the intestine where C. perfringens type C was either not present or present in numbers below the sensitivity of the culture method.
Neonates acquire type C organisms from an environment contaminated by sick animals or, less frequently, by asymptomatic carriers. 9 CPB is exquisitely sensitive to the action of trypsin and other proteases. 4 Ingestion of C. perfringens type C into a protease-deficient intestinal tract, as in the case of neonatal animals, allows for subsistence of CPB resulting in intestinal necrosis, invasion of deeper intestinal layers, and absorption into the systemic circulation. 4 Death may result from the direct effects of the severe intestinal necrosis and diarrhea, from secondary toxemia, or from a combination of both. 6 Clostridium perfringens type A was isolated from the intestinal content of calves 1 and 2. Isolation of type A that produces only CPA among the major toxin in calves is considered of no diagnostic significance as this type is present in the intestinal content of most healthy calves as well. 15
Calves 1 and 3 had lesions in the reticulum, rumen, and abomasa. The circulatory changes observed in the prestomachs of both calves are nonspecific and probably resulted from toxemia. The pustular lesions observed in the mucosa of the prestomachs of calf 3 are commonly seen in young calves associated with the accumulation of milk in the rumen. Salmonella sp. group E, Cryptosporidium spp., and BRV were detected in the intestine of calf 3. In this case, the presence of Cryptosporidium spp. and BRV was not considered to be of diagnostic significance, as neither of these 2 agents produces necrotizing lesions similar to those described in the 3 calves. It cannot be ruled out, however, that these 2 pathogens detected in calf 3 could have had a synergistic role with C. perfringens type C in the development of intestinal lesions in this calf. Salmonella spp., however, can produce similar lesions to those described in the 3 calves, and it is possible that this organism was at least partly responsible for the lesions seen in calf 3.
Clostridium perfringens type C enterotoxemia has been reported to affect several animal species. However, detailed descriptions of the pathology of this disease, to the authors’ knowledge, have only been published for foals and piglets.4,15 An outbreak of enterotoxemia suspected to be produced by C. perfringens type C in neonatal Canadian calves with hemorrhagic and/or necrotic enteritis has been reported. 10 In the Canadian calves, CPB was detected in intestinal content by mouse bioassay; however, a detailed description of the intestinal pathology was not provided for those calves. 10
Footnotes
a.
Hardy Diagnostics, Santa Maria, CA.
b.
VMRD Inc., Pullman, WA.
c.
ELISA kit for enterotoxaemia, Bio-X Diagnostics SPRL, Jemelle, Belgium.
d.
EnVision, Dako North America Inc., Carpinteria, CA.
e.
GenWay Biotech Inc., San Diego, CA.
f.
Sure-Vue Rota test kit, Fisher Healthcare, Houston, TX.
Declaration of conflicting interests
The author(s) declare that they have no conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding for the current report was provided by the California Animal Health and Food Safety Laboratory, School of Veterinary Medicine, University of California, Davis.